
大豆幼苗对套作玉米遮荫环境的光合生理生态响应
2015-02-08 05:26苏本营宋艳霞陈圣宾杨文钰
生态学报 2015年10期
苏本营, 宋艳霞, 陈圣宾, 杨文钰
1 四川农业大学农学院农业部西南作物生理生态与耕作重点实验室, 成都 611130 2 四川省安岳县农业技术推广中心, 资阳 642350 3 环境保护部南京环境科学研究所, 南京 210042
大豆幼苗对套作玉米遮荫环境的光合生理生态响应
苏本营1, 宋艳霞2, 陈圣宾3, 杨文钰1,*
1 四川农业大学农学院农业部西南作物生理生态与耕作重点实验室, 成都 611130 2 四川省安岳县农业技术推广中心, 资阳 642350 3 环境保护部南京环境科学研究所, 南京 210042
以2个耐荫性不同的大豆品种为材料,田间试验设置大豆单作和玉米-大豆套作2个种植模式处理,研究不同耐荫性大豆品种的幼苗光合生理生态特性对套作玉米遮荫环境的响应。结果表明:1)玉米-大豆套作模式中,玉米遮荫显著降低大豆冠层的光合有效辐射,导致大豆幼苗光合速率、气孔导度、蒸腾速率显著下降(P< 0.05),分别达37.9%、54.2%和42.4%,但品种间无显著差异;而胞间二氧化碳浓度和Fv/Fm无显著变化,且光合速率的下降与气孔导度存在显著相关关系,光合速率下降主要是由气孔限制和CO2同化过程中能量不足所致;2) 玉米遮荫显著降低大豆幼苗叶面积指数、叶片碳含量、叶片和根系干重及总生物量,且品种间差异显著,相关性分析显示,叶面积指数下降是导致生物量减少的主要原因;3) 玉米遮荫环境中,大豆幼苗的叶片叶绿素和氮素含量提高以增强光捕获能力,但它们并不能补偿因光截获面积降低而引起的光截获量下降。
间套作; 遮荫; 气体交换; 光合色素; 干物质积累
间套作在没有扩大土地面积的前提下提高了粮食产量,为解决世界人口的温饱问题做出了重要贡献[1- 2]。研究表明,间套作种植能够有效提高资源利用率和粮食产量[3- 5],增强农业系统的抗风险能力[6],增加水土保持能力[7],提高土壤肥力[8],同时能够抑制病虫草害的发生[9- 11],是一种基于生物多样性的生态农业范式[11- 12]。在当前全球环境变化加剧大背景下,间套作种植模式的研究越来越受到关注[13- 15]。在我国西南丘陵地区,玉米-大豆带状套作种植近年发展迅速,四川省自2003年开始推广以来,种植面积已达115.1万hm2,农民增加收益60.65亿元,显著提高了区域粮食生产能力[16]。
