
伏牛山国家级自然保护区森林生态系统草本优势种空间分布格局
2014-09-26 09:26范玉龙丁圣彦卢训令
河南农业大学学报 2014年5期
范玉龙, 胡 楠, 丁圣彦, 卢训令
(1.河南大学环境与规划学院,河南 开封 475001;2.南阳理工学院,河南 南阳 473004)
伏牛山国家级自然保护区森林生态系统草本优势种空间分布格局
范玉龙1,2, 胡 楠1,2, 丁圣彦1, 卢训令1
(1.河南大学环境与规划学院,河南 开封 475001;2.南阳理工学院,河南 南阳 473004)
对伏牛山国家级自然保护区不同海拔梯度林下草本植被进行了调查,研究不同演替阶段草本优势种的空间分布格局及其动态变化.运用扩散系数、聚集指数、平均拥挤度、聚块性指数、Green指数等指标进行统计分析.结果表明,草本优势种多呈聚集分布,如羊胡子草(Carexlanceolata)、芒(Miscanthussinensis)、蕨(Pteridiumaquilinum)和野菊花(Dendranthemaindicum)等,呈随机分布的只有费菜(Sedumkamtschaticum),呈均匀分布的是苍术(Atractylodeslancea)和兔儿伞(Syneilesisaconitifolia).从演替初期到演替后期,呈聚集分布的优势种增多,大部分种群的空间分布格局指数的数值成倍地增加,有少数种群的空间格局类型发生了改变,如掐不齐(Kummerowiastriata)和蒿(Artemisiaargyi)从聚集分布变为随机分布.这些种群分布格局的变化,可能是群落稳定的一种外在表现.草本植物对环境变化的敏感性较强,对环境变动的响应有较明显的指示作用.
伏牛山自然保护区;草本优势种;空间格局;演替;森林生态系统
伏牛山自然保护区森林生态系统发育完整,优势种控制着森林生态系统的结构与功能.植被分层现象明显,乔木层、灌木层和草本层有各自的优势种.作为林下植被,草本植物能不断、快速地调整个体的形态、组织结构等自身特性,为整个森林生态系统的资源利用最大化、长期稳定共存起到不可替代的作用.种群分布格局是种群生物学特征的重要方面,个体、种群和群落的空间结构对生态过程非常重要[1],是物种与环境长期相互作用的产物.不同演替期森林生态系统林下草本植物分布格局的变动,可以反映出草本植物对环境变化的响应,也有助于理解整个森林生态系统的协同进化过程.种群分布格局一般可以分为随机分布、集群分布、均匀分布3种类型.随机分布很少出现,只有在环境因子在水平空间的分配位于许多种的生态幅之内,且对它们的作用程度近似相等时,或者环境极端恶劣时,或者某一主导因素成随机分布时,才会引起种群的随机分布.在较大的空间范围内生境的异质性较显著,只有在小面积上,生境因子的均匀性才可能得到满足,所以随机分布这种类型的尺度局限性非常明显.均匀分布在自然种群中极为少见.集群分布则是自然情况下常见的类型,与随机分布相比,个体有较高的超平均密度占据的范围[2].
种群空间格局的研究多以优势种为对象,以与人类生活密切相关的种群为重点,以不同环境因子对种群分布的影响为手段,以不同演替期的分布格局差异为比较等多种方法来分析种群空间分布格局的成因及其生态学意义.范娟等[3]研究了江西九连山亚热带常绿阔叶林优势种空间分布格局,用完全随机零模型不排除生境异质性时,4个优势种总体及其不同生长阶段在0~30 m所有尺度上主要呈现聚集分布.陈志伟等[4]对浙江省加拿大一枝黄花(Solidagocanadensis)的空间分布格局及其与人类活动的关系进行了研究,表明加拿大一枝黄花在浙江省的分布存在明显的空间异质性.杜丽侠等[5]对北京八达岭地区灌木群落空间分布格局与土壤环境因子的关系进行了分析,表明土壤环境因素对灌木空间分布格局影响较大.蒋雪琴等[6]对湖北神农架地区巴山冷杉(Abiesfargesii)种群结构特征与空间分布格局进行了研究,表明巴山冷杉种群不同年龄级的个体呈现出集群分布或随机分布.这些研究多以乔木优势种、灌木优势种或草地草本优势种为对象,而对森林生态系统林下草本植物的系统研究较少.草本植物作为森林生态系统的重要组成部分,受海拔、不同森林类型和森林不同盖度的影响较大,由于乔木和灌木在不同时空条件下发育程度不同,为草本植物提供了异质性的生存环境.同时,草本层对森林生态系统的结构和功能作用独特,对森林生态系统林下草本优势种的空间分布格局进行研究,可以理解一些细微的环境变动如何影响植物分布,也可理解草本植物如何弥补乔木、灌木在协同进化过程中空缺出的生存空间,从而使整个森林生态系统的结构和功能更加稳定和完善.
