
基于遥感数据分析南海叶绿素与颗粒物的季节变化与相互关系
2015-01-05 05:12王晓琦邢小罡王金平李娜
海洋学报 2015年10期
王晓琦,邢小罡*,王金平,李娜
(1. 中国海洋大学 海洋环境学院,山东 青岛 266100)
基于遥感数据分析南海叶绿素与颗粒物的季节变化与相互关系
王晓琦1,邢小罡1*,王金平1,李娜1
(1. 中国海洋大学 海洋环境学院,山东 青岛 266100)
本文基于卫星遥感的叶绿素a浓度与颗粒物后向散射系数月平均数据以及其他海洋与气象参数,详细分析了两个生物光学参量在季节尺度上的相关性及其与物理参数的耦合关系,并运用光驯化模型分析了南海表层水体浮游植物的生理学季节变化特征。结果表明,受南海地形和风场等物理参量的变化,南海叶绿素a浓度与颗粒物后向散射系数存在显著的季节和空间分布特征,具有一定的共变性和差异性。在南海近岸及浅水区,叶绿素a浓度与颗粒物后向散射系数有很好的耦合关系;而在南海深水海盆区,叶绿素a浓度冬高夏低,其季节循环过程与颗粒物后向散射系数相反,这主要是受浮游植物生理学过程的影响。“生物量控制区”与“光驯化控制区”的分界在南海与陆架-海盆分界线一致,体现了水深条件对浮游植物生理状态的影响。此外本文还发现,在吕宋海峡西部海区,叶绿素a与颗粒物后向散射系数的关系表现出“生物量-光驯化共同控制”的特点。
叶绿素a浓度;颗粒物后向散射系数;南海;季节尺度;光驯化
1 引言
海洋覆盖了地球表面的70%,浮游植物是海洋生态系统中主要的初级生产力,贡献了整个地球净初级生产力的一半[1—2]。浮游植物的光合作用是海洋生物泵的主要过程之一,驱动了海洋的生物地球化学循环,也影响着整个地球系统的气候变化。过去20年间,随着海洋遥感技术的发展,水色卫星传感器可以提供全球海洋表层生态系统的遥感数据,解决了传统的船基走航观测时空分辨率不足的问题,大大提升了海洋生态学与生物地球化学——特别是对浮游植物——在大时空尺度上的观测能力[3]。尤其随着各种半分析算法(Semi-analytical algorithms)的发展[4—7],多个独立的水色参数能够同时反演(如叶绿素a浓度、非藻类吸收系数、颗粒物后向散射系数),人们不仅可以更全面地了解从区域到全球尺度的水体类型和生物光学特征,而且通过在不同时空尺度下水色参数间相互关系的研究,还可以更深入地理解海洋表层浮游植物的动态变化过程及其与物理过程的耦合机制[8]。
作为西太平洋最大的边缘海,南海的物理、生态与生物地球化学参数的时空分布及其驱动机制一直是区域海洋学研究的重点。基于卫星遥感数据,前人已多次研究了南海叶绿素a浓度的时空变化及其同物理过程的耦合关系[4,9—15],然而,对于其他生物光学参数,特别是颗粒物后向散射系数的时空分布以及叶绿素与颗粒物后向散射系数间相互关系的研究却非常缺乏。单一参数的分析已无法满足南海生态学研究的需要,尤其对于浮游植物的研究,虽然叶绿素a目前仍然是浮游植物生物量最常用的指标性参数[16—18],但有研究早已指出浮游植物的生理学过程也会影响叶绿素a的浓度[19—20]。浮游植物的生长环境因素(包括温度、营养盐、光照)不仅影响藻类光合作用效率和生长率[20],还调控其生理状态,特别是浮游植物会跟随环境光照和营养盐水平调整其细胞内的叶绿素a含量[19,21—22]。在生长光照不足且营养盐充足的情况下,浮游植物自身会增加其细胞内的叶绿素a以便更有效地吸收光能;而当营养盐不足的情况下,浮游植物则会减少叶绿素a含量。生理学调整甚至可以浮游植物叶绿素a与碳含量比率使产生超过10倍的(CChl a/Cphy)变化[3]。在自然水体中,光照的季节性循环通常是影响生理学调整的主要因素,这一浮游植物依赖于光照的变化过程称为“光驯化(photoacclimation)”或“光适应(photoadaptation)”[21—22]。Siegel等[8]和Behrenfeld等[23]发现,颗粒物后向散射系数与叶绿素在富营养海区(高纬度和上升流海区)一般存在较好的耦合关系,说明浮游植物生物量的改变主导了叶绿素的变化,在这些海区叶绿素是很好的生物量评价指标;然而在一些贫营养海区(大部分的热带和亚热带海区)则出现两者相关性的减弱,说明在这些海区生理学过程对叶绿素a的变化影响很大,如果仍然将叶绿素a作为生物量的指标进行分析就会存在很大的局限性,甚至可能导致错误地理解浮游植物的季节变化特征。而作为颗粒物丰度的一个重要指标,颗粒物后向散射系数虽然在河口区和近岸浅水区易受到无机悬浮泥沙的影响(河流直接输入、底层泥沙的再悬浮),但在深水大洋海区则显示出与浮游植物生物量和碳强烈的正相关关系,在这些海区,颗粒物后向散射系数通常可以作为更好的浮游植物生物量的指标[8,23]。
南海纵跨热带和亚热带,其深水海盆是典型的贫营养海区[24],然而生理学变化过程却一直在南海浮游植物的研究中很少被提及,叶绿素a一直作为浮游植物生物量的唯一指标进行分析。在此背景下,本文基于水色卫星遥感数据,在季节尺度上详细分析南海叶绿素与颗粒物后向散射系数的时空变化与相互关系,以期初步揭示南海浮游植物的季节变化过程、光驯化特征以及生态与物理耦合过程的区域性差异。
2 材料和方法
叶绿素a浓度(CChla,单位:mg/m3)、440 nm颗粒物后向散射系数(bbp,单位:m-1)、海表面日有效光合辐照度[Photosynthetically Active Radiation,PAR,单位:mol/(m2·d)]、海表面温度(Sea Surface Temperature,SST,单位:℃)均来自美国国家航空航天局(National Aeronautics and Space Administration,NASA)提供的Aqua卫星上搭载的中分辨率成像光谱仪(MODerate resolution Imaging Spectroradiometer,MODIS)遥感观测的月平均数据产品,空间分辨率(1/12)°×(1/12)°(约9 km×9 km)(http://oceancolor.gsfc.nasa.gov/cgi/l3)。其中,CChla与bbp产品通过GSM(Garver-Siegel-Maritorena)半分析算法反演得到[4—5];PAR数据基于Frouin和Chertock提出的辐射传输方程[25]和大气数据估算获得;SST产品来自于MODIS的11 μm热红外通道遥感观测。本文选取2002年7月到2014年6月的12年月平均数据,为去除年际变化的影响,将多年月平均数据进行逐月平均得到气候态月平均数据。
风场资料为美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration,NOAA)下属的国家气候数据中心(National Climatic Data Center,NCDC)提供的融合海表面风矢量气候态月平均数据(http://www.ncdc.noaa.gov/oa/rsad/air-sea/seawinds.html),空间分辨率(1/4)°×(1/4)°。该数据的风速融合了6台卫星(散射计QuikSCAT,微波辐射计AMSR-E、TMI,以及DMSP系列卫星F13-SSM/I、F14-SSM/I、F15-SSM/I)的遥感观测结果,风向来源于NCEP再分析数据(NRA-2)以及ECMWF(欧洲中期天气预报中心)模型的模拟结果。
混合层深度(Mixed Layer Depth,MLD)资料来源于SODA(Simple Ocean Data Assimilation)再分析得到的月平均数据(http://orca.science.oregonstate.edu/1080.by.2160.monthly.hdf.mld.soda.php#monthly)。空间分辨率为(1/6)°×(1/6)°,通过1997年7月到2004年6月共7年的月数据,进行逐月平均得到MLD的气候态月平均数据。
为了进一步评估南海表层浮游植物的生理学变化特征,本文基于Behrenfeld等提出的方法[23],通过气候态月平均的叶绿素a浓度、日有效光合辐照度以及混合层深度,计算了混合层内的浮游植物生长光照(growth irradiance,Ig):
Ig=PAR exp(Kd490×MLD/2),
(1)
式中,Kd490称为490 nm的下行漫衰减系数,由叶绿素a浓度通过经验关系估算得到[26]:
Kd490=0.016 6+0.082 5 CChl a0.652 9.