株高不同的作物搭配构成的间套复合群体中,高位作物对低位作物造成的遮荫将会影响到低位作物的生长发育,进而影响到间套复合系统的生产力[17- 21]。国内外在高位作物对低位作物形态发育和生理特征影响方面已有相关报道,不同荫蔽程度、不同作物间均存在较大差异[21- 27]。在玉米-大豆套作模式中,大豆播种于玉米行间,幼苗出土后便生长在玉米荫蔽环境中直至玉米收获,共生期约60d左右,玉米荫蔽环境将会导致套作大豆光合有效辐射截获减少,叶片结构发生变化[28- 29],茎秆变细变长[30],这将可能伴随着一系列生理生态特性的适应,却尚未见相关报道。本文以耐阴性不同的大豆品种为研究对象,从光合生理生态的适应性角度探讨大豆幼苗对玉米遮荫环境的响应,旨在为大豆耐荫品种选育提供理论依据,同时为玉米-大豆套作技术完善和应用推广提供技术支持。
1 材料与方法
1.1 研究地点概况
试验于2012年在四川农业大学教学科研农场进行,研究区位于四川省雅安市雨城区(29°59′N,103°00′E)。该区域属于亚热带湿润季风气候,年平均气温16.2 ℃,月最低气温6.1 ℃,月最高气温25.4 ℃,无霜期280—310 d;降水资源丰富,年降雨量1250—1750 mm;光资源相对不足,属于全国太阳辐射较低的区域,年均日照时数仅1005 h,太阳总辐射3750 MJm-2a-1左右。试验地土壤类型为紫色粘土类型,pH值约7.5,有机质含量29.8 g/kg,全氮1.3 g/kg,全磷0.5 g/kg,全钾26.2 g/kg,速效氮176.9 mg/kg,速效磷38.1 mg/kg,速效钾129.6 mg/kg。
1.2 试验设计
试验采用双因素随机区组设计,每小区面积6 m×6 m=36 m2,3次重复,共12个处理小区。主因素为种植方式:大豆单作(M)和大豆-玉米套作(RI);副因素为品种:南豆12(ND12)和南冬抗022- 2(NDK)。在玉米-大豆套作种植模式中,带宽2 m,玉米品种选用川单418(株型半紧凑、株高269 cm),由四川农业大学玉米研究所提供;大豆选择2个耐荫性存在差异的品种:南豆12和南冬抗022- 2,由四川省南充市农科院提供。玉米于4月初播种,采用南-北行向宽窄行种植方式,窄行行距为40 cm,穴距19 cm,每穴1苗,全田玉米密度为5.25×104株/hm2;大豆于6月初在玉米吐丝期播种2行于玉米宽行内,行距为40 cm,穴距19 cm,每穴2苗,全田密度为1.05×105株/hm2,大豆、玉米行间距离为60 cm。大豆单作种植模式中,大豆播种期与套作模式相同,行距50 cm,穴距19 cm,每穴1苗,密度1.05×105株/hm2,保证单株的生长空间与套作模式相同。试验以每公顷纯氮36 kg、P2O563 kg、K2O 52.5 kg做为大豆底肥施入,其他农艺措施管理同常规大田生产。
1.3 测定指标与方法
1.3.1 大豆生长微气候环境指标测定
