
大豆孢囊线虫不同寄主种群的形态变异及对大豆寄主的适应性差异分析
2014-08-10 12:29王慧敏赵洪海王凤龙
植物保护 2014年6期
王慧敏, 赵洪海*, 王凤龙
(1.青岛农业大学农学与植物保护学院,山东植物病虫害综合防控重点实验室,青岛 266109;2.中国农业科学院烟草研究所,青岛 266101)
大豆孢囊线虫不同寄主种群的形态变异及对大豆寄主的适应性差异分析
王慧敏1, 赵洪海1*, 王凤龙2
(1.青岛农业大学农学与植物保护学院,山东植物病虫害综合防控重点实验室,青岛 266109;2.中国农业科学院烟草研究所,青岛 266101)
在对大豆、烟草与地黄寄主上14 个大豆孢囊线虫(SCN)种群形态与分子鉴定的基础上,对雌虫的孢囊和阴门锥进行形态观察和测计。结果表明:烟草SCN和大豆SCN的各测量值之间无明显差异(P>0.05),地黄SCN的阴门窗长(43.8 μm)和宽(33.9 μm)显著小于其他两者(P<0.05);同一寄主不同SCN种群之间的形态也存在一定差异;阴门裂长和下桥长具有稳定性,可以作为鉴定大豆孢囊线虫可靠的形态特征。3 种寄主种群侵染大豆(‘荷豆12号’)的试验表明,大豆SCN能侵染大豆并能形成雌虫,而烟草SCN和地黄SCN未能侵染大豆和形成雌虫。本研究结果表明SCN不同寄主种群存在形态变异,而它们对大豆的寄主适应性也不相同。
大豆孢囊线虫; 形态变异; 分子鉴定; 寄主适应性
大豆孢囊线虫(HeteroderaglycinesIchinohe)(soybean cyst nematode, 简称SCN)是一种雌雄异形、两性生殖的定居性内寄生线虫,1899 年首次在我国东北地区被发现,目前已遍布黑龙江、吉林、辽宁、内蒙古、河北、河南、山东、山西、陕西、安徽、北京、江苏、湖北、上海、浙江等15 个省(直辖市、自治区)[1-2]。大豆孢囊线虫病是世界上危害大豆最重要的毁灭性病害之一[3]。
SCN的寄主范围较窄,主要寄生在豆科植物上,如大豆、大果田菁、胡枝子、绿豆、赤豆和菜豆等[4]。此外,该线虫也可寄生在非豆科植物上,如玄参科的地黄、黄花毛蕊花和比利牛斯金鱼草,唇形科的高黄芩,杂草类的宝盖草和荠菜等[5]。在我国,已报道的寄主植物除豆科植物外,还有地黄[6]、芝麻[7]、泡桐[8]和烟草[9]。线虫的形态在种内会存在一定的差异,这种差异可能是由遗传差异、寄主因素、地理起源等多种因素造成[10-12]。对于SCN,不同地理种群间的形态变异已有报道[13-14],但是有关寄主植物影响形态变异的研究未见报道。此外,SCN可表现出不同的寄主适应性,如菜豆品系‘G122’对美国田纳西州大豆上SCN的3 号生理小种高感,却对同样来自田纳西州大豆上SCN的5号和14 号生理小种表现出抗性[15]。
为了确定寄主对大豆孢囊线虫的形态分化及适应性的影响,笔者从烟草、大豆、地黄3 种寄主上分离到14 个大豆孢囊线虫种群(下文简称烟草SCN,大豆SCN和地黄SCN),对其形态变异和寄主适应性进行了研究。