(2)
通过bbp估算浮游植物碳含量(Cphy,单位mg/m3):
Cphy=13 000×(bbp-0.000 35).
(3)
通过PhotoAcc模型[23,27]建立CChla/Cphy与生长光照Ig之间在季节尺度上的经验回归关系:
CChla/Cphy=A+B×exp(-3Ig),
(4)
式中,A和B为回归系数。
3 结果与讨论
3.1 南海物理与生物参量的季节变化与空间分布特征
图1显示了南海气候态月平均数据中1、4、7、10月(分别代表冬、春、夏、秋四季特征)的海表面风场、温度场以及CChla与bbp的分布特征。从图1a~d可以看出,南海地处东亚季风区,冬半年南海地区为东北季风,在吕宋海峡西北部和湄公河口东部达到最高值;而夏半年南海主要为西南季风,其盛行期在7月,17°N以南海域风力较大,多为西南风;20°N以北海域风力较弱,多为偏南风,风速高值区出现在湄公河口东部。春季和秋季处于季风转换时期,4月风速达到全年最小值。南海SST的季节变化主要受呈现纬度差异(见图1e~h),并在一定程度上受到风场和流场的影响。冬季全海区降温到最低值,受冬季强西边界流输送北部冷沿岸水的影响[28],在越南东部外海形成一条明显的冷舌。春季随着东北季风的松弛,南海开始增暖,越南东部外海的冷舌开始消失。夏季南海SST达到最高值,且整个海区的空间差异性最小,但在湄公河口东部(10°~15°N)存在一个低值区,与此时强劲的西南季风产生局地上升流有关[29]。
受南海的地形特征(陆架与深水海盆的地理分布)以及风场、SST等物理参量的复杂时空变化,南海海域叶绿素a和颗粒物后向散射系数也呈现出显著的季节和空间变化。总体来说,南海叶绿素a浓度与bbp的空间分布基本一致,存在显著的共变特征(见图1i~p)。两者都具有近岸海域高、海盆中央区低的特点,以及西部近岸海区(越南东部外海)明显高于东部近岸海区(菲律宾西部外海)。海盆中央区域整体变化不大,但值得注意的是,在吕宋海峡的西北部(17°~20°N)存在一个冬季的高值区,之前人们通过卫星观测的叶绿素a浓度数据已多次讨论过该区域的冬季藻华现象[9,11,30—31],也已得到现场观测的确认[32—33],一般认为与冬季强烈的东北季风产生的局地上升流有关。吴日升和李立[34]指出,形成该上升流的原因不仅与风场有关,还包括海盆尺度环流的强迫作用。本文发现,该现象不仅出现在叶绿素a浓度数据中,bbp同样也有一定程度地升高,但叶绿素a浓度增加的信号更为明显,范围也更大,从吕宋海峡一直延伸至珠江外海。从时间变化上来看,在中央海盆区叶绿素a浓度基本存在冬高夏低的季节特征,但bbp变化相比叶绿素a浓度不太明显。在河口区两者都存在冬夏两季高、春季低的特征。如在珠江口-粤东海区以及湄公河口-越南东部外海,叶绿素a浓度与bbp的高值区在夏季面积最大,这与河流的径流量有密切的联系(具体讨论见3.2节),也与局地的夏季上升流有关[34]。
为了进一步研究物理与生态参数以及生态参数之间在空间分布与季节变化上的相似性与差异性,本文计算了气候态月平均叶绿素a浓度数据分别与海表面风速、SST、bbp的相关性以及bbp与SST的相关性在南海的空间分布(见图2)。从图2a可以看出,叶绿素a浓度与风速在南海整个海区基本存在正相关关系,尤其在吕宋海峡西部的正相关性最为显著,但在近岸部分海域有所减弱,甚至在东沙群岛海山环礁、珠江河口羽流区和台湾南部附近等,二者出现负相关。此外,叶绿素a浓度与SST在整个南海主要呈负相关关系,负相关最显著的区域也位于吕宋海峡西部(见图2b)。同样,叶绿素a浓度与SST在近岸海域的负相关性也明显降低,甚至在某些海域(如广东沿海、湄公河口、台湾岛南部沿岸等)存在正相关关系。叶绿素a浓度与风速以及SST的相关性在之前的许多研究中已有讨论,一般解释为:风速越大,表层混合越强(或促进局地上升流),因而导致海表温度降低、以及将下层的营养盐向上补充,从而促进了表层浮游植物的生长;此外,冬季海面冷却增密,产生对流混合,同样导致表层的营养盐水平以及浮游植物生物量的增加[9,30]。
图2c和2d分别显示了bbp与叶绿素a浓度以及bbp与SST在季节尺度上相关性的空间分布。虽然图1显示出叶绿素a浓度与bbp在空间上存在显著的相关性,且该相关性在时间尺度上的变化并不大(气候态月平均数据中两者逐月的空间分布相关性最低0.75,最高0.9,该结果未通过图表显示),但在时间尺度上相关性则存在显著的空间差异。如图2c所示,两者在近岸海域存在很强正相关性;而在中央海盆区,相关性则开始降低,甚至在南北两个区域(图中N和S区域)中出现负相关关系。虽然颗粒物后向散射系数bbp可能受到无机悬浮物、颗粒物粒径以及藻种类型的影响,但一阶近似的条件下仍然正相关于颗粒有机碳(Particulate Organic Carbon,POC)的含量,特别是在远离河口的深海大洋(如南海海盆区),无机悬浮物的影响非常小。此外,Behrenfeld等[23]提出可以通过卫星遥感的bbp近似地估计浮游植物的生物量,并建立了bbp与浮游植物碳含量的经验关系(公式(3)),基于这一方法,他们发现叶绿素a浓度与bbp在高纬度海区存在强耦合关系,但在一些低纬度海区耦合关系很弱,两者可能存在相位差,甚至出现反向变化(负相关)。Siegel等[8]进一步提出叶绿素a浓度与bbp显著正相关(强耦合)的海区为“生物量控制区”,而耦合关系弱的海区则称为“光驯化控制区”。按照上述定义,南海近岸海区可称为“生物量控制区”,该区域中颗粒物与叶绿素a浓度有良好的耦合性。虽然近岸海水中含有更多的无机物(颗粒物的含量并不能代表浮游植物的丰度),但由于水深较浅,表层无机物浓度的分布也与混合作用关系密切。混合作用不仅将营养盐也包括底层碎屑、泥沙输送至海表面,从而促进浮游植物生物量的增加以及叶绿素a浓度的增长、以及总颗粒物含量的增加,因而导致叶绿素a浓度和bbp存在强耦合关系。中央海盆则表现出“光驯化控制区”的主要特征:混合层相对较浅[35],导致表层营养盐常年匮乏[24],表层叶绿素a浓度与bbp的含量都很低,浮游植物在垂向分布上会出现叶绿素次表层极大值[36—37]以及生物量极大值[38],且两个极大值存在深度上的差异,类似于副热带环流区中心贫营养水体的生态学特征[39]。在此情况下,浮游植物的生理学调整过程可能是叶绿素a浓度季节变化的主要原因,类似于其他贫营养海区[8],而导致两者的耦合关系减弱,甚至出现负相关关系[23,40—41]。