选择晴朗天气(分别于大豆出苗后第30天、36天和43天)从6:00—18:00,每间隔2 h,在大豆幼苗冠层上方(冠层上方10 cm处)利用光量子传感器LI- 190 (LI-COR, Lincoln, USA) 进行入射光合有效辐射(PAR)日动态变化的测定(每小区选择6个点),同时利用HOBO H8温、湿度数据采集器(Onset Computer Corp, Bourne, USA)进行温度(TEMP)和相对湿度(RH)日变化的测定。
1.3.2 光合速率和叶绿素荧光参数测定
大豆出苗后第30、36和43天,从9:00—11:00,每处理小区选择6株大豆幼苗的功能叶(倒二叶),利用LI- 6400便携式气体交换测定系统 (Li-COR, Lincoln, USA)进行瞬时气体交换参数的测定。测定时光照采用人工光源,光强控制在1000 μmol m-2s-1,叶室内温度为24—28 ℃,相对湿度65%—75%。在测定气体交换参数的同时,利用Mini-PAM便携式脉冲调制式叶绿素荧光仪(Mini-PAM, Heinz Walz, Effeltrich, Germany) 测定最大荧光Fm和初始荧光F0,测定时将叶片充分的暗适应30min,每一处理小区选择有代表性的6株,测定其功能叶片(倒二叶)。可变荧光Fv和PSⅡ最大光化学量子产量Fv/Fm计算获得:Fv=Fm-F0;Fv/Fm= (Fm-F0)/Fm。
1.3.3 叶片采集和叶绿素含量测定
大豆出苗后第30、36和43天,每处理小区选择12株,取其功能叶(倒二叶)迅速放入液氮中,带回实验室储存于-40 ℃的冰箱待测。进行测定时,取冷冻鲜样0.2 g于20 mL 95%的乙醇溶液中研磨进行萃取,萃取液用于叶绿素含量的分析,Chla、Chlb和Car分析参考[31],Chls和Chla/b则通过计算获得。
1.3.4 叶面积指数、干物质积累和叶片C、N含量测定
于大豆出苗后第30、36和43天,每处理小区选择12株取样,用蒸馏水清洗后,区分成根、茎和叶,用叶面积扫描仪扫描叶片,并采用图像分析软件Scion Image计算叶面积,叶面积指数根据种植密度和叶面积计算获得。根、茎和叶于105 ℃杀青,80 ℃烘干至恒重,然后用电子天平(BP221S, Germany)称重,生物量和根冠比通过计算获得。称重后的部分叶片用小型高速粉碎机粉碎,过孔径0.246 mm筛,用于全碳、全氮含量的测定。全碳含量采用K2Cr2O7氧化还原滴定法[32],全氮含量采用凯氏定氮法[33],碳氮比(C/N)为全碳含量与全氮含量的比值。
1.4 统计分析
数据的统计分析应用SPSS软件(version15,SPSS, Chicago, IL, USA)进行。本研究中实验数据用单因素方差分析(one-way ANOVA)检验处理间差异的显著性,不同的字母表示差异显著(P< 0.05),相同字母表示差异不显著(P> 0.05)。不同参数之间的相关性采用Pearson相关矩阵进行分析。
