
水曲柳不同颜色细根生理功能变化
2014-08-15 09:41汤园园
安徽农业科学 2014年18期
卫 星,汤园园,王 婧
(东北林业大学林学院,黑龙江哈尔滨 150040)
根系生理功能变化是根系代谢、能量转化和生长发育等生命现象的直接表现。细根是吸收养分和水分的重要器官,在不同阶段发育,细根表皮和皮层组织细胞的形态变化及内含物的积累会导致细根颜色发生改变[1]。不同颜色细根生理功能存在着一定的差异。Cruz等[2]研究角豆树(Ceratonia siliqua)发现,白色细根吸收氮的速率是黄色根的1.0倍、棕色根1.5 倍,细根颜色越深,吸收速率越低[2]。Comas等[3]也发现,白色细根呼吸速率最强,其次是棕色细根,褐色皱缩细根呼吸速率最低。笔者在之前的研究中也发现干旱胁迫下水曲柳苗木不同颜色细根随根序升高,颜色加深,根系活力下降,质膜透性增加[4]。可见细根颜色与生理功能具有明显的相关性,研究不同颜色细根的生理功能对于准确评价细根寿命及其在生态系统能量流动中的作用具有重要的意义。
水曲柳(Fraxinus mandshurica)是东北地区主要的造林和用材树种。了解其复杂的根系系统对研究我国北方森林生态系统地下物质和能量流动具有重要的意义。虽然目前已经开展了水曲柳细根生物量[5-6]、死亡和周转[7]、寿命[8]、土壤呼吸[9]、细根形态[10]、细根构型[11]以及细根分解[12-13]等方面的研究,但多数研究均以直径或级别定义细根,忽略了不同颜色细根的生理功能差异,不利于准确估计细根的生态效益。该研究对23年生水曲柳不同颜色细根全氮含量、质膜透性、呼吸速率和根活力的变化进行分析,以期为准确定义不同颜色水曲柳细根的生理功能提供依据。
1 材料与方法
1.1 试验材料 取样地点位于东北林业大学帽儿山实验林场尖砬沟森林培育实验站(127°30'~127°34'E,45°21'~45°25'N),该地属温带大陆性季风气候区,林分为23年生水曲柳人工林。2008年9月上旬,用内径为60 mm的根钻钻取土芯样品,用恒温箱低温保存带回实验室分析根样。按照Pregitzer等[14]的方法先将水曲柳1~3级根不同颜色的细根分开,再进行生理指标分析。
1.2 研究方法 全氮含量分析采用Foss全自动凯氏定氮仪进行测定;呼吸速率采用气相氧电极法(model LD 2/3,Hansatech,England)测定[15];质膜透性采用相对电导率法[16]测定;根系活力采用TTC染色法[17]测定。
1.3 数据分析 每个指标采用SPSS软件包(v11010,SPSS Inc,Chicago,Illinois,USA)进行方差分析,然后进行 LSD 检验,采用SigmaPlot(Version 9101,USA)软件进行示图制作。
2 结果与分析
2.1 不同颜色细根的全氮含量变化 水曲柳细根随着根序升高,全氮含量下降,不同颜色的细根全氮含量存在较大差异(图1)。在1级根中,细根全氮含量是随着颜色加深而呈现上升趋势,褐色根全氮含量最高,其次是皱缩根,白色根含量最低。褐色根和褐色皱缩根含量的变化不显著。在2级根中,褐色根全氮含量也是最高,皱缩根含量最低,4种颜色根全氮含量之间差异显著(P<0.05)。在3级根中,细根全氮含量随着颜色的加深逐渐下降,但褐色根和褐色皱缩根的下降趋势变缓,两者之间差异不显著(P>0.05),白色和黄色根之间差异显著(P<0.05)。
2.2 不同颜色细根的质膜透性 质膜透性表征细胞结构的完整性。研究发现,同一级别内各颜色根质膜透性均差异极显著(P<0.01)。随颜色的加深,根系的质膜透性极显著增加(P<0.01),1~3级根表现了相似的变化规律。在同一根序中,白色根质膜透性最小,皱缩根质膜透性最大,为白色根的4~5倍。但不同级别中相同颜色根质膜透性差异不显著(P >0.05)。

图1 水曲柳不同颜色细根全氮含量的变化

图2 水曲柳不同颜色细根质膜透性的变化