图1 南海海表面风(SSW,a~d)、海表面温度(e~h)、叶绿素a浓度(i~l)以及颗粒物后向散射系数(m~p)的月平均气候态数据在1、4、7、10月的空间分布Fig.1 Spatial distributions of monthly climatology data of sea surface wind (a-d),sea surface temperature (e-h),chlorophyll a concentration (i-l) and particle backscattering coefficient (m-p) in the South China Sea in January,April,July and October
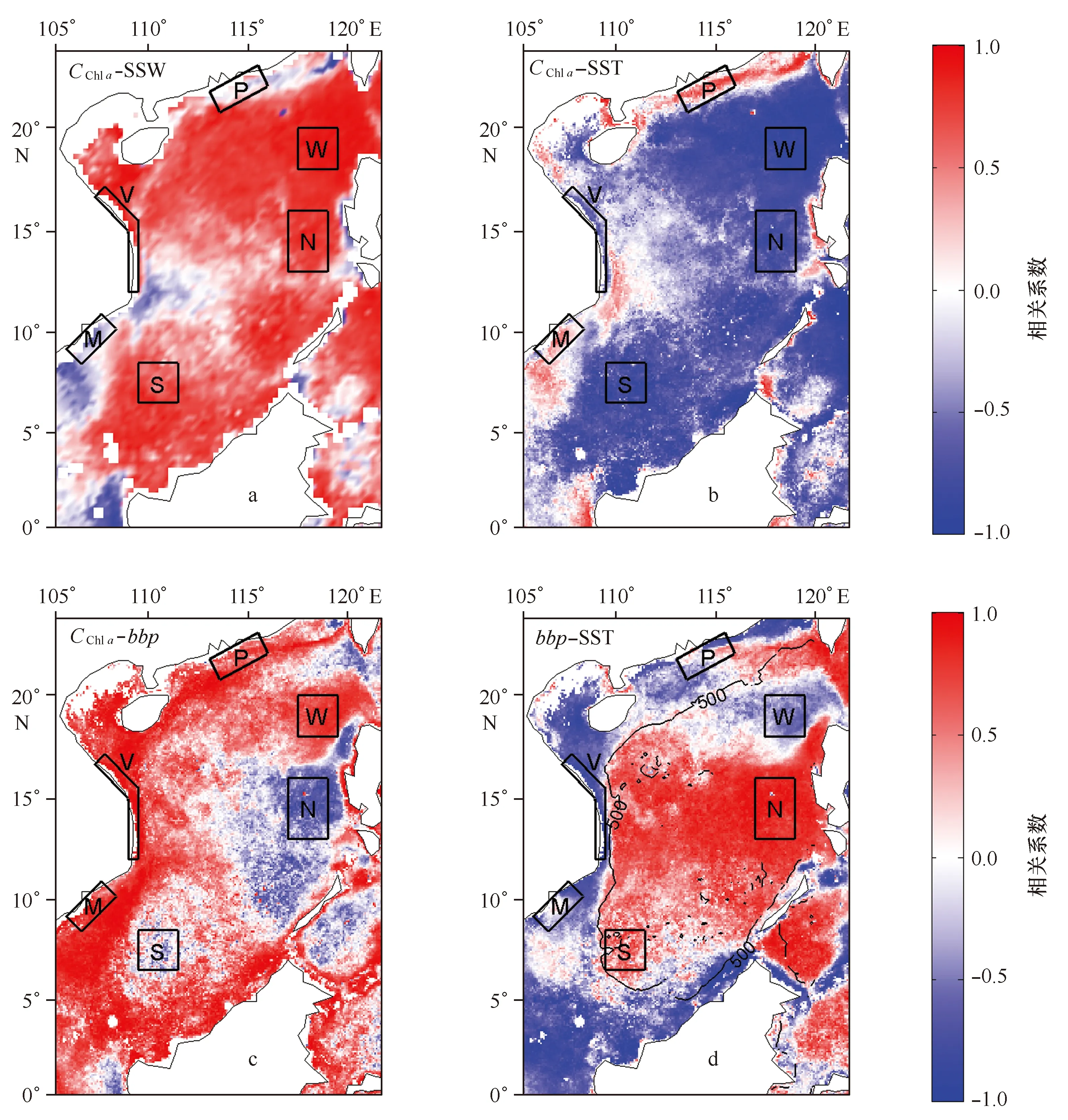
图2 南海叶绿素a浓度(CChl a)、颗粒物后向散射系数(bbp)、海表面风速(SSW)、海表面温度(SST)气候态月数据之间相关系数的空间分布Fig.2 Spatial distributions of correlation coefficients between chlorophyll a concentration (CChl a),particle backscattering coefficient (bbp),sea surface wind speed (SSW) and sea surface temperature (SST) of monthly climatology data in the South China Sea图c中黑色实线为500 m等深线,黑框表示了图3与表1中对应的6个典型区域,M:湄公河口,P:珠江口,V:越南东部陆架,W:吕宋海峡西部,S:南部深水海盆,N:北部深水海盆The black solid lines plotted in the panel c represent the 500 m depth contour; The black boxes represent the corresponding 6 typical regions shown in Fig. 3 and Table 1,M: Mekong River Plume; P: Pearl River Plume; V: Viet-namese Continental Shelf; W: West of Luzon Strait; S: Southern Deep Basin; N: Northern Deep Basin
此外,图2d显示出,bbp与SST的负相关关系仅出现在近岸海区,而在中央海盆两者则呈现正相关关系。特别是在越南东部海域,负相关到正相关的梯度变化非常明显。结合图中的500 m等深线,我们可以进一步看出梯度变化最大的区域基本处于陆架与海盆交界的陆坡区(除吕宋海峡西部外)。二者交界线北部沿广东沿岸海岸线平形分布至台湾岛西南部,西部交界线自北部湾向南至湄公河口三角洲以北,南部交界线西起昆仑群岛,东至纳土纳群岛。关于bbp与SST相关性空间差异的具体原因,本文将在下一节结合几个典型区域内物理与生物参数具体的季节变化详细讨论。
3.2 南海典型区域的物理与生物参量季节变化以及相互关系
基于气候态月平均叶绿素a浓度与bbp数据的空间分布特征(见图1)及其各参数时间相关性的空间分布特征(见图2),本文在整个南海范围内选取了6个典型区域(3个近岸区和3个外海区,见图2),详细讨论其物理与生态参数的季节变化过程,其中包括:两个重要的河口区,珠江口(Pearl River Plume,PRP)和湄公河口(Mekong River Plume,MRP);南海西部典型的近岸海区,越南东部大陆架(Vietnamese Continental Shelf,VCS);冬季藻华的典型海区,吕宋海峡西部(West of Luzon Strait,WLS);以及叶绿素a浓度与bbp呈负相关的两个海盆区域,南部深水海盆(Southern Deep Basin,SDB)和北部深水海盆(Northern Deep Basin,NDB)。典型区域中平均叶绿素a浓度、bbp、海表面风速(SSW)、SST以及混合层深度的季节变化如图3所示,参量间相关系数列于表1。
3.2.1 河口区
在湄公河口(MRP),叶绿素a浓度与bbp在季节尺度上具有较高的相关性(r=0.59)。从图3中也可以看出,二者在季节变化上具有明显的同步性:夏季(8月)最高,冬季(12月)也保持高值,春季(4月)最低。赵辉等[9]基于高度计观测资料发现,湄公河口夏季为上升流,冬季为下降流,说明夏季西南季风加强了上升流,带来丰富的营养盐,同时丰水期河流径流增大,使得叶绿素a浓度和bbp达到高值。而冬季东北季风在该区域达到高值,强风使得沿岸浅海区引起对流混合,削弱了下降流的影响,因此在冬季叶绿素a浓度和bbp也达到高值。