1 材料与方法
1.1 线虫的采集
于2013 年在中国3 省8 市共采集14 份SCN土壤样品,具体采集地点、寄主及编号见表1。采用对角线5 点取样法,在种有寄主植物的地块上选定5 个点,用铁锹铲去较干燥的3~5 cm表层土,每个点挖取根及根围土壤500 g作为一个小样,将5 个点采集到的小样混合作为1 份土壤样品。
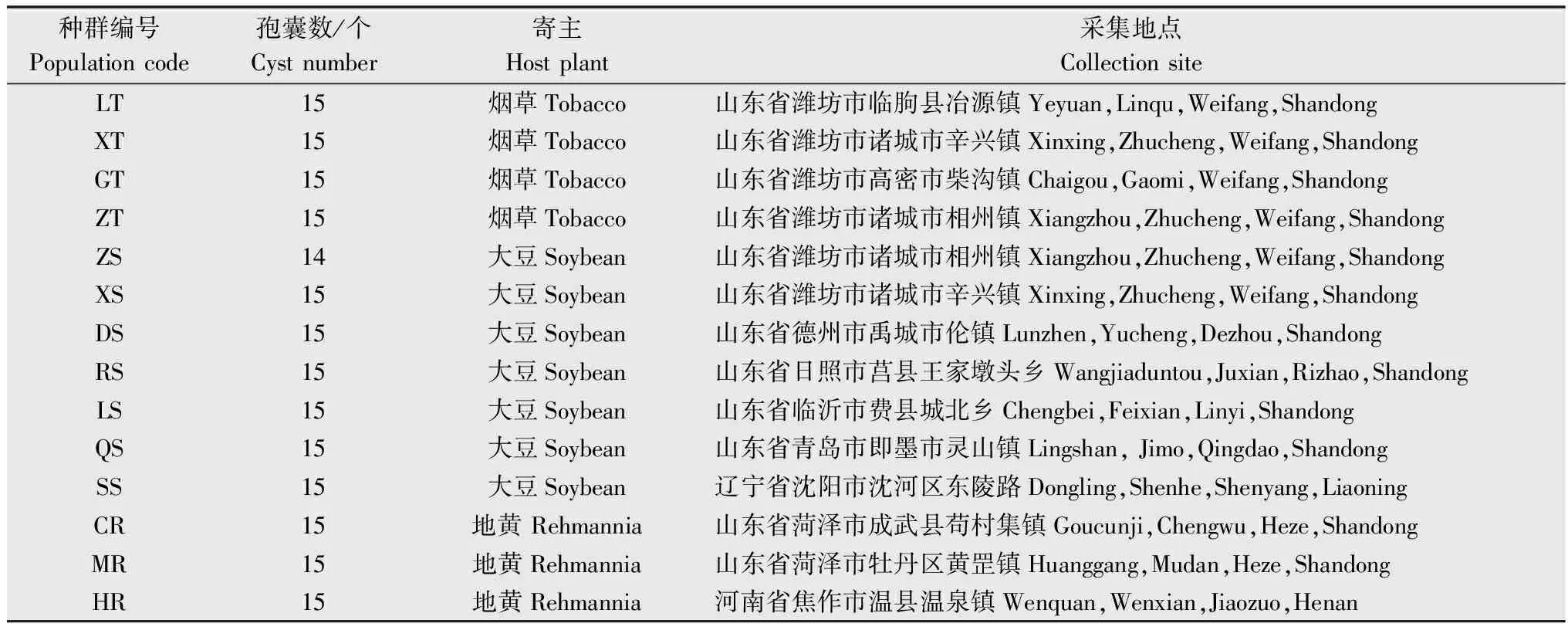
表1 不同寄主的孢囊线虫种群样品Table 1 The samples of cyst nematode populations on different host plants
1.2 基于形态学与rDNA-ITS基因的物种鉴定
利用直接过筛法分离土中的孢囊,在解剖镜下挑取饱满的孢囊放在-4 ℃冰箱内冷藏备用。切取孢囊的阴门锥部分制成玻片,直接过筛法及孢囊阴门锥的制备参照刘维志的方法[16]。
单个孢囊线虫DNA的提取参考Ou等的方法[17],PCR扩增引物采用ITS通用引物,AB28(5′-ATATGCTTAAGTTCAGCGGGT-3′)和TW81(5′-GTTTCCGTAGGTGAACCTGC-3′)[18]。采用50 μL的反应体系,配比如下:10×PCR buffer(含Mg2+)5 μL,dNTP 1 μL,AB28(10 μmol/L)3 μL, TW81(10 μmol/L)3 μL,TaqDNA多聚酶0.5 μL,模板DNA 3 μL,最后加入ddH2O补足至50 μL。扩增条件为:94 ℃预变性5 min;94 ℃变性45 s,55 ℃退火1 min,72 ℃延伸2 min,35 个循环;最后72 ℃延伸10 min,在4 ℃下保存。取7 μL PCR产物经1%琼脂糖凝胶电泳后,用EB染色,在紫外灯下成像检测。选取有条带的PCR产物送至上海生工进行双向测序,测序结果与GenBank中收录的序列进行BLAST比对(登录号为KJ161289~KJ161302)。从GenBank中下载不同SCN种群的ITS序列并筛选出所有单倍型,以印度和叙利亚的小麦孢囊线虫(H.avenae)和意大利的烟草球孢囊线虫(Globoderatabacum)为外群(表2),采用neighbor-joining(NJ)法(MEGA5.0)构建系统发育树,系统发育树各分支的置信度(bootstrap)均进行1 000 次重复检验。

表2 GenBank 数据库下载的SCN的ITS参考序列Table 2 Reference ITS sequences of SCNdownloaded from GenBank database
1.3 形态差异分析
在光学显微镜下观察孢囊阴门锥的形态特征并拍照,对孢囊线虫的重要分类特征进行测量,包括孢囊的长与宽、阴门裂长、阴门窗长、阴门窗宽、下桥长。每个种群孢囊测量数量见表1,每个孢囊重复测量3次,记平均值。对测量的原始数据,用Excel 2003软件计算各组数据的平均数、标准差和标准误。用SPSS 19软件的One-way ANOVA程序的LSD多重比较法对SCN不同寄主种群间及同一寄主种群间的形态进行差异显著性分析,显著水平为0.05。