2.3 不同颜色细根的呼吸作用 根系通过呼吸作用为根生物量合成、维持和离子吸收等提供能量。研究发现1级细根皱缩根呼吸速率最低,其次是褐色根,黄色根呼吸速率最高(图3)。在2种温度条件下都具有相同的变化规律。相同的温度条件下,皱缩根呼吸速率与其他3种颜色根的呼吸速率之间差异显著(P<0.05),但是,白色根、黄色根和褐色根之间差异不显著(P>0.05)。
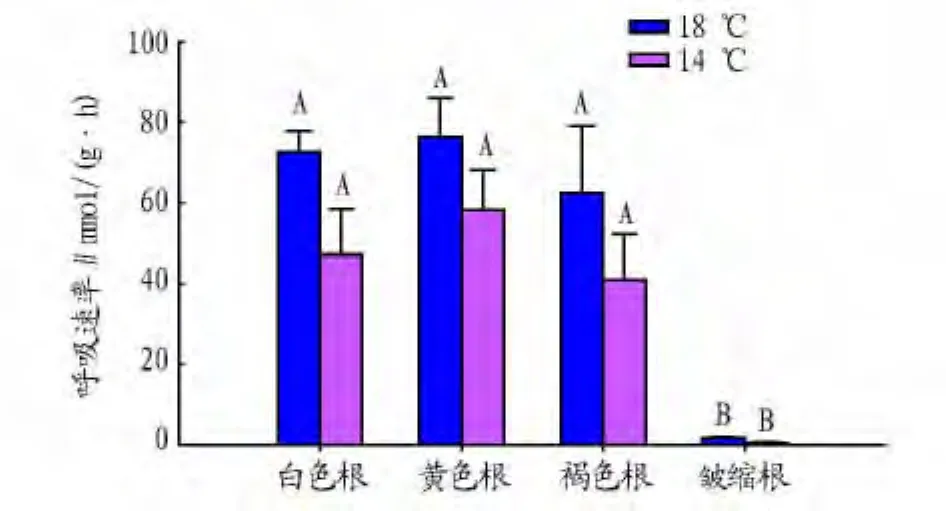
图3 水曲柳不同颜色1级根呼吸速率
2.4 不同颜色细根的根系活力 根系活力降低是细根衰老死亡的最早生理表现形式之一。研究发现,1级根活力随其颜色的加深呈现出先升高后下降的趋势(图4)。不同颜色根系活力具有显著的差异(P<0.05),其中黄色细根活力显著高于其他颜色细根(P<0.05),为皱缩根的7倍左右,白色根活力也显著高于皱缩根(P<0.05),约为褐色根的4倍。皱缩根的细根活力最低,但是与褐色根之间差异不显著(P>0.05)。
3 结论与讨论
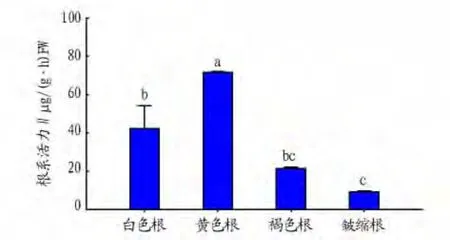
图4 水曲柳不同颜色1级细根活力
(1)颜色是根系形态发育的主要外在表现之一。目前关于根系颜色加深的原因主要有2种观点。1种观点认为颜色加深是细根成熟的标志,细胞完整性没有破坏,深色的根上仍然会长出新的根[18];另1种观点则认为细根颜色加深是衰老死亡的外在表现,根细胞大量解体[19]。随着细根生理生态功能研究的深入,发现在细根发育的不同阶段,可能存在这2种颜色变化机理。笔者曾研究干旱胁迫下水曲柳苗木细根颜色变化时发现细根从白色变化到黄色,根内部结构形态完整,是成熟的表现;根系颜色从黄色到褐色乃至黑色时,细胞程序化死亡,是细根衰老的表现[4]。
(2)质膜透性能直接反映根细胞的完整性,当细胞发生衰老死亡时质膜透性明显增加。该研究发现随颜色加深,细根质膜透性增加,黄色根质膜透性虽然高于白色根,但黄色根仍然具有很高的根系活力和呼吸速率。由此可见黄色根是吸收功能完整、比白色根要成熟的根。
(3)根呼吸速率高低代表了根系生理活动的强弱。根系呼吸速率高低代表其综合生理代谢强弱[3,20]。Pregitzer等[15]在研究北美糖槭林(Acer saccharum)时发现,细根随着组织氮浓度增加,呼吸速率呈线性关系(R2=0.69)。贾淑霞等[21]对水曲柳和落叶松前5级根研究也发现,细根呼吸速率与其氮含量线性相关(分别为R2=0.89和R2=0.97)。该研究发现褐色根和皱缩根呼吸速率和根活力均很低,说明褐色及皱缩可能是根衰老死亡过程中的外在表现。这一分析可通过进一步的细胞解剖结构完整性研究加以论证。
(4)氮素是影响植物体代谢的重要元素之一,根组织氮素含量对维持呼吸发挥着重要的作用[20,22]。该研究发现水曲柳1级根氮含量最高,不同颜色之间的氮含量在前3级根中存在差异。在1级根中,白色和黄色根氮含量较低,而褐色和皱缩根较高,在2级根和3级根中,皱缩根氮含量最低。产生这种现象的主要原因可能是:①在1级根中,白色根为新生出的根,功能还不完善,氮的吸收和同化能力不强。深色根出生时间较早,发育相对成熟,氮吸收和同化能力较高。Cruz等[2]研究发现,在养分丰富的条件下,黄色和褐色根吸收铵态氮速率较高。在该研究的林分中,土壤养分丰富,尤其是氮含量较高,这可能导致颜色较深的根全氮含量高于浅色的根。②菌根侵染通常会改变细根的颜色[3]。Cruz等[2]研究表明,被菌根侵染的细根氮含量是没有被菌根侵染细根氮含量的1倍,磷含量差异更大。水曲柳1级白色根还没有被真菌侵染,而颜色较深的根绝大多数可能被菌根侵染,菌根吸收氮的能力很强,因此,1级根中深色的根氮含量较高。③在2级根和3级根中,皱缩根皮层细根已经死亡,出现单宁等物质沉淀,主要剩下的是木质部细胞,因此氮含量较低。以上解释仅是从细根出生时间和菌根侵染等方面考虑,其解剖学和生理学机制还不清楚。
[1]WELLS C E,EISSENSTAT D M.Beyond the roots of young seedlings:the influence of age and order on fine root physiology[J].Journal of Plant Growth Regulation,2003,21:324 -334.