珠江口(PRP)同MRP均为近岸河口区域,物理与生态参数的季节特征均非常相似。夏季西南季风会增强上升流的影响,冬季东北季风会削弱下降流的影响,导致PRP也在夏季和冬季有较高的叶绿素a浓度和bbp,并在春季达到最低。通过比较相关系数我们还可以发现,在两个河口区,bbp与风速的相关性都是最高的(r分别达到0.69和0.96),可以看出季风对河口区颗粒物的影响非常显著。同时,夏季汛期河流水量的增加,携带更多的营养盐,也促进了两个河口区浮游植物和颗粒物的夏季峰值。通过比较径流量数据我们发现,珠江最大流量出现在6、7月[42],而湄公河一般出现在8月[43]。与之相对应,在PRP和MRP区域,夏季bbp的最大值分别出现在6月和8月,与相应河流的径流量在春-夏季变化非常相似。此外值得注意的是,MRP和PRP的bbp与物理量的相关性基本都比叶绿素a浓度与物理量更显著(除MRP区域bbp与混合层深度的相关性较弱),说明由于河口区水体中含有较多的无机悬浮颗粒物,bbp中除浮游植物的贡献外,无机物也占有很大的比重。而无机物的含量主要与径流输运以及混合作用有关,因而bbp受物理过程的影响比叶绿素a浓度更大。而且,两个河口区的叶绿素a浓度和bbp虽然季节变化特征相似,但也存在区域差异性:MRP与PRP的叶绿素a浓度基本一致,但bbp则高出大约2倍。根据统计数据,珠江和湄公河的年输沙量相当(约8 000×104t/a),但珠江的水通量大约是湄公河的5倍(分别为3 083×108和599×108m3/a)[44],说明湄公河携带的泥沙(无机悬浮物)含量比珠江更高,因此造成了MRP比PRP具有更高bbp的现象。此外,这种差异也可能与MRP比PRP含有更多的微微型浮游植物有关[45—46]。
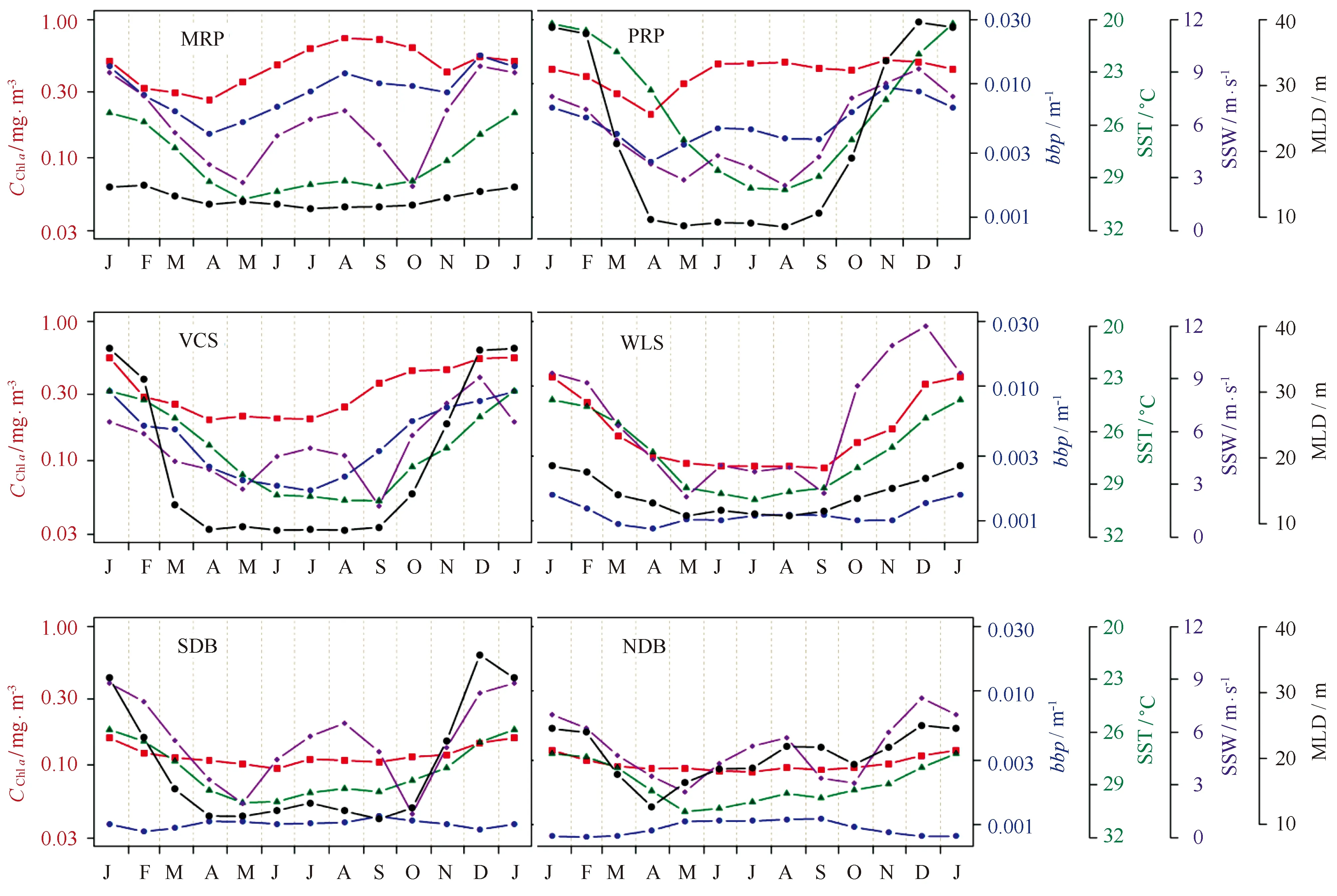
图3 南海6个典型海区的叶绿素a浓度(CChl a)、颗粒物后向散射系数(bbp)、海表面温度(SST)、海表面风速(SSW)、混合层深度(MLD)气候态月数据的季节变化Fig.3 Seasonal variations of chlorophyll a concentration (CChl a),particle backscattering coefficient (bbp),sea surface temperature (SST),sea surface wind speed (SSW) and mixed layer depth (MLD) of monthly climatology data in 6 typical regions of the South China Sea 湄公河口:MRP;珠江口:PRP;越南东部陆架:VCS;吕宋海峡西侧:WLS;南部深水海盆:SDB;北部深水海盆:NDB; 注意图中SST的坐标是倒转绘制的,为了显示其与SSW、MLD的负相关关系Mekong River Plume,MRP; Pearl River Plume,PRP; Vietnamese Continental Shelf,VCS; West of Luzon Strait,WLS; Southern Deep Basin,SDB; Northern Deep Basin,NDB; note that the SST was plotted reversely,for showing its negative relationship with SSW and MLD
虽然在河口区,无机颗粒物是影响后向散射系数的重要因素,但由于bbp与叶绿素a浓度在MRP与PRP的相关性仍然达到0.59和0.40,说明bbp与浮游植物的生物量在统计学意义上仍然存在一定程度的耦合关系。如果无机颗粒物的影响降低了bbp与浮游植物生物量的相关性,则真实的浮游植物生物量与叶绿素a浓度的相关性应该更高。
3.2.2 越东陆架与吕宋海峡西部
在越南东部陆架区(VCS),叶绿素a浓度与bbp含量比两个河口区要低,但高于其他3个外海区,且两者的耦合关系是所有典型区域中最高的(r=0.93)。生态参量的季节变化与两个河口区差别很大:叶绿素a浓度在4—7月维持低值,8月开始突然增加,到冬季(12—1月)达到最高值;bbp则在7月达到全年最低,1月最高。由于VCS没有大河流输入,两者仅与局地的混合过程有关,因而耦合关系更强。此外,VCS的生态参量与物理参量(特别是与SST和混合层深度)的耦合关系显著高于两个河口区,其中与混合层深度的相关性最高,r分别达到0.82和0.92,也说明在没有河流影响的浅水陆架区,生态参量主要与混合过程有关。
在吕宋海峡西部(WLS),叶绿素a浓度与bbp的典型季节特征是:夏季与其他海盆区(NDB和SDB)相似;冬季发生藻华,数值上接近近岸海区(VCS、PRP、MRP)。关于冬季藻华的成因,一方面,由于中国大陆、台湾岛和吕宋岛形成的“狭管效应”,使得南海北部风速加大,并在WLS形成一个风速的极大值中心[30]。强烈的东北季风增加了Ekman抽吸,从而促进蒸发冷却作用,使得SST降低,进而驱动上层海洋的对流混合,混合层深度增大。通过这种方式,营养盐被卷挟到表层,从而促进了浮游植物的生长。此外,赵辉等[9]、Tang等[11]以及Wang等[31]也指出在冬季WLS的局地上升流也会促进浮游植物的生长。虽然,叶绿素a浓度与bbp在WLS也存在很强的耦合关系,但两者的季节变化过程仍然不同:叶绿素a浓度冬季(1月)最高并在春末到秋初的很长时间内(5-9月)维持最低值;bbp最高值也出现在1月,但最低在春秋两季(4月和11月),夏季则有一定程度的增加(夏季与叶绿素a浓度的变化过程相反)。
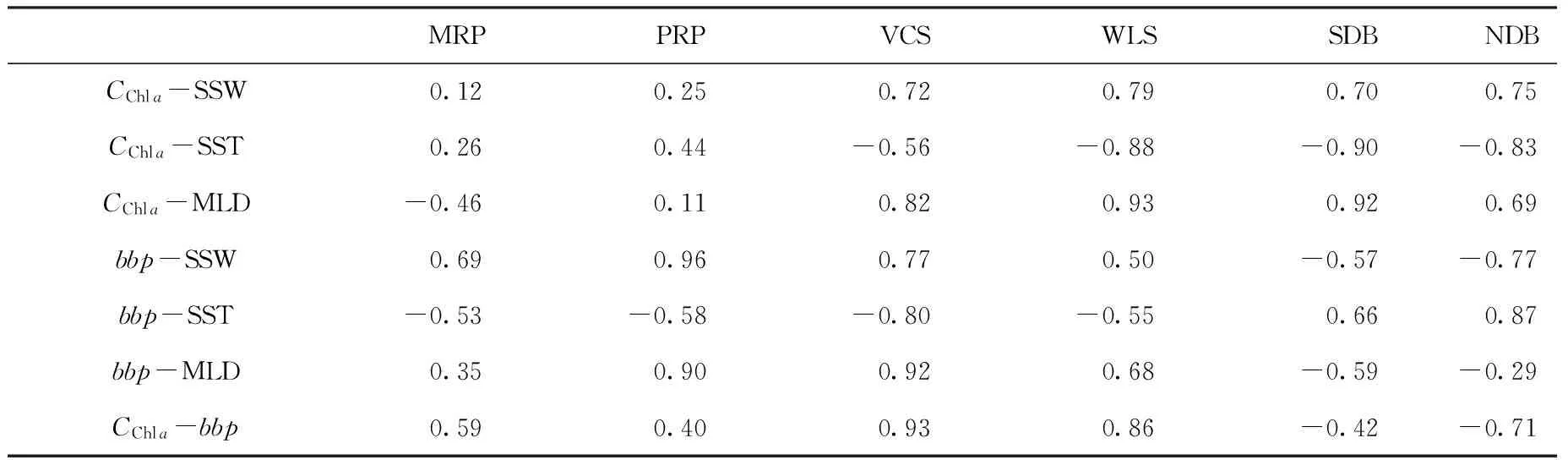
表1 南海6个典型区域生物光学与物理参量气候态月平均数据的相关系数Tab. 1 The correlation coefficients of climatological monthly bio-optical and physical parameters in six typical regions of South China Sea
注:区域位置见图2。
3.2.3 深水海盆区
在北部深水海盆(NDB)和南部深水海盆(SDB)两个区域中,物理与生态参数的季节变化有一定的相似性:叶绿素a浓度与bbp均代表了南海的最低水平,且全年变化的幅度都很小;叶绿素a浓度基本维持在0.1 m/m3左右,仅在冬季稍高;bbp则在夏季稍高,数值基本在0.001 m-1附近;两者因而在季节尺度上具有负相关关系(在NDB更明显,r达到-0.71)。虽然Behrenfeld等[23]基于卫星数据以及邢小罡等[39]、Mignot等[41]基于现场数据都发现在其他贫营养海区也存在叶绿素a浓度与颗粒物之间负相关性的现象,与MLD相较于营养盐跃层过浅有关。