1 研究区概况
伏牛山自然保护区位于河南省西部(表1),东西走向,是我国北亚热带和暖温带的气候分区线,也是华北、华中、西南植物的镶嵌地带,属暖温带落叶阔叶林向北亚热带常绿落叶混交林的过渡区[7].特殊的地理位置和复杂多样的生态环境条件,加之人为干扰较小,使保护区保存了丰富的生物多样性资源.本区森林生态系统保存完整,优势种显著,林下的草本植物发育良好,且随海拔梯度变化明显.

表1 研究区概况Table 1 The conditions of experimental sites
2 研究方法
2.1样方设置
在南坡的宝天曼和北坡的白云山,从海拔600~2 000 m的山体上选择具有代表性的地段(主要是山脊及两侧优势种明显的地段),每隔海拔100 m确定至少1个20 m×20 m的样地,共设置样地66个[8],在样地的中心以及4个角分别设1个2 m×2 m灌木层样方和1个1 m×1 m草本层样方.其中包括草本层样方1 m×1 m 330个(图1).调查草本层的盖度、种类、个体数量、高度、冠幅、基径(丛径),以种群密度作为数量性状进行种群分布格局的测定.
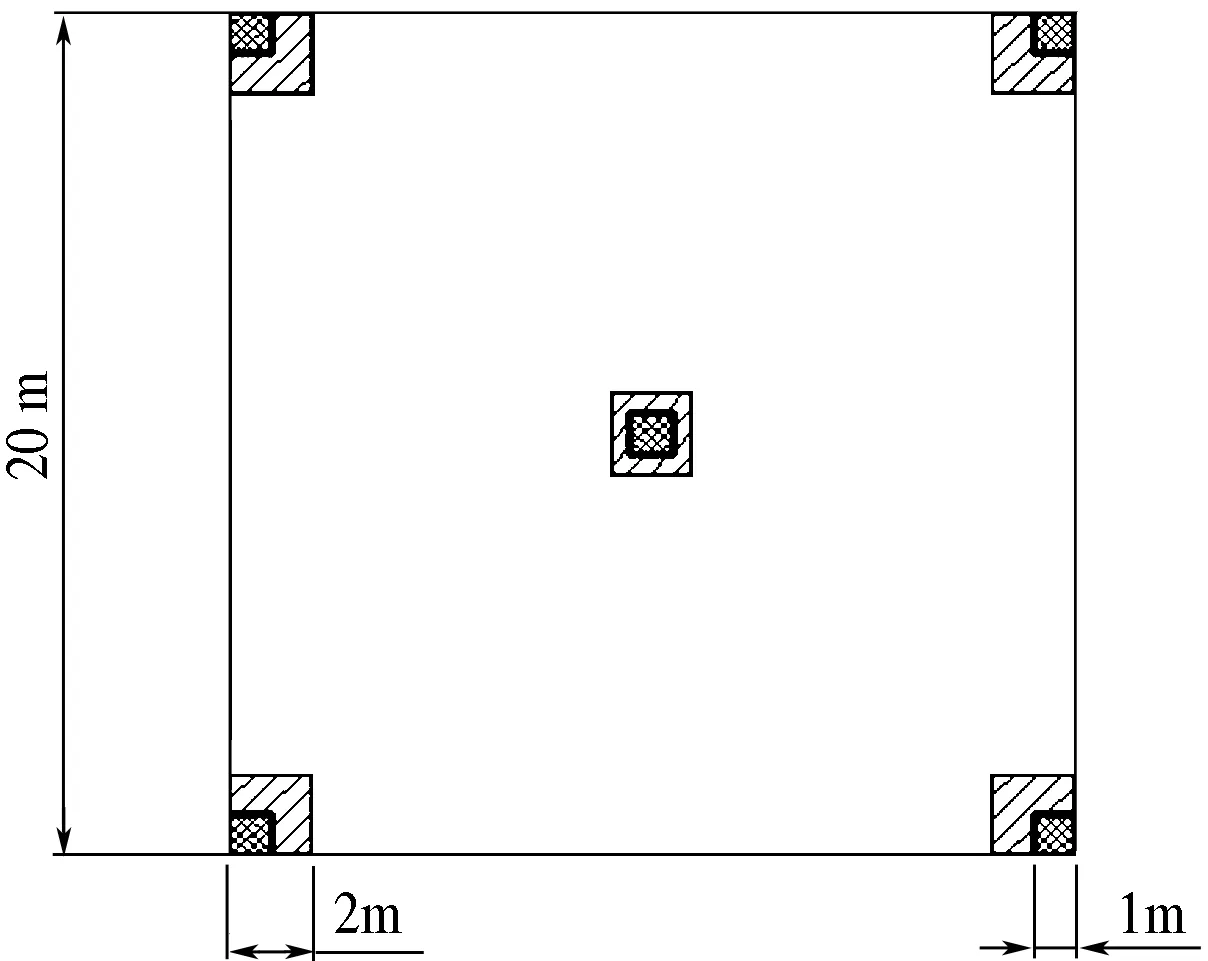
图1 样方设置图Fig.1 Quadrat settings
2.2计算方法
用以下方法计算优势种群的分布格局:
2.2.1 扩散系数[9]
DI=s2/x
式中:s2是种群数量性状的方差;x是种群数量性状的均值.当DI<1时,为均匀分布;DI=1时,为随机分布;DI>1时,为聚集分布.用DI值可对种群的分布格局做出初步判断.
2.2.2 聚集指数[10]
CI=(S2/X)-1