[2]CRUZ C,GREEN J J,WATSON C A,et al.Functional aspects of root architecture and mycorrhizal inoculation with respect to nutrient uptake capacity[J].Mycorrhiza,2004,14:177 -184.
[3]COMAS L,ESSENSTANT D M,LAKSO A.Assessing root death and root system dynamics in a study of grape canopy pruning[J].New Rhytologist,2000,147:171 -178.
[4]卫星.干旱胁迫对水曲柳苗木细根衰老的影响[D].哈尔滨:东北林业大学,2008.
[5]梅莉,王政权,韩有志,等.水曲柳根系生物量、比根长和根长密度的分布格局[J].应用生态学报,2006,17(1):1 -4.
[6]刘金梁,梅莉,谷加存,等.内生长法研究施氮肥对水曲柳和落叶松细根生物量和形态的影响[J].生态学杂志,2009,28(1):1 -6.
[7]史建伟,王政权,于水强,等.落叶松和水曲柳人工林细根生长、死亡和周转[J].植物生态学报,2007,31(2):333 -342.
[8]于水强,王政权,史建伟,等.水曲柳和落叶松细根寿命的估计[J].植物生态学报,2007,31(1):102 -109.
[9]贾淑霞,王政权,梅莉,等.施肥对落叶松和水曲柳人工林土壤呼吸的影响[J].植物生态学报,2007,31(3):372 -379.
[10]王向荣,王政权,韩有志,等.水曲柳和落叶松不同根序之间细根直径的变异研究[J].植物生态学报,2005,29(6):871 -877.
[11]王向荣,谷加存,梅莉,等.水曲柳和落叶松细根形态及母根与子根比例关系[J].生态学报,2006,26(6):1686 -1692.
[12]宋森,谷加存,全先奎,等.水曲柳和兴安落叶松人工林细根分解研究[J].植物生态学报,2008,32(6):1227 -1237.
[13]张秀娟,吴楚,梅莉,等.水曲柳和落叶松人工林根系分解与养分释放[J].应用生态学报,2006,17(8):1370 -1376.
[14]PREGITZER K S,DEFOREST J L,BURTON A J,et al.Fine root architecture of nine North American trees[J].Ecological Monographs,2002,72:293-309.
[15]PREGITZER K S,LASKOWSKI M J,BURTON A J,et al.Variation in sugar maple root respiration with root diameter and soil depth[J].Tree Physiology,1998,18:665 -670.
[16]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[17]STEPONKUS P L,LANPHEAR F O.Refinement of the triphenyl tetrazolium chloride method of determining cold injury[J].Plant Physiology,1967,42:1423 -1426.
[18]WELLS C E,EISSENSTAT D M.Marked differences in survivorship among apple roots of different diameters[J].Ecology,2001,82:882 -892.
[19]CHENG W,COLEMAN D C,BOX J E.Root dynamics,production and distribution in agroecosystems on the Georgia piedmont using minirhizotrons[J].J Appl Ecol,1990,27:592 -604.
[20]RYAN M G,HUBBARD R M,PONGRACIC S,et al.Foliage,fineroot,woody-tissue and stand respiration in Pinus radiatain relation to nitrogenstatus[J].Tree Physiology,1996,16(3):333 -343.
[21]贾淑霞,赵妍丽,丁国泉,等.落叶松和水曲柳不同根序细根形态结构、组织氮浓度与根呼吸的关系[J].植物学报,2010,45(2):174 -181.
[22]段永宏,徐程扬,周睿智.林木根系呼吸及形态对氮的反应研究概述[J].安徽农业科学,2008,36(3):1010-1013.
猜你喜欢
林业科技情报(2021年1期)2021-04-14
延边大学学报(自然科学版)(2019年2期)2019-10-08
森林工程(2018年4期)2018-08-04
广西林业科学(2016年2期)2016-03-20
生物加工过程(2015年2期)2015-11-11
化工生产与技术(2014年1期)2014-04-04
中国病理生理杂志(2014年6期)2014-01-26
华南农业大学学报(2012年1期)2012-11-10
食品工业科技(2011年12期)2011-11-02