(1)首先,由于南海深水海盆区的表层水体全年处于营养盐限制(主要是氮限制)的环境下[24],过浅的混合层难以对表层的浮游植物增长提供必要的营养盐支持。Qu等[35]基于现场观测的大量温盐数据,指出南海的混合层深度浅于吕宋海峡外的西太平洋大约10~20 m。通过全球MLD的气候态月数据(见De Boyer Montegut等[47],该文中图5),我们发现,南海海盆的混合层深度很浅,且季节变化很小:夏季与北太平洋环流中心区(North Pacific Gyre,NPG)的MLD基本一致,此时南太平洋环流中心区(South Pacific Gyre,SPG)的MLD很深(大于100 m);而冬季又与SPG的MLD相当,此时NPG的MLD已增大到约100 m。对于远离河流输入的深海大洋,浮游植物的生长所需的营养盐主要依靠混合作用携带底层营养盐输入,而南海常年较浅的MLD难以为表层水体带来充足的营养盐补充。
(2)在这种情况下,MLD的增加反而会产生降低浮游植物生物量的效应:混合加强过程不仅直接稀释了浮游植物的浓度(与北大西洋秋初的混合层加深导致的浮游植物生物量降低类似[48]),同时还减弱了混合层内的生长光照Ig,从而降低了浮游植物的光合效率。由于MLD的加深通常伴随风速的增加以及SST的降低,因而导致在海盆区bbp与物理参数的相关性与近岸的VCS正好相反:bbp负相关于风速和MLD、而正相关于SST。在近岸浅水区,混合过程的加强可迅速增加表层的营养盐,从而增加浮游植物的生物量(同时也增加了无机颗粒物的再悬浮);但在深水海盆区,混合过程的加强难以向表层补充营养盐,但稀释与降低光照的作用则导致浮游植物生物量减小。
(3)由于MLD全年变化幅度并不大,bbp虽有夏高冬低的趋势但也并不十分显著。与此同时,叶绿素a浓度则由于浮游植物对光照变化的生理学响应而表现出高光条件下(夏季)减小、低光条件下(冬季)增多,从而在季节尺度上与bbp表现出负相关关系。此外,相关性分析还发现,叶绿素a浓度与物理参数的相关性在NDB和SDB要高于bbp与物理参数的相关性,也是由于浮游植物的生理学过程导致的。由于叶绿素a浓度随混合层内的生长光照Ig变化,而南海光照充足且季节变化不大,Ig的季节特征主要受到MLD的调制,因此叶绿素a浓度与物理参数的相关性比bbp更高。
3.3 物理与生态参量相关分析的动力学解释
从动力学角度,海表物质浓度与海面风速和海表温度的相关分析结果,可以用于判断该种物质的变化是受次表层过程(如垂向混合作用的跨等密度面输送)的影响还是受表层过程(如平流作用的沿等密面输送)的影响。
通过表1分析,在MRP和PRP两个河口羽流区,叶绿素a浓度和bbp与SSW均为正相关,说明两者都受到一定的次表层过程影响;但SST与叶绿素a浓度正相关,而与bbp负相关,说明叶绿素a浓度还受到表层河口羽流等沿等密面输运的控制;特别是叶绿素a浓度与SSW的正相关性非常弱,说明叶绿素a浓度可能受平流作用的影响更大。如前文所述,河口羽流区的特点是虽然bbp与叶绿素a浓度在一定程度上存在共变性,但两者的来源并不完全一致。河口区悬浮泥沙含量较高,导致颗粒物后向散射的数据中除浮游植物外,还包含了无机物的信息。
在SDB和NDB 两个深水海盆区,叶绿素a浓度 与SSW正相关,与SST负相关,说明叶绿素a浓度受到垂向混合的控制为主;但bbp与SSW负相关,与SST正相关,说明bbp仅受表层过程的影响。如前文所述,深水海盆区的浮游植物由于混合层较浅,难以通过垂向混合过程带来下层的营养盐补充,因此浮游植物以及颗粒物的季节变化受环流水平输运的影响为主。而叶绿素a浓度则由于受到浮游植物生理学过程的调制,表现出与垂向混合作用(调制混合层内的生长光照)具有更为密切的关系。
海盆边缘的VCS和WLS两个海区则表现出明显的次表层影响。叶绿素a浓度与SSW正相关,与SST负相关,说明叶绿素a浓度主要受到垂向混合作用的控制,这一特征与海盆区一致。而bbp也是与SSW正相关而与SST负相关,说明bbp也受垂向混合的控制,与河口区一致。叶绿素a浓度与bbp两个生态参数的季节变化都受跨等密面的次表层过程主导,这是由于VCS的浅水特征以及WLS的上升流导致的。值得注意的是,叶绿素a浓度和bbp之间的相关性在这两个海区是所有典型区域中最高的,也说明主导两者季节变化的动力学过程是非常相似的。
3.4 南海海盆区浮游植物的光驯化特征
Behrenfeld等[27]首次提出CChla/Cphy随光照变化的浮游植物光驯化模型——PhotoAcc模型(见公式(4)),并应用到全球遥感数据中[23]。CChla/Cphy与Ig的负相关性表现出光照对于叶绿素a浓度的调制作用,而营养盐与温度的调制作用则体现在回归系数中(公式(4)中的回归系数A表征了营养盐的限制作用,而B+A表征了温度的限制作用)[23]。基于这一模型,Siegel等[3]通过卫星数据以及邢小罡等[39]、Xing等[49]通过现场数据分别研究了全球和局地尺度的光驯化特征,而Westberry等[50]则将其应用到垂向估算浮游植物生物量与初级生产力的遥感模型中。本文分析南海浮游植物光驯化特征的方法与Behrenfeld等[23]一致,首先通过公式(1)和(2)计算南海气候态月平均的生长光照Ig,以及通过公式(3)估算浮游植物碳含量Cphy,最后基于公式(4)回归分析得到图中每个点处的季节尺度上CChla/Cphy与Ig的经验回归关系(即得到回归系数A和B)。