2 结果和分析
2.1 玉米-大豆套作对大豆幼苗生长微环境的影响
随全天时间变化,套作(RI)和单作(M)种植条件下光合有效辐射(PAR)和温度(TEMP)均呈先增加后降低的趋势,而湿度(RH)则表现为与TEMP变化相反的趋势(图1)。6:00—18:00时间内,RI条件下PAR均显著低于M (P< 0.01),平均下降程度达52.5%;但TEMP和RH差异不显著(P> 0.05) (图1)。这表明RI条件下的玉米遮荫将改变大豆幼苗的生长环境,显著降低了大豆幼苗生长发育所必须的可利用光资源。

图1 单作(M)和套作(RI)环境下光合有效辐射、温度和相对湿度的日动态变化
2.2 玉米遮荫对大豆幼苗叶片叶绿素含量的影响
玉米遮荫能够显著提高大豆叶片Chla、Chlb和Chls的含量(图2)。与M相比,RI条件下,Chla、Chlb和Chls均显著升高(P< 0.05),2个大豆品种ND12和NDK的Chla分别升高了22.1%和18.1%,Chlb则分别升高了58.4%和49.4%,Chls分别升高了29.3%和24.4%。这表明,玉米遮荫诱导了大豆幼苗叶片合成更多的Chlb来吸收更多的散射光,提高光的吸收率。而Chla/b和Car则表现为RI低于M,且差异显著(P< 0.05)。2个大豆品种在3次测定时均表现一致,但品种间无显著差异(图2)。

图2 大豆幼苗叶片叶绿素a、叶绿素b、总叶绿素、类胡萝卜素含量和叶绿素a/b
2.3 玉米遮荫对大豆幼苗叶片C、N的影响
玉米遮荫显著提高了大豆幼苗叶片全氮(TON)含量(P< 0.05)(图3),这表明,玉米遮荫条件下,大豆幼苗将通过增加叶片氮含量来补偿因光照不足引起的光合速率下降。RI条件下,叶片全碳(TOC)含量和碳氮比(C/N)均显著低于M(P< 0.05)(图3),南冬抗022- 2(NDK)品种叶片TOC的下降程度更大(11.1%),显著高于南豆12(ND12)(3.8%)(P< 0.05)。

图3 大豆幼苗叶片氮含量、碳含量和碳氮比
2.4 玉米遮荫对大豆幼苗气体交换和叶绿素荧光参数的影响
套作环境下,大豆幼苗叶片净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)受玉米遮荫的影响,均显著低于单作(P< 0.05)(图4),平均下降程度分别为37.9%、54.2%和42.4%,品种间无显著差异(P> 0.05),且3次测定时表现一致,玉米遮荫所导致的Pn的下降程度与Gs和TOC的下降程度呈极显著的正相关关系,相关系数分别为0.654和0.764(表1)。而胞间二氧化碳浓度(Ci)和PSⅡ光化学效率(Fv/Fm) (图4)在净作和套作条件下,2品种在3次测定时均未表现出显著差异(P> 0.05)。
