
中华大蟾蜍胚后原始大脑皮层神经元自发电活动的发育变化
2014-05-16 07:27梁传成王丽文
沈阳师范大学学报(自然科学版) 2014年2期
彭 霞,郑 妍,王 云,王 畅,梁传成,王丽文
(1.沈阳师范大学 实验教学中心,沈阳 110034;2.沈阳师范大学 化学与生命科学学院,沈阳 110034;3.辽宁中医药大学 附属第二医院,沈阳 110034)
中华蟾蜍(Bufogargarizans)隶属于两栖纲,无尾目,蟾蜍科,蟾蜍属,在我国栖居广泛,数量多,最为常见[1],是生态环境的重要指示动物。目前对蟾蜍及近缘种端脑发育的研究多在形态学和组织学上[2-5],有关其发育电活动的研究尚未见报道。笔者应用微电极电生理学技术,对中华蟾蜍胚后6个典型时期端脑原始大脑皮层进行了电生理研究,探讨中华蟾蜍胚后各阶段端脑原始大脑皮层的发育特性,以期充实无尾两栖动物神经系统资料,为进一步的研究工作奠定基础。
1 材料与方法
1.1 实验动物
实验中华蟾蜍幼体(蝌蚪和变态期)采自辽宁省沈阳市郊区。亚成体和成体中华蟾蜍购自沈阳医学院。以王昌对中华蟾蜍胚胎发育分期为标准[6],选用鳃盖完全愈合期、圆锤状肢芽期、肛管退化期、变态完成期、亚成体期以及成体期6个典型时期。发育各期中华蟾蜍各5例。
1.2 自发放电的测定
应用微电极电生理技术,对中华蟾蜍胚后端脑原始大脑皮层神经元的自发放电活动进行在体胞外记录。常规手术方法暴露不同发育时期蟾蜍的端脑并剥离硬脑膜。用微电极推进器(西北光学仪器厂,WF-2型)将尖端直径小于1μm、内充3moL KCl、阻抗5~20mΩ的玻璃微电极插入原始大脑皮层中。放电信号经由高阻微电极放大器(成都仪器厂,WF-IB型)后进入信号采集处理系统(成都仪器厂,RM6240型),在计算机上观察并记录原始大脑皮层神经元的电活动。
1.3 数据统计分析
实验结果用Microsoft Excel和SPSS 16.0软件统计分析并作图,各组之间的差异性采用单因素方差分析(One Way ANOVA)和Tukey's多重比较,文中数据以平均值±标准差表示(mean±SD),P<0.05即认为差异显著。
2 结 果
2.1 胚后各阶段原始大脑皮层神经元的自发放电
实验共记录到5种自发放电类型,分别为单个放电、连续放电、簇状放电、连续簇状放电和不规则放电(图1),在不同的发育时期各类型放电单位所占比例不同,见图2(图中1~6分别是鳃盖完全愈合期、圆锤状肢芽期、肛管退化期、变态完成期、亚成体期和成体期)。胚后发育的早期鳃盖完全愈合期和圆锤状肢芽期只有4种放电类型,没有记录到不规则放电。分别共记录到40和36个放电单位,分别以簇状放电和连续放电为主,为35%和50%。
胚后发育的后期,肛管退化期到成体均为5种放电类型,分别共记录到43、49、46和44个放电单位,以簇状和连续簇状放电为主要放电类型。肛管退化期以簇状放电为主,占34.88%;变态完成期则以簇状和连续簇状放电为主,分别是24.49%和26.53%。亚成体期以连续簇状放电和不规则放电为主,分别为28.26%和41.3%;成体期以簇状和连续簇状放电为主,分别为22.72%和31.82%。
2.2 胚后各阶段原始大脑皮层自发放电的电生理学特性