图4a显示了6个典型站位的回归曲线。3个近岸区(MRP、PRP、VCS)的斜率B都为负值,说明近岸区的CChla/Cphy并没有表现出光驯化的特征,与光照基本上不存在相关性。而在3个深水区(WLS、NDB、SDB),图4a则表现出明显的CChla/Cphy与Ig的负相关关系,斜率B均为正值,在低光条件下,CChla/Cphy显著升高。这一结果说明,通过PhotoAcc模型回归得到的斜率B值可以用于评价生理学过程对浮游植物叶绿素的影响程度。整个南海斜率值B的空间分布如图4b所示,近岸海区的B值一般都在0左右甚至为负值(白色和蓝色),说明光驯化特征不明显,这与图2c显示出近岸海区叶绿素a浓度与bbp强相关性是一致的,具有“生物量控制区”的特点;而在海盆区,斜率B几乎都为正值,高值区出现在北部深水海盆NDB,是典型的“光驯化控制区”。两个控制区的分界线恰好与陆架-海盆分界线一致(图4b),体现了水深条件对于浮游植物生理状态的影响。水深的差别反映了营养盐补充程度的差异:浅水区的混合过程相对较强,加之河流输入以及近岸上升流的作用,表层水体的营养盐可以得到充分的补充;而在深水区,表层浮游植物生长所需的营养盐主要来自于底层向上的输入,由于层化强烈,底层营养盐一般难以补充到表层水体。而营养盐的补充程度则对浮游植物生理状态产生重要的影响[8]:在浅水区,丰富的营养盐环境使浮游植物体内的叶绿素a浓度与生物量同步增长,比例变化不大;而在营养盐限制的海盆区,浮游植物体内的叶绿素a则对光照的变化非常敏感。
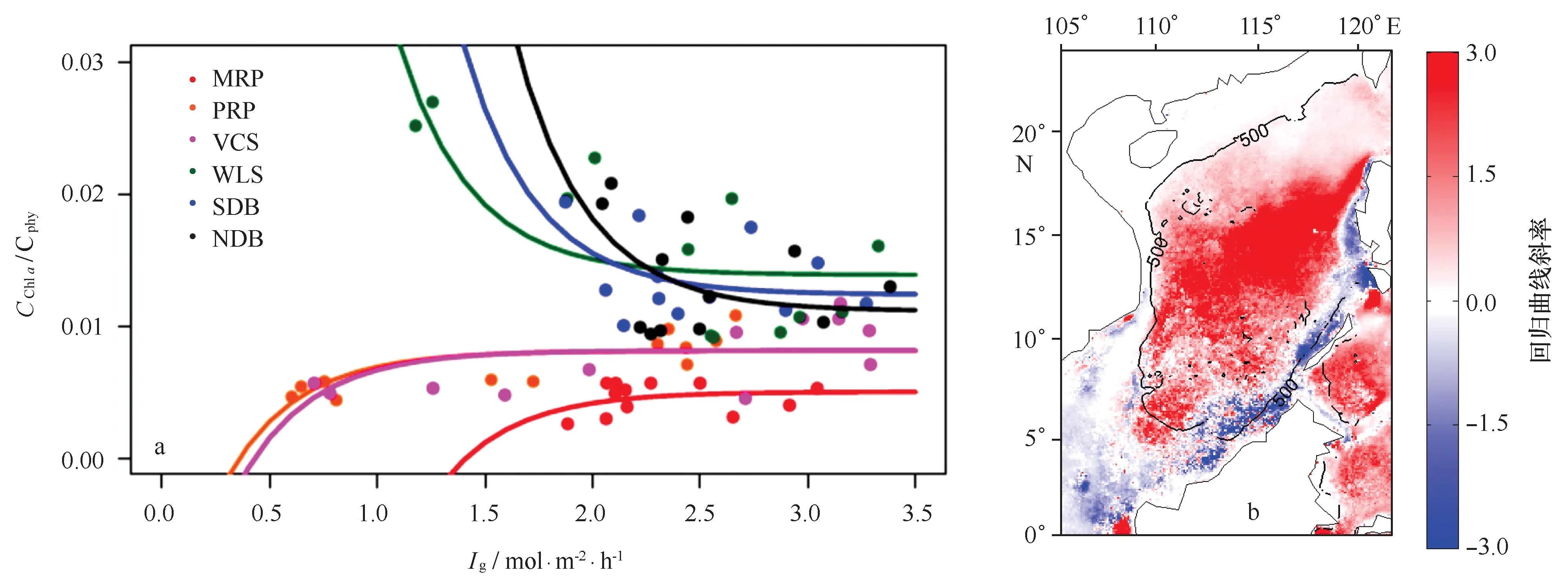
图4 a. 南海6个典型海区的叶绿素a浓度与浮游植物碳含量的比值(CChl a /Cphy)与生长光照(Ig)的散点图与回归曲线(湄公河口:MRP;珠江口:PRP;越南东部陆架:VCS;吕宋海峡西侧:WLS;南部深水海盆:SDB;北部深水海盆:NDB);b.南海两者回归曲线斜率的空间分布(黑色实线为500 m等深线)Fig.4 a. Scatter plot and regressed curve between the ratios of chlorophyll a concentration to phytoplankton carbon (CChl a /Cphy) and corresponding growth irradiance (Ig) in 6 typical regions of the South China Sea (Mekong River Plume,MRP; Pearl River Plume,PRP; Vietnamese Continental Shelf,VCS; West of Luzon Strait,WLS; Southern Deep Basin,SDB; Northern Deep Basin,NDB); b. Spatial distribution of regressed slopes in the South China Sea(the black solid lines represent the 500 m depth contour)
值得注意的是,在WLS海区,虽然叶绿素a浓度与bbp是良好耦合的,但光驯化特征仍然非常显著。从相关分析的结果来看(见图2c,d),WLS海区的bbp正相关于叶绿素a浓度、负相关于SST,与近岸区是一致的;但通过PhotoAcc模型的回归系数B则表明该海区存在显著的生理学变化特征,与海盆区是类似的(图4b)。尤其在冬季藻华期(11-4月),叶绿素a浓度与bbp的相关系数达到0.98,按照Siegel等[3]的定义,应属于“生物量控制区”;但此时CChla/Cphy与exp(-3Ig)的相关系数也达到0.83,且其生理学调整的程度(即CChla/Cphy的变化幅度)还要大于NDB和SDB(图4a)。因此,Siegel等[3]简单地通过相关性的定义并不能完整描述WLS的特点,该海区实际上表现出“生物量-光驯化共同控制”的特点。
需要指出,本文从叶绿素与生物量(以颗粒物后向散射系数代表)变化的差异性来计算的“光驯化现象”,是一种针对浮游植物群落整体的“表观光驯化”。CChla/Cphy的变化除了浮游植物的光驯化特性以外,还会受到浮游植物种群分布的影响。由于浮游植物种间的差异,仅浮游植物的群落演替也可能表现出“表观光驯化”的效果。南海沿岸区的浮游植物以硅藻、甲藻等较大细胞个体藻类为主要类群,而在深水海盆区则以原绿球藻等微微型藻类为主,后者细胞中以二乙烯基叶绿素为主要色素,因此不同区域CChla/Cphy的变化还与浮游植物的种群分布有关。而在时间尺度上,浮游植物的群落也存在季节更替,例如,微微型藻类在夏季群落中的百分比通常比冬季更高[51],因此也可以导致冬季的CChla/Cphy会高于夏季。然而考虑到藻种比例的变化不太可能导致叶绿素a浓度与bbp在季节变化过程上完全相反(如NDB和SDB),因此,浮游植物的光驯化过程仍然认为是导致上述参量在深水海盆区负相关关系的主要原因。
4 结论
利用卫星反演的多种生物光学参数以及其他大时空尺度的海洋与气象参数,研究各种参数间时空变化的耦合性与差异性,将不断加深我们对局地海洋生态系统及其动态变化过程的理解。本文基于MODIS卫星遥感的南海叶绿素a浓度与颗粒物后向散射系数数据,详细分析了两者在季节尺度上的相关性与独立性、及其与物理参数的耦合关系,首次讨论了南海表层水体的光驯化特征。
研究发现,南海的“生物量控制区”主要集中在近岸浅水区,表现为叶绿素a浓度与bbp较高的正相关性以及CChla/Cphy与Ig的弱相关性,说明近岸区叶绿素a浓度的变化主要受到浮游植物生物量本身变化的影响;而“光驯化控制区”则主要集中在深水海盆区,表现为叶绿素a浓度与bbp的负相关性以及CChla/Cphy与Ig的负相关性,说明深水区叶绿素a浓度的季节变化受浮游植物生理学过程的影响更大,导致其季节循环过程甚至与浮游植物生物量的变化过程相反。两个区域的分界线基本与南海陆架-海盆分界线一致,说明水深的差异——反映营养盐补充的程度——对于浮游植物生理状态的影响。
此外,本文还发现WLS海区存在“生物量-光驯化共同控制”的特点——叶绿素a浓度与bbp的正相关性很高,但CChla/Cphy与Ig的负相关性也很高,这种海区特征在之前的研究中尚未被提及。这一特点是否与其特殊的局地物理过程有关,在全球其他海区是否有类似情况出现,值得我们做更多的分析与研究。
[1] Field C B,Behrenfeld M J,Randerson J T,et al. Primary production of the biosphere: Integrating terrestrial and oceanic components[J]. Science,1998,281(5374): 237-240.