当CI<0时,为均匀分布;CI=0时,为随机分布;CI>0时,为聚集分布.
2.2.3 平均拥挤度[11]
m*=x+(s2/x-1)
m*表示生物个体在一个样方中的平均邻居数,它反映了样方内生物个体的拥挤程度,数值越大聚集强度越大,表示个体受其他个体的拥挤效应越大.
2.2.4 聚块性指数[9]
PAI=m*/x
PAI=1,分布格局为随机分布;PAI>1,为聚集分布;PAI<1,为均匀分布.
2.2.5 Green指数[9]
GI=(s2/x-1)/(n-1)
GI<0,为均匀分布;GI=0,为随机分布;GI>0,为聚集分布.
3 结果与分析
3.1研究区优势种分布
通过分析伏牛山自然保护区南北坡森林生态系统群落调查数据,根据出现次数并结合盖度,选出草本层主要优势种(表2).森林生态系统林下草本植物大部分个体体幅相对较小,重要值在统计上有一定的现实困难,用出现次数并结合盖度可以较好地反映其在某一环境的分布状况,由此确定研究区的优势物种.

表2 研究区草本层优势种Table 2 Dominant species of herbage layer at the experimental sites
3.2草本层优势种空间分布格局分析
在伏牛山自然保护区,草本植物分布格局与乔木层和灌木层的生长情况密切相关,特别是乔木层的演替情况对草本植物的分布影响较大,这主要与森林生态系统不同发育期林下光线的强弱有关.在低海拔区域,乔木层树龄较低,密度较大,林下光线较弱,草本植物的种类和数量较少,随着海拔的升高,乔木树种逐渐发育成熟,林下光线明显改善,草本植物的种类和数量明显升高.格局分析指标表明,优势度较大的草本植物都呈聚集分布(表3,表4).