对中华蟾蜍胚后发育各阶段原始大脑皮层的单个和连续放电幅度以及连续单个和连续簇状放电频率进行了统计分析,结果见图3和图4(图中1~6分别是鳃盖完全愈合期、圆锤状肢芽期、肛管退化期、变态完成期、亚成体期和成体期;a、b、c、d表示差异显著性,同一放电类型不同发育时期的比较)。图3表明,单个放电振幅变化不大,仅肛管退化期存在减小,显著差异(P<0.05);连续放电幅度在胚后发育的中期均高于早期和成体期,且差异显著(P<0.05)。图3表明,两种放电频率均随着发育时期而降低,但在亚成体期再次升高,且连续单个放电较连续簇状放电变化显著。
我们还观察测量了每个放电单位的放电持续时间(表1),结果发现,单个放电的动作电位时程在变态期开始缩短,为早期的一半,且差异显著(P<0.05);连续放电和簇状放电的持续时间变化趋势相似,随着原始大脑皮层的发育,放电持续时间延长,但分别在亚成体期和变态期出现缩短后再次增加;连续簇状放电在变态期和亚成体期明显延长,不规则放电在成体期时间最长。
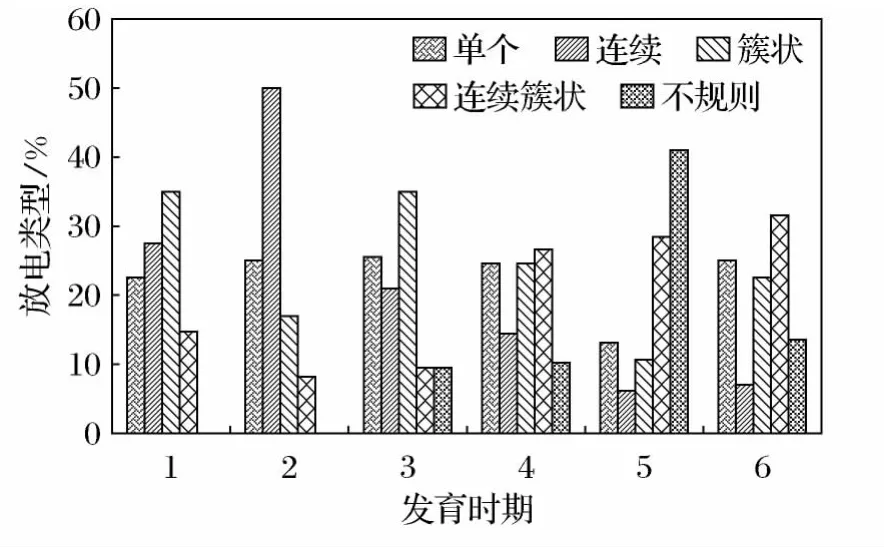
图2 中华蟾蜍不同发育时期原始大脑皮层各放电类型的放电单位比较
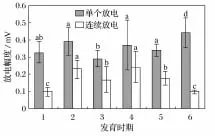
图3 中华蟾蜍不同发育时期原始大脑大脑皮层2种放电类型的放电幅度
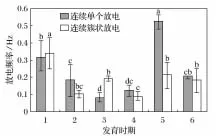
图4 华蟾蜍不同发育阶段原始大脑大脑皮层2种放电类型的放电频率
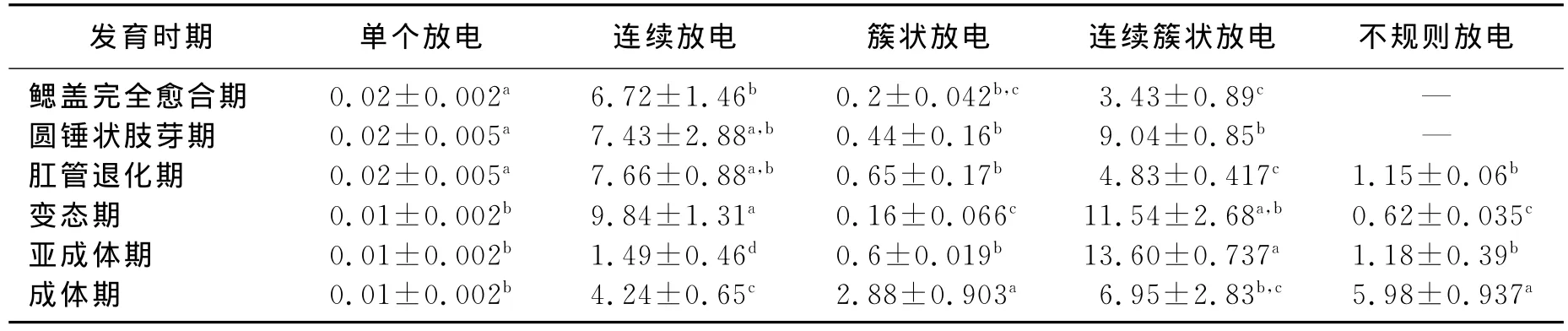
表1 中华蟾蜍胚后各阶段原始大脑皮层自发放电的出现时间 s
3 讨 论
两栖类原始大脑皮层位于端脑背侧,侧脑室上方,主要司嗅觉作用,但已是机体机能协调和整合活动的原始中枢[7]。
实验结果表明,从圆锤状肢芽期到成体期,连续簇状放电占总放电数量的比例逐渐增加,说明某些特异神经元兴奋性可能随发育的进行呈增强的趋势。变态完成期和成体期单个放电、簇状放电、不规则放电所占比例类似,这一结果可能说明变态期时原始大脑皮层的发育程度已经接近成体期。连续放电所占比例的减少可能和发育过程中神经元的凋亡有关[5,8]。
另外,中华蟾蜍端脑原始大脑皮层的电生理特性可能与神经细胞的兴奋性程度有关。在中华蟾蜍发育的较早阶段,功能性神经细胞数量较少,神经系统的功能尚未完全形成,因此细胞兴奋性较低,个别放电类型的数量较少,频率较低,单个放电的时程较长,为一种慢去极化放电[9]。伴随着发育的进行,细胞增殖、分化,功能性神经元的兴奋性增加,细胞放电类型、频率和数量均有增加的趋势,单个放电时程缩短,成为快去极化放电,表明神经元的兴奋性提高。神经系统发育过程中神经细胞的增殖、迁移、限定、分化、建立突触联系,从而产生生理功能。在这一系列的过程中,大约有20%~90%的神经元发生了退化、衰退和死亡。神经细胞在发育过程中的增殖和退化可能是导致某些自发放电形式的数量或频率出现差异的原因[10-11]。
[1]刘明玉,解玉浩,季达明,等.中国脊椎动物大全[M].沈阳,辽宁大学出版社,1999:477-478.
[2]HOFFMAN H R.The olfactory bulb,accessory olfactory bulb,and hemisphere of some anurans[J].J Comp Neurol,1963,120:318-368.