2.5 玉米遮荫对大豆幼苗叶面积指数的影响
由图5可以看出,M件下,2大豆品种幼苗的叶面积指数(LAI)无显著差异,玉米遮荫显著降低大豆幼苗的LAI,降低程度平均高达57.1%。套作环境中NDK幼苗的LAI,3次测定值分别为0.11、0.28和0.47,显著低于ND12(0.17、0.37和0.58)和单作条件下的LAI(0.29、0.91和1.19),表明ND12的耐荫性优于NDK。
2.6 玉米遮荫对大豆幼苗干物质积累与分配的影响
由图6可以看出,RI环境中,玉米遮荫显著降低了大豆幼苗的茎秆干重(SDM)(P< 0.05),且NDK的降低程度(69%)显著高于ND12(59%)。不同种植模式下,不同大豆品种的叶片干重(LDM)、根系干重(RDM)和总生物量(BIOMASS)存在显著的差异,具体表现为:M-NDK > M-ND12 > RI-ND12 > RI-NDK,玉米遮荫导致的LDM、RDM和BIOMASS的下降程度,NDK(78.5%、81.9%、75.3%)显著高于ND12(66.2%、71.3%和64.1%)(P< 0.05)(图6),表明ND12的耐荫性优于NDK。玉米遮荫所导致的BIOMASS下降程度与LAI、SDM、LDM、RDM的下降程度和Chls含量的变化程度存在极显著的正相关关系(表1),而与Pn不存在显著的相关关系。玉米遮荫还显著降低了大豆幼苗的根冠比(Root/Shoot)(P< 0.05),但品种间差异不显著(图6)。
3 讨论
间套作作为一种基于生物多样性的可持续农业发展模式,具有多种重要的生态服务功能[2, 14]。然而高位作物(玉米)的遮荫将会改变低位作物(大豆)的微生长环境,进而影响到系统的生产力[18- 21],因此理解低位作物对系统微环境变化的适应机理,实现共生作物和谐发展和双高产是间套作模式推广和发展的关键。本研究发现,在玉米-大豆套作模式中,玉米遮荫降低了抵达大豆冠层的光合有效辐射(PAR),下降程度高达52.5%(图1),这将引起大豆幼苗Pn下调,同时Gs和Tr下降(图4),与前人的研究结果一致[19, 29]。光合作用是物质代谢和能量转化的最初源泉,受植物自身生理特性和环境的共同影响[34- 35]。光环境对植物的直接作用就是光合作用,而关于荫蔽引起光合速率下降的原因国内外研究者存在一定的争议,有研究认为是气孔限制引起[36- 37]。也有研究认为是非气孔限制引起[38- 39],主要是包括叶片叶绿素和可溶性蛋白含量降低导致捕获光合电子的能力下降[39],PSⅡ损伤导致电子传输能力降低[39]或叶绿体的结构、数目和功能下降所引起[40]。还有研究认为是气孔限制和非气孔限制共同作用的结果引起[41]。