在低海拔区域(1 000 m以下)(表3),大片聚集分布着栓皮栎与短柄枹的中幼龄林[8],林下光线较弱.草本植物以羊胡子草为优势种,且呈聚集分布,平均拥挤度较高.野菊花、莎草、掐不齐、臭草、荩草、蒿等植物经常相伴在中低海拔的林缘出现,反映出它们喜光等生物学特征[12],且它们也呈聚集分布,但是平均拥挤度远远低于羊胡子草.野菊花的扩散系数要高于其他草本物种,说明其在低海拔分布较为广泛,而它的聚集指数相对也较高,平均拥挤度仅次于羊胡子草.这些呈聚集分布的种群,说明它们的个体与个体之间存在吸引或衍生关系.费菜呈随机分布,多在潮湿的石旁出现,生存条件特殊,说明费菜种与种之间有着相互独立性,而且它们之间没有相互制约.苍术和兔儿伞呈均匀分布,说明种群的个体与个体之间存在着某种排斥关系,或者是个体间被某种力量或化学信息约束.

表3 低海拔(1 000 m以下)区域草本优势种空间分布格局Table 3 Herbage dominant species spatial distribution patterns at low-altitude (<1 000 m)

表4 高海拔(1 500 m以上)区域草本优势种空间分布格局Tab.4 Herbage dominant species spatial distribution patterns at high-altitude (>1 500 m)
在中高海拔区域(1 500 m以上)(表4),随着乔木发育逐渐成熟,林下的光线增加,加之人为干扰较小,草本植物的多样性比较丰富,特别是在山顶地带,草本植物的盖度达到最高.羊胡子草、芒、宽叶苔草等优势度较大的几种植物仍呈聚集分布.特别是鬼灯擎、蜂窝草等叶片较大的植物往往是成片出现,主要分布在中高海拔的成熟林区中,环境湿润,光线充足.而蕨类植物由于其繁殖方式的不同,也经常相伴出现,分布在潮湿的林下或石壁旁,是森林生态系统的较原始的物种.橐吾、酸模叶蓼和珍珠菜也呈聚集分布,主要分布在山谷溪流旁,喜阴湿的环境,叶质较厚.掐不齐、蒿在高海拔呈随机分布,它们在高海拔地段分布较少,零星地散布在林隙处,生存土壤较为贫瘠,可以理解为“先锋物种”.兔儿伞、唐松草呈均匀分布,它们主要分布在中海拔疏林内.
在伏牛山自然保护区,草本种群多呈聚集分布,只有少量的植物呈随机分布或均匀分布.呈聚集分布的种群,主要是由其生长、繁殖方式及环境资源和空间资源等决定的.如羊胡子草、芒、鬼灯擎等,它们根系分蘖萌生的繁殖方式可能是其表现为聚集分布的重要原因.随着环境的变化,由于不同植被适应环境范围的不同,它们的分布格局类型也会发生变化,如掐不齐和蒿在低海拔呈聚集分布,在高海拔呈随机分布,说明种群分布格局的地域性和尺度的相对性.
从低海拔区域到高海拔区域,乔木种群由幼龄林到成熟林,物种多样性增加,群落稳定性由弱变强.林下的草本种群的空间分布格局也有明显的变化(图2),群落的结构和功能也大不相同.演替初
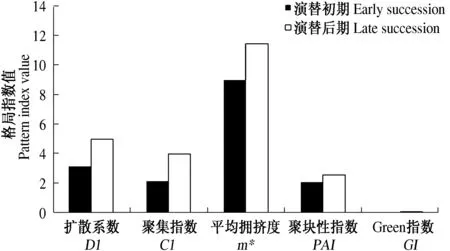
图2 不同演替期草本优势种空间分布格局比较Fig.2 Spatial pattern comparison between different succession of herbs dominant species
期,群落中乔木物种密度较大,林内光亮不足,下层草本层不发达,保持水土、养分能力不强.演替后期,由于种群中乔木密度降低,树龄较高,林下光线充足,草本层十分发达[12],保持水土、养分能力增强,整体上种群的扩散系数、聚集指数、平均拥挤度、聚块性指数和Green指数都较演替初期有显著的提高.如羊胡子草的平均拥挤度由24.19升高至43.45,扩散系数由6.33升至22.22;芒的平均拥挤度由9.82升高至28.14,扩散系数也由4.46升至14.68.当然也有个别种群由于环境变化,空间分布能力减小.如野菊花平均拥挤度由13.67升至16.61,扩散系数由7.27降低至3.26,说明其分布范围缩小,只能分布在较为适应的环境条件下.而费菜在低海拔呈随机分布,在高海拔呈聚集分布,可能和它的生物学特性有关,因为费菜喜光,经常在林窗出现,高海拔地段的成熟林林下可以提供更为充足的阳光.