[2] Behrenfeld M J,Randerson J T,McClain C R,et al. Biospheric primary production during an ENSO transition[J]. Science,2001,291(5513): 2594-2597.
[3] Siegel D A,Behrenfeld M J,Maritorena S,et al. Regional to global assessments of phytoplankton dynamics from the SeaWiFS mission[J]. Remote Sensing of Environment,2013,135: 77-91.
[4] Garver S A,Siegel D A. Inherent optical property inversion of ocean color spectra and its biogeochemical interpretation: I. Time series from the Sargasso Sea[J]. J Geophys Res,1997,102(C8): 18607-18625.
[5] Maritorena S,Siegel D A,Peterson A R. Optimization of a semianalytical ocean color model for global-scale applications[J]. Applied Optics,2002,41(15): 2705-2714.
[6] Lee Z P,Carder K L,Arnone R A. Deriving inherent optical properties from water color: a multiband quasi-analytical algorithm for optically deep waters[J]. Applied Optics,2002,41(27): 5755-5772.
[7] Werdell P J,Franz B A,Bailey S W,et al. Generalized ocean color inversion model for retrieving marine inherent optical properties[J]. Applied Optics,2013,52(10): 2019-2037.
[8] Siegel D A,Maritorena S,Nelson N B,et al. Independence and interdependencies among global ocean color properties: reassessing the bio-optical assumption[J]. J Geophys Res,2004,110: C07011.
[9] 赵辉,齐义泉,王东晓,等. 南海叶绿素浓度季节变化及空间分布特征研究[J]. 海洋学报,2005,27(4): 45-52.
Zhao Hui,Qi Yiquan,Wang Dongxiao,et al. Study on the features of chlorophylladerived from SeaWiFS in the South China Sea[J]. Haiyang Xuebao,2005,27(4): 45-52.
[10] 赵辉,唐丹玲,王素芬. 南海西北部夏季叶绿素a浓度的分布特征及其对海洋环境的响应[J]. 热带海洋学报,2005,24(6): 31-37.
Zhao Hui,Tang Danling,Wang Sufen. Spatial distribution of chlorophyll a concentration in summer in western South China Sea and its response to oceanographic environmental factors[J]. Journal of Tropical Oceanography,2005,24(6): 31-37.
[11] Tang D L,Ni I H,Kester D R,et al. Remote sensing observations of winter phytoplankton blooms southwest of the Luzon Strait in the South China Sea[J]. Mar Ecol Prog Ser,1999,191: 43-51.
[12] Gao S,Wang H. Seasonal and spatial distributions of phytoplankton biomass associated with monsoon and oceanic environments in the South China Sea[J]. Acta Oceanologica Sinica,2008,27(6): 17-32.
[13] 林丽茹,赵辉. 南海海域浮游植物叶绿素与海表温度季节变化特征分析[J]. 海洋学研究,2012,30(4): 46-54.
Lin Liru,Zhao Hui. Analysis on the relations between sea surface temperature and phytoplankton Chlorophyllain the South China Sea[J]. Journal of Marine Sciences,2012,30(4): 46-54.
[14] Gao S,Wang H,Liu G M,et al. Spatio-temporal variability of chlorophyllaand its responses to sea surface temperature,winds and height anomaly in the western South China Sea[J]. Acta Oceanologica Sinica,2013,32(1): 48-58.
[15] 马翱慧,刘湘南,李婷,等. 南海北部海域叶绿素a浓度时空特征遥感分析[J]. 海洋学报,2013,35(4): 98-105.
Ma Aohui,Liu Xiangnan,Li Ting,et al. The satellite remotely-sensed analysis of the temporal and spatial variability of chlorophyllaconcentration in the northern South China Sea[J]. Haiyang Xuebao,2013,35(4): 98-105.
[16] Martinez E,Antoine D,D’Ortenzio F,et al. Climate-driven basin-scale decadal oscillations of oceanic phytoplankton[J]. Science,2009,326(5957): 1253-1256.
[17] Henson S A,Sarmiento J L,Dunne J P,et al. Detection of anthropogenic climate change in satellite records of ocean chlorophyll and productivity[J]. Biogeosciences,2010,7(2): 621-640.
[18] Boyce D G,Lewis M R,Worm B. Global phytoplankton decline over the past century[J]. Nature,2010,466(7306): 591-596.
[19] Laws E A,Bannister T T. Nutrient-and light-limited growth of Thalassiosira fluviatilis in continuous culture,with implications for phytoplankton growth in the ocean[J]. Limnol Oceanogr,1980,25(3): 457-473.
[20] Geider R J. Light and temperature dependence of the carbon to chlorophyllaratio in microalgae and cyanobacteria: implications for physiology and growth of phytoplankton[J]. New Phytol,1987,106(1): 1-34.
[21] Falkowski P G,LaRoche J. Acclimation to spectral irradiance in algae[J]. J Phycol,1991,27(1): 8-14.
[22] Lewis M R,Cullen J J,Platt T. Relationships between vertical mixing and photoadaptation of phytoplankton: similarity criteria[J]. Marine Ecology Progress Series,1984,15: 141-149.
[23] Behrenfeld M J,Boss E,Siegel D A,et al. Carbon-based ocean productivity and phytoplankton physiology from space[J]. Global Biogeochemical Cycles,2005,19(1): GB1006.
[24] 袁梁英. 南海北部营养盐结构特征[D]. 厦门: 厦门大学,2005.
Yuan Liangying. Distribution and characteristics of nutrients in the northern South China Sea[D]. Xiamen: Xiamen University,2005.
[25] Frouin R,Chertock B. A technique for global monitoring of net solar irradiance at the ocean surface. Part Ⅰ: Model[J]. J Appl Meteor,1992,31(9): 1056-1066.
[26] Morel A,Huot Y,Gentili B,et al. Examining the consistency of products derived from various ocean color sensors in open ocean (Case 1) waters in the perspective of a multi-sensor approach[J]. Remote Sensing of Environment,2007,111(1): 69-88.
[27] Behrenfeld M J,Maranon E,Siegel D A,et al. Photoacclimation and nutrient-based model of light-saturated photosynthesis for quantifying oceanic primary production[J]. Mar Ecol Prog Ser,2002,228: 103-117.
[28] Liu Q Y,Jiang X,Xie S P,et al. A gap in the Indo-Pacific warm pool over the South China Sea in boreal winter: seasonal development and interannual variability[J]. J Geophys Res,2004,109: C07012.
[29] Xie S P,Xie Q,Wang D X,et al. Summer upwelling in the South China Sea and its role in regional climate variations [J]. J Geophys Res,2003,108(C8): 3261.
[30] 刘昕,王静,程旭华,等. 南海叶绿素浓度的时空变化特征分析[J]. 热带海洋学报,2012,31(4): 42-48.
Liu Xin,Wang Jing,Cheng Xuhua,et al. The temporal and spatial evolution of chlorophyllaconcentration in the South China Sea[J]. Journal of Tropical Oceanography,2012,31(4): 42-48.
[31] Wang J J,Tang D L,Sui Y. Winter phytoplankton bloom induced by subsurface upwelling and mixed layer entrainment southwest of Luzon Strait[J]. Journal of Marine Systems,2010,83(3/4): 141-149.
[32] Ning X,Chai F,Xue H,et al. Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea[J]. J Geophys Res,2004,109: C10005.
[33] Chen Y L. Spatial and seasonal variations of nitrate-based new production and primary production in the South China Sea[J]. Deep-Sea Res Ⅰ,2005,52(2): 319-340.
[34] 吴日升,李立. 南海上升流研究概述[J]. 台湾海峡,2003,22(2): 269-277.
Wu Risheng,Li Li. Summarization of study on upwelling system in the South China Sea[J]. Journal of Oceanography in Taiwan Strait,2003,22(2): 269-277.