本研究发现套作条件下,Pn的下降程度与Gs的下降存在显著的相关关系(表1),这表明玉米遮荫引发的大豆幼苗叶片气孔导度降低可能是引起光合速率下降的一个原因。另外,与单作相比,套作条件下大豆幼苗叶片Ci和Fv/Fm无显著的差异(图4),这意味着玉米遮荫并未造成PSⅡ受到损伤,Pn的下降可能由CO2同化过程中的能量不足引起,而与光反应过程无关[42]。因此,RI环境中的玉米遮荫导致的Pn降低可能是由气孔限制和CO2同化过程中的能量不足共同引起,对于其具体的影响过程还有待于更深入的研究。
外界环境发生变化时,植物将会改变自身形态、结构和生理生化特性来适应环境,以便维持最佳的生长发

图4 大豆幼苗叶片光合速率、气孔导度、胞间二氧化碳浓度、蒸腾速率和PSⅡ光化学效率
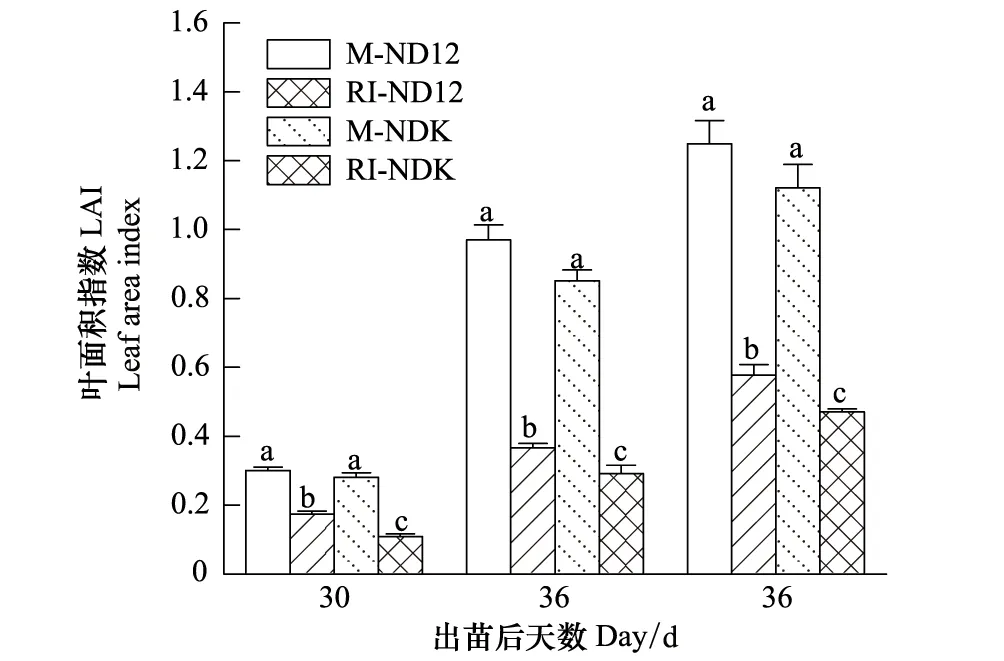
图5 大豆幼苗的叶面积指数
育水平[43]。弱光环境中,植物叶片将会调整自身结构和生理特征来吸收利用更多的光来维持生长发育,如叶片变薄、叶面积变大[28]、光合色素含量增加[22- 23,25]、叶片N含量升高[44]等。本研究中套作荫蔽环境会引起大豆幼苗叶片Chla、Chlb、Chls(图2)和TON含量(图3)升高来吸收更多的散射光提高光能的捕获能力[22- 23]。然而,叶片TOC含量、LAI和LDM的降低则使套作环境下大豆幼苗Chlb和TON含量升高所引起的光能捕获能力增强并不能补偿玉米遮荫所导致的光截获面积降低而引起的光截获量下降[45]。玉米遮荫引起的叶片TOC含量的下降程度与Pn的下降程度存在极显著的相关关系(表1),说明TOC含量的下降可能主要是由玉米遮荫导致的大豆幼苗Pn降低所引起。而LAI和LDM的下降可能主要是由弱光环境下大豆幼苗叶片形成的数量较低所引起[45]。有研究表明,叶面积指数和叶片干物质重的变化是评价弱光环境中植物光敏感性的重要指标[46],本研究也发现,套作环境中光敏感型大豆品种NDK的LAI和LDM显著的低于耐荫大豆品种ND12,因此,大豆叶片LAI和LDM可作为判断套作环境下大豆光敏感性差异的表观特征。
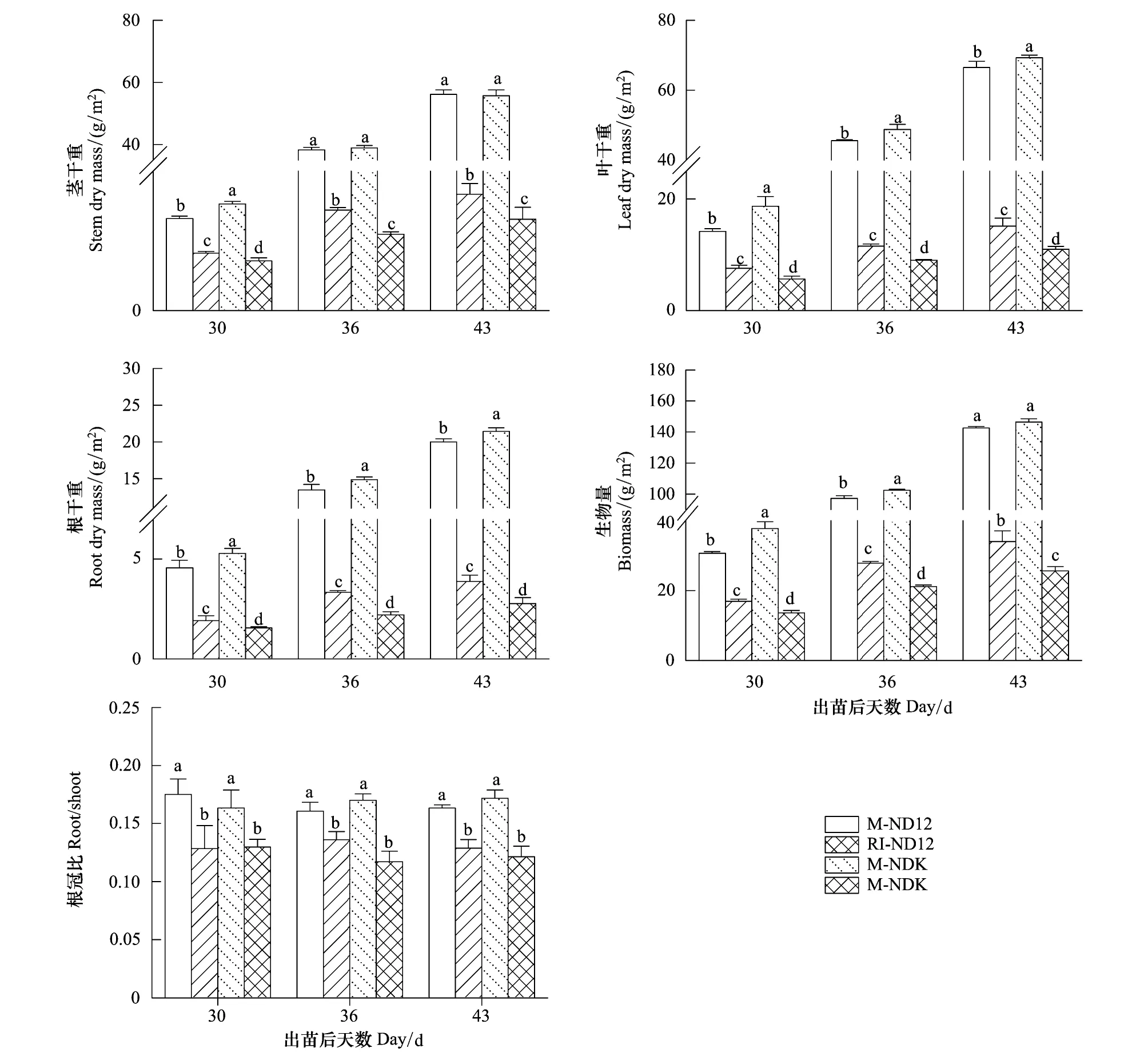
图6 大豆幼苗茎干重(SDM)、干重(LDM)、根系干重(RDM)、总生物量(BIOMASS)和根冠比(Root/Shoot)
干物质积累是植物同化和固定CO2能力的最直接表现,而弱光引起的植物干物质积累量变化的差异是直接反应物种间光敏感性的重要指标[45- 46]。套作荫蔽环境中,光敏感型大豆品种NDK的SDM、RDM和BIOMASS下降程度显著高于耐荫性品种ND12 (图6),表明NDK的生长发育和CO2固定能力受荫蔽胁迫程度更大,ND