1.4 三种寄主种群对大豆寄主适应性测定
在直径为8 cm的盆栽容器中放入260 g土(50%灭菌土壤+50%灭菌沙子),每个容器中撒入2 粒大豆种子(‘荷豆12 号’),将容器放在28 ℃[19](L∥D=16 h∥8 h)的温室中,大约1 周后,待大豆种子长成幼苗后,每盆留取1 株健康的大豆苗,并且保证所有供试的大豆苗长势相同,然后开始准备接种卵。将孢囊破裂后收集卵,制成卵悬浮液。向大豆苗盆中分别接种采集自大豆XS种群、烟草XT种群和地黄CR种群的线虫卵,在盆内靠近大豆苗的四周挖出深4 cm的小孔,将卵悬浮液倒入4 个孔内,每盆接种2 000 粒卵,试验重复3 次[20]。接种30 d后,将大豆连根拔出,采用过筛法和过筛-贝曼漏斗法分离并测定整盆土壤中的孢囊数目和2龄幼虫(J2)数量;采用次氯酸钠-酸性品红染色法检测大豆根系受侵染情况[16]。
2 结果
2.1 物种形态鉴定与分子鉴定
对采自烟草、大豆和地黄上的14 个供试群体的孢囊进行观察,发现其主要形态特征与大豆孢囊线虫完全一致,所有群体的孢囊都为柠檬形,深褐色;有明显的颈和突出的阴门锥,阴门锥附近有较多排列不规则的泡状突,下桥发达,膜孔为双半膜孔结构;每个孢囊内有大约100~200 个卵。通过形态学观察和形态测量值的比较,将3 种寄主上的14 个供试群体的孢囊线虫鉴定为大豆孢囊线虫(HeteroderaglycinesIchinohe,1952)。
电泳结果显示,14 个供试群体的SCN均能扩增出1 条约1 000 bp的条带。测序、比对后构建的系统发育树(图1)显示,14 个供试种群的特异性片段与来自美国、加拿大、伊朗及我国云南、甘肃、宁夏、山东、安徽、河南、河北和内蒙古SCN种群聚在一个分支上(遗传距离 ≤ 0.03),置信度达到99%;与来自印度的H.avenae种群(KC152906)和叙利亚的H.avenae种群(JX024198)的亲缘关系较远(遗传距离为0.24~0.44),而烟草上常见的球孢囊属线虫G.tabacum(HQ260403)则处于另一进化分支。

图1 用NJ法构建的大豆孢囊线虫及其近缘种的rDNA-ITS序列系统发育树Fig.1 Phylogenetic tree of H. glycines and the closely related species based on rDNA-ITS sequences using NJ method
2.2 SCN不同寄主种群间形态差异
从形态测量值的统计分析来看(表3),烟草SCN和大豆SCN在孢囊形态上无显著性差异;地黄SCN在体长、体宽、阴门裂长和下桥长4 个测量值上与大豆SCN和烟草SCN无显著性差异,在阴门窗长(43.8 μm)和阴门窗宽(33.9 μm)上显著小于大豆SCN(分别为49.4 μm和35.7 μm)和烟草SCN(分别为47.4 μm和35.6 μm)(P< 0.05)。