[35] Qu T D,Du Y,Gan J P,et al. Mean seasonal cycle of isothermal depth in the South China Sea[J]. J Geophys Res,2007,112: C02020.
[36] 高姗,王辉,刘桂梅,等. 南海叶绿素a浓度垂直分布的统计估算[J]. 海洋学报,2010,32(4): 168-176.
Gao Shan,Wang Hui,Liu Guimei,et al. The statistical estimation of the vertical distribution of chlorophyllaconcentration in the South China Sea[J]. Haiyang Xuebao,2010,32(4): 168-176.
[37] 柯志新,黄良民,谭烨辉,等. 2008年夏末南海北部叶绿素a的空间分布特征及其影响因素[J]. 热带海洋学报,2013,32(4): 51-57.
Ke Zhixin,Huang Liangmin,Tan Yehui,et al. Spatial distribution of chlorophyllaand its relationships with environmental factors in northern South China Sea in late summer 2008[J]. Journal of Tropical Oceanography,2013,32(4): 51-57.
[38] 柯志新,黄良民,谭烨辉,等. 2007年夏季南海北部浮游植物的物种组成及丰度分布[J]. 热带海洋学报,2011,30(1): 131-143.
Ke Zhixin,Huang Liangmin,Tan Yehui,et al. Species composition and abundance of phytoplankton in the northern South China Sea in summer 2007[J]. Journal of Tropical Oceanography,2011,30(1): 131-143.
[39] 邢小罡,赵冬至,王林,等. 南太平洋副热带环流区次表层叶绿素极大值的季节变化特征[J]. 高技术通讯,2013,23(10): 1038-1047.
Xing Xiaogang,Zhao Dongzhi,Wang Lin,et al. Seasonal variation characteristics of subsurface chlorophyll maximum in the South Pacific subtropical gyre[J]. Chinese High Technology Letters,2013,23(10): 1038-1047.
[40] 邢小罡,丘仲锋,王林. Bio-Argo浮标观测贫营养水体浮游植物的光适应特性[J]. 激光生物学报,2014,23(6): 523-533.
Xing Xiaogang,Qiu Zhongfeng,Wang Lin. Characterization of phytoplankton photoadaptation in oligotrophic waters observed by Bio-Argo Floats[J]. Acta Laser Biology Sinica,2014,23(6): 523-533.
[41] Mignot A,Claustre H,Uitz J,et al. Understanding the seasonal dynamics of phytoplankton biomass and the deep chlorophyll maximum in oligotrophic environments: a Bio-Argo float investigation[J]. Global Biogeochemical Cycles,2014,28,doi: 10.1002/2013GB004781.
[42] 张强,崔瑛,陈永勤. 基于水文学方法的珠江流域生态流量研究[J]. 生态环境学报,2010,19(8): 1828-1837.
Zhang Qiang,Cui Ying,Chen Yongqin. Evaluation of ecological instream flow of the Pearl River basin,south China[J]. Ecology and Environmental Sciences,2010,19(8): 1828-1837.
[43] 钟华平,王建生. 湄公河干流径流变化及其对下游的影响[J]. 水利水运工程学报,2011(3): 48-52.
Zhong Huaping,Wang Jiansheng. An analysis of Mekong River runoff change and its impact on the Mekong River downstream[J]. Hydro-Science and Engineering,2011(3): 48-52.
[44] 刘成,胡春宏,史红玲. 中国大陆太平洋流域河流下泄水沙通量的变化[J]. 泥沙研究,2011(1): 70-75.
Liu Cheng,Hu Chunhong,Shi Hongling. Changes of runoff and sediment fluxes of rivers in mainland of China discharged into Pacific Ocean[J]. Journal of Sediment Research,2011(1): 70-75.
[45] Morel A,Ahn Y H. Optics of heterotrophic nanoflagellates and ciliates: a tentative assessment of their scattering role in oceanic waters compared to those of bacterial and algal cells[J]. J Mar Res,1991,49(1): 177-202.
[46] Stramski D,Kiefer D A. Light scattering by microorganisms in the open ocean[J]. Prog Oceanogr,1991,28(4): 343-383.
[47] De Boyer Montegut C,Madec G,Fischer A S,et al. Mixed layer depth over the global ocean: an examination of profile data and a profile-based climatology[J]. J Geophys Res,2004,109: C12003.
[48] 邢小罡,邱国强,王海黎. Bio-Argo浮标观测北大西洋色素与颗粒物的季节分布[J]. 高技术通讯,2014,24(1): 55-64.
Xing Xiaogang,Qiu Guoqiang,Wang Haili. Seasonal distributions of pigment and particle in the North Atlantic observed by a Bio-Argo float[J]. High Technology Letters,2014,24(1): 55-64.
[49] Westberry T K,Behrenfeld M J,Siegel D A,et al. Carbon-based primary productivity modeling with vertically resolved photoacclimation[J]. Global Biogeochemical Cycles,2008,22: GB2024.
[50] Xing X G,Claustre H,Uitz J,et al. Seasonal variations of bio-optical properties and their interrelationships observed by Bio-Argo floats in the subpolar North Atlantic[J]. J Geophys Res,2014,119(10): 7372-7388.
[51] 朱根海,宁修仁,蔡昱明,等. 南海浮游植物种类组成和丰度分布的研究[J]. 海洋学报,2003,25(S2): 8-23.
Zhu Genhai,Ning Xiuren,Cai Yuming,et al. Studies on species composition and abundance distribution of phytoplankton in the South China Sea[J]. Haiyang Xuebao,2003,25(S2): 8-23.
A satellite-based analysis on the seasonal variations and inter-relationships between chlorophyll and particle in the South China Sea
Wang Xiaoqi1,Xing Xiaogang1,Wang Jinping1,Li Na1
(1.CollegeofPhysicalandEnvironmentalOceanography,OceanUniversityofChina,Qingdao266100,China)
On the basis of satellite remotely sensed monthly chlorophyllaconcentration and particle backscattering coefficient,as well as other oceanographic and meteorological parameters,this study analyzed on the correlation coefficients between concentration of chlorophyllaand particulate backscattering on seasonal scale and their coupling relationships with physical parameters in the South China Sea. In addition,a phytoplankton photoacclimation model was utilized to identify the physiological characteristics. The results showed that,due to the influences of specific topography,wind forcing and other physical parameters,both consistency and discrepancy between the seasonal cycles of chlorophyllaand particle backscattering coefficient were found in the South China Sea. In coastal and shallow waters,two properties were strongly coupled; while in the central deep-water basin,chlorophyllawas higher in winter and lower in summer,totally differing from the seasonal evolution of particles,such phenomenon was mainly owing to phytoplankton physiological process. The boundary of “biomass regime” and “photoacclimation regime” in the South China Sea located at the shelf-basin interface,suggesting the influence of water depth on the phytoplankton physiological status. Furthermore,a characteristics of “biomass-photoacclimation joint controlling” was found in the west of Luzon Strait.
chlorophyllaconcentration; particle backscattering coefficient; South China Sea; seasonal scale; photoacclimation
2015-02-03;
2015-08-04。
国家基础科学人才培养基金(J1103402,J1310013)。
王晓琦(1993—),女,山东省莱州市人,物理海洋专业。E-mail:xiaran93@foxmail.com
*通信作者:邢小罡(1982—),男,山东省青岛市人,博士,副教授,主要从事海洋水色遥感方向研究。E-mail:xing@ouc.edu.cn
10.3969/j.issn.0253-4193.2015.10.003
TP79; Q948.8
A
0253-4193(2015)10-0026-13
王晓琦,邢小罡,王金平,等. 基于遥感数据分析南海叶绿素与颗粒物的季节变化与相互关系[J].海洋学报,2015,37(10):26—38,
Wang Xiaoqi,Xing Xiaogang,Wang Jinping,et al. A satellite-based analysis on the seasonal variations and inter-relationships between chlorophyll and particle in the South China Sea[J]. Haiyang Xuebao,2015,37(10):26—38,doi:10.3969/j.issn.0253-4193.2015.10.003
猜你喜欢
海洋信息技术与应用(2021年3期)2022-01-17
石油钻探技术(2020年3期)2020-08-24
海洋学报(2020年7期)2020-08-06
建筑热能通风空调(2018年5期)2018-07-09
海洋信息技术与应用(2017年2期)2017-06-21
中国工程咨询(2016年9期)2016-02-13
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
天然气勘探与开发(2014年4期)2014-02-28