4 讨论
4.1影响伏牛山森林生态系统草本种群分布格局的原因
种群分布格局一方面决定于物种的生物学特性和物种竞争等生态学过程,另一方面与外界环境密切相关.在伏牛山自然保护区,影响林下草本种群分布的限制性因子主要是光照和温度.对植物本身来说,由于草本植物个体矮小,繁殖和扩散能力有限,特别在森林生态系统中,林下草本物种以无性繁殖为主,如羊胡子草主要就是以分蘖方式进行繁殖,这就使种群更趋向于在小范围内的聚集分布.
在伏牛山自然保护区,森林覆盖率较高,草本物种生存所需要的光照主要是散射光或林窗、枝叶缝透射光,光照就成为影响草本植物分布的主要生态因子,而山地的地形地貌多变,这就加剧了这一生态因子的空间异质性.同时,山区小气候随海拔、坡向、坡度等间接因素呈现较明显的空间异质性.
所以,森林生态系统中的草本物种的繁殖方式等生物学特性,加上控制森林生态系统结构和功能的乔木树种的多样性分布,以及其他生态因子的空间异质性,使草本种群的空间分布多呈聚集分布.草本植物的聚集分布,在一定程度上填补了上层植物的空白,促进群落盖度增加,为分解者及微生物创造生存条件,提高森林生态系统保持水分、养分能力,加速植被恢复过程,对森林生态系统起到相当重要的稳定作用.
4.2不同演替期草本种群分布格局强度变化
从低海拔的演替初期群落到高海拔的演替末期群落,草本主要优势种的聚集分布格局类型基本上没有发生变化,但是格局强度发生规律性变化.以羊胡子草为例,它在演替的各个阶段的聚集分布格局并未发生变化,但是其扩散系数由6.33变为22.22,聚集指数由5.33变为21.22,平均拥挤度由24.19变为43.45,聚块性指数由1.38变为2.17,Green指数由0.10变为0.38.扩散系数的增加,说明羊胡子草种群进一步扩展,完成对空间的侵占,平均拥挤度也在增加,主要原因是因为演替后期,群落环境稳定,土壤肥力、光照度增加.而生物多样性的增加,使得群落的结构更加复杂,空间异质性增强,加上不同物种的资源利用方式的不同,几个物种在同一群落同时形成聚集分布是完全有可能的[12],各种群的聚集指数和聚块指数进一步提高,呈现高度的镶嵌性分布,不同的物种都能在各自的空间占有一席之地,且得到极大发展,使得整个森林生态系统更加稳定.
在森林生态系统中,草本植物可以理解为“基础种”,它们在森林形成初期为“先锋种”,在森林稳定期为“保持种”,在退化的森林生态系统中为“保留种”[13],草本植物这种灵活的生存机制能使其适应不同的生态条件.研究群落空间格局及其生态学过程是生态学的中心任务之一[14],不仅可以了解种群的空间分布特点,更重要的是可以根据格局的变化与关联性推演种群的动态过程及潜在的维持机理,从而推断自然群落中物种共存的维持机制[15,16],而生存机制相对灵活的草本植物更适合此类研究的开展.
[1] 张大勇, 姜新华,雷光春,等.理论生态学研究[M].北京:高等教育出版社,2000:170-172.
[2] 胡 楠, 范玉龙,丁圣彦. 伏牛山自然保护区不同演替期乔木优势种空间分布格局比较[J]. 河南大学学报:自然科学版,2010,40(5):493-497.
[3] 范 娟,赵秀海,汪金松,等. 江西九连山亚热带常绿阔叶林优势种空间分布格局[J]. 生态学报,2012,32(9):2729-2737.
[4] 陈志伟,杨京平,王荣洲,等. 浙江省加拿大一枝黄花(Solidagocanadensis)的空间分布格局及其与人类活动的关系[J]. 生态学报,2009,29(1):120-129.