2.3 SCN同一寄主不同种群间形态差异
从表4中可看出,烟草SCN LT种群体宽(429.2 μm)最大,阴门窗长(38.0 μm)和阴门窗宽(30.9 μm)最小,与其他3个同寄主种群XT、GT和ZT存在显著性差异(P< 0.05)。
大豆SCN的QS种群体长(554.7 μm)最小,与其他大豆种群差异显著(P< 0.05);种群SS的体宽(425.2 μm)最大,只与种群XS无显著性差异;种群DS和QS体宽最小(分别为361.7和364.1 μm),与种群SS和XS差异显著(P< 0.05);种群XS和DS的阴门窗长(分别为43.1和44.2 μm)显著小于其他寄主种群(P< 0.05);种群SS、XS和DS的阴门窗宽(分别为33.8、33.7和33.3 μm)显著小于其他寄主种群(P< 0.05)。
地黄SCN的HR、CR 2个种群在孢囊形态上无显著差异;种群MR与HR和CR种群存在显著性差异(P< 0.05),其中体长(614.7 μm)和阴门窗长(35.6 μm)均大于其他两者。
2.4 三种寄主种群的SCN侵染大豆后的生长发育情况
接种30 d后发现,所有大豆盆土中均有孵出的2龄幼虫,但大豆SCN种群孵出的2龄幼虫数量(1 268.7条/盆)远远高于烟草SCN和地黄SCN(分别为772.7 条/盆和109.7 条/盆)(P< 0.05);在所有接种SCN的盆土中均未发现成熟孢囊。大豆根系内线虫染色检查发现,在接种大豆SCN的大豆根内有J2(6.4 条/株根)和雌虫(1.3 个/株根),而在接种烟草SCN和地黄SCN的大豆根内未发现SCN的任何虫态(图2)。

图2 三种寄主SCN种群接种大豆后的表现Fig.2 Performance of the three host SCN populations after inoculation
3 结论与讨论
陈金堂等[6]与程子超等[9]分别首次报道地黄和烟草上的孢囊线虫为SCN。本研究通过形态学鉴定与rDNA-ITS序列分析,将采自烟草、大豆和地黄上的14 个孢囊种群鉴定为SCN。将本次形态测量数据与前人的测量数据进行比较,发现大豆SCN的阴门窗长,烟草SCN的体长和体宽,地黄SCN的体长、体宽、阴门裂长和阴门窗长比前人的测量数据偏小。究其原因,可能与其寄主种类、营养状况和生存环境等有关。

表3 不同寄主上的大豆孢囊线虫形态测量值1)Table 3 Morphological measurements of H. glycines from different hosts
1) 不同小写字母表示处理间差异显著(P<0.05)。下同。
Different small letters indicated significant difference among different treatments at 0.05 level. The same below.