[3]王畅,毕远阁,王欢欢,等.亚成体中华蟾蜍端脑形态学与组织学的初步研究[J].四川动物,2009,28(1):35-39.
[4]ZHOU Yingying,SHAO Ran,LIANG Chuancheng,et al.Histological studies on the telencephalon of Hynobius leechiiat the metamorphosis phase and the adult phase[J].Neurosci Bull,2009,25(3):196-202.
[5]WANG Huanhuan,LI Liyan,WANG Liwen,et al.Morphological and histological studies on the telencephalon of the salamander Onychodactylus fischeri[J].Neurosci Bull,2007,23(3):170-174.
[6]王昌,贾杏珍,李琰,等.中华大蟾蜍的胚胎发育的初步观察[J].两栖爬行动物学报,1984,3(2):39-48.
[7]解谦.脊椎动物从水生到陆生的结构演变[J].山西农业大学学报,2003,23(4):384-386.
[8]NARUSE I,KEINO H.Apoptosis in the developing CNS[J].Prog Neurobiol,1995,47(2):135.
[9]LIANG Chuancheng,BIAN Shaokang,PENG Xia,et al.Studies on spontaneous firing in olfactory bulb of Bufo bufo gargarizans in hibernation and after hibernation[J].NRR,2011,6(11):865-869.
[10]NORDGREEN J,HORSBERG E,RANHEIM B,et al.Somatosensory evoked potentials in the telencephalon of Atlantic salmon (Salmo salar)following galvanic stimulation of the tail[J].J Comp Physiol A,2007,193:1235-1242.
[11]LENG Jie,JIANG Li,CHENG Hengsheng.Electrophysiological characteristics of early differentiated neural stem cells in embryonic rats[J].Acta Acad Med,2010,32(3):238-241.
猜你喜欢
红蜻蜓·低年级(2021年6期)2021-08-23
红蜻蜓·低年级(2021年6期)2021-07-21
当代水产(2021年5期)2021-07-21
读友·少年文学(清雅版)(2020年12期)2020-05-21
科普童话·百科探秘(2020年3期)2020-05-11
湖南畜牧兽医(2020年6期)2020-03-22
水利科技与经济(2016年2期)2016-04-09
中学生物学(2008年12期)2008-12-27
祝您健康(1991年1期)1991-12-30