[5] 杜丽侠,邢韶华. 北京八达岭地区灌木群落空间分布格局与土壤环境因子的关系分析[J]. 西北植物学报,2009,29(3):601-607.
[6] 蒋雪琴,刘艳红,赵本元. 湖北神农架地区巴山冷杉(Abiesfargesii)种群结构特征与空间分布格局[J]. 生态学报,2009,29(5):2211-2218.
[7] 尚富德,王磐基,冯广平,等. 伏牛山南北自然过渡地带植物多样性的特征及其成因分析[J].河南大学学报:自然科学版,1998,28(1): 54-60.
[8] 胡 楠, 范玉龙, 丁圣彦,等. 伏牛山自然保护区森林生态系统乔木植物功能型分类[J].植物生态学报, 2008,32(5):1104-1115.
[9] 丁岩钦. 昆虫数学生态学[M].北京:科学出版社, 1994:25-27.
[10] 郑元润. 沙地云杉种群分析中各种方法的有效性[J].植物生态学报, 1997,21:480-484.
[11] LLOYD M. Mean crowding[J]. Animal Ecology, 1967,36:1-30.
[12] 张大勇,姜新华,雷光春,等. 理论生态学研究[M].北京:高等教育出版社,2000:169-172.
[13] 范玉龙,胡 楠,丁圣彦,等. 伏牛山自然保护区森林生态系统草本植物功能群的分类[J].生态学报,2008,28(7):3092-3101.
[14] 幸晓平,徐 斌,王秀山,等. 碱化草地群落恢复演替空间格局动态分析[J].生态学报, 2001,21(6):877-882.
[15] CONDIT R,HUBBELL S P,FOSTER R B.Density dependence in two understory tree species in a neotropical forest[J]. Ecology,1994,75(3):671-680.
[16] HE F L,DUNCAN R P. Density-dependent effects on tree survival in an old-growth Douglas fir forest[J]. Journal of Ecology,2000,88(4):676-688.
(责任编辑:朱秀英)
SpatialpatternofherbsdominantspeciesatFuniuMountainNationalNaturalReserve
FAN Yu-long1,2, HU Nan1,2, DING Sheng-yan1, LU Xun-ling1
(1.The College of Environment and Planning, Henan University, Kaifeng 475001, China;2.Nanyang Institute of Technology, Nanyang 473004, China)
Through the vegetation surveys of different altitudes herbs, we study the herbaceous dominant species’ spatial distribution pattern and its dynamics. The results showed that: the dominant species are mostly aggregated distribution, such asCarexlanceolata,Miscanthussinensis,PteridiumaquilinumandDendranthemaindicum;Sedumkamtschaticumis randomly distributed,AtractylodeslanceaandSyneilesisaconitifoliaare evenly distributed. From the early to late-successional succession, aggregated distribution of multi-species in the same community is increasing. The value of spatial distribution pattern is increasing, and only a few species’ spatial pattern types are changed, withKummerowiastriataandArtemisiaargyichanged from aggregated distribution to even distribution. The sensitivity of herbaceous plants to environmental changes is strong, making them indicating the environmental changes.
Funiu Mountain Nature Reserve; herbage dominant species; spatial pattern; succession; forest ecosystem
Q 948
:A
2014-03-04
国家自然科学基金资助项目(41071118,40671175);河南省科技厅重点科技攻关项目(112102310416);河南省教育厅项目(2010C180006)
范玉龙,1978年生,男,河南南阳人,博士研究生,主要从事植被生态学研究.
丁圣彦,1963年生,男,河南开封人,教授,博士生导师.
1000-2340(2014)05-0590-06
猜你喜欢
散文诗(2021年22期)2022-01-12
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
文物鉴定与鉴赏(2019年14期)2019-09-24
启迪与智慧·下旬刊(2019年1期)2019-09-10
环球时报(2019-05-23)2019-05-23
现代园艺(2017年13期)2018-01-19
现代园艺(2017年13期)2018-01-19
小说月刊(2014年1期)2014-04-23
郑州大学学报(工学版)(2014年6期)2014-03-01