表4 不同地理群体的大豆孢囊线虫的形态测量值Table 4 Morphological measurements of different geographical SCN populations
对3种不同寄主上的大豆孢囊线虫的6个形态指标进行测量和统计分析,结果表明烟草SCN和大豆SCN各测量值之间无明显差异,而地黄SCN阴门窗长和阴门窗宽显著小于其他两者,说明寄主对大豆孢囊线虫形态分化存在一定的影响。同一寄主不同大豆孢囊线虫种群之间的形态值也存在着一定的差异,说明了大豆孢囊线虫形态的多样性和不稳定性。14个不同群体之间的阴门裂长和下桥长无显著性差异,可以将两者作为鉴定大豆孢囊线虫的稳定和可靠的形态特征。
3种寄主种群侵染大豆的试验表明,大豆SCN的2龄幼虫能侵染大豆并较好地在大豆上繁殖,而烟草SCN和地黄SCN虽然能在大豆苗土中孵化出2龄幼虫,但是数量较少,且不能侵染大豆根部和形成雌虫,说明寄主转换后新寄主对SCN的孵化和侵入产生抑制作用,SCN对新寄主的适应能力不强。对于此结果有2种原因可以解释,一是适宜寄主植物的根系分泌物对SCN卵的孵化刺激较大,非适宜寄主植物对SCN卵的孵化刺激较小[21],因而大豆SCN能在大豆上产生较多的2龄幼虫进而更容易有机会侵染大豆,而地黄和烟草SCN孵出2龄幼虫数量较少,侵入根系的机会减少;二是烟草SCN和地黄SCN对大豆无致病性,原来发生在大豆上的SCN在对烟草和地黄的适应过程中致病基因发生某种改变,但是这种改变如何变化还不得而知。
[1]许艳丽,王丽芳,战丽莉.大豆胞囊线虫病研究进展(续一)[J].大豆科技,2010(1):21-24.
[2]Zheng J,Zhang Y,Li X,et al.First report of the soybean cyst nematode,Heteroderaglycines,on soybean in Zhejiang,eastern China[J].Plant Disease,2009,93(3):319.
[3]Li X X,Wu H Y,Shi L B,et al.Comparative studies on some physiological and biochemical characters in white and brown cysts ofHeteroderaglycinesrace 4[J].Nematology,2009,11(3):465-470.
[4]Heydari R,Pourjam E,Maafi Z T,et al.Comparative host suitability of common bean cultivars to the soybean cyst nematode,Heteroderaglycines,in Iran[J].Nematology,2010,12(3):335-341.
[5]Venkatesh R,Harrison S K,Riedel R M.Weed hosts of soybean cyst nematode(Heteroderaglycines)in Ohio[J].Weed Technology,2000,14(1):156-160.
[6]陈金堂,李知.为害地黄的大豆孢囊线虫的初步研究[J].植物病理学报,1981,11(1):37-44.
[7]山西省大豆孢囊线虫病调查协作组.大豆孢囊线虫病在山西省的发生分布、为害程度和寄主范围[J].华北农学报,1987,2(3):74-79.
[8]张东升.大豆胞囊线虫侵染泡桐和豌豆的研究[J].植物病理学报,1995,25(3):275-278.
[9]程子超,赵洪海,李建立,等.山东省寄生烟草的孢囊线虫种类鉴定及种内群体间rDNA-ITS-RFLP分析[J].植物病理学报,2012,42(4):387-395.
[10]黄健,戚龙君,王金成,等.腐烂茎线虫种内不同群体形态及遗传分析[J].植物病理学报,2009,39(2):125-131.
[11]Ye W,Robbins R T.Morphological observation onLongidoruscrassusThorne,1974(Nematoda:Longidoridae)and its intraspecies variation[J].Journal of Nematology,2005,37(1):83-93.
[12]Stock S P,Mracek Z,Webster J M.Morphological variation between allopatric populations ofSteinernemakraussei(Steiner,1923)(Rhabditida:Steinernematidae)[J].Nematology,2000,2(2):143-152.
[13]Faghihi J,Ferris J M,Ferris V R.Heteroderaglycinesin Indiana:II.Morphology of geographical isolates[J].Journal of Nematology,1986,18(2):173-177.
[14]Lax P,Doucet M E.HeteroderaglycinesIchinohe,1952(Nematoda:Tylenchida)from Argentina.4.Morphological and morphometrical characterisation of cysts[J].Nematology,2002,4(7):783-789.
[15]Smith J R,Young L D.Host suitability of diverse lines ofPhaseolusvulgaristo multiple populations ofHeteroderaglycines[J].Journal of Nematology,2003,35(1):23-28.
[16]刘维志.中国检疫性植物线虫[M].北京:中国农业科学技术出版社,2004:72-117.
[17]Ou S,Peng D,Liu X,Li Y,et al.Identification ofHeteroderaglycinesusing PCR with sequence characterised amplified region(SCAR)primers[J].Nematology,2008,10(3):397-403.
[18]Zheng J,Subbotin S A,Waeyenberge L,et al.Molecular characterisation of ChineseHeteroderaglycinesandH.avenaepopulations based on RFLPs and sequences of rDNA-ITS regions[J].Russian Journal of Nematology,2000,8(2):109-113.
[19]赵洪海,程子超,王凤龙.山东省烟草孢囊线虫的群体动态和世代发生特点[J].植物保护学报,2013,40(6):529-532.
[20]Masler E P,Rogers S T.Effects of cyst components and low temperature exposure ofHeteroderaglycineseggs on juvenile hatchinginvitro[J].Nematology,2011,13(7):837-844.
[21]吴海燕,段玉玺,李秀侠,等.SCN侵染对不同大豆品种根系氨基酸/低分子肽类分泌物的影响[J].植物病理学报,2007,37(6):616-622.
MorphologicalvariationofHeteroderaglycinesfromdifferenthostsandtheiradaptabilitytosoybean
Wang Huimin1, Zhao Honghai1, Wang Fenglong2
(1.CollegeofAgronomyandPlantProtection,QingdaoAgriculturalUniversity,KeyLaboratoryofIntegratedCropPestManagementofShandongProvince,Qingdao266109,China; 2.TobaccoResearchInstitute,ChineseAcademyofAgriculturalSciences,Qingdao266101,China)
Based on both morphological and molecular identification of 14 soybean cyst nematode (SCN) populations on soybean, tobacco and rehmannia, cyst and vulval cone were characterized by morphological observation and morphometry. The results showed that SCN populations collected from tobacco and soybean had no obvious difference in morphological measurements (P>0.05), but the fenestral length (43.8 μm)and width (33.9 μm)in the population collected from rehmannia were obviously shorter than those of SCN from other two hosts (P<0.05). There were some morphological differences in different SCN populations collected from the same host. The length of vulval slit and underbridge could be used as reliable identification characters for their high stability.SCN populations collected from soybean could infect soybean cultivar ‘Hedou 12’ and produced females; however,the populations collected from tobacco and rehmannia failed to infect the soybean cultivar or formed females. The results indicated that there were variations in morphological characters and host adaptability of different populations of SCN.
Heteroderaglycines; morphological variation; molecular identification; host adaptability
2014-03-19
:2014-05-12
中国烟草总公司科技项目(110200902065);公益性行业(农业)科研专项(200903040);山东省“泰山学者”建设工程专项
S 435.651
:ADOI:10.3969/j.issn.0529-1542.2014.06.019
致谢:沈阳农业大学段玉玺教授和陈立杰教授为本研究提供辽宁省大豆孢囊线虫样品,青岛农业大学植病专业硕士研究生王磊磊和李志辉参加部分调查工作,在此表示感谢。
* 通信作者 E-mail:hhzhao@qau.edu.cn
猜你喜欢
畜牧兽医学报(2022年9期)2022-09-30
今日农业(2022年14期)2022-09-15
热带海洋学报(2022年4期)2022-08-08
暨南大学学报(自然科学与医学版)(2022年3期)2022-06-16
中国乳业(2020年12期)2020-01-16
中国森林病虫(2019年5期)2019-10-11
中国畜禽种业(2019年9期)2019-01-06
中成药(2018年2期)2018-05-09
植物保护(2017年6期)2017-11-29
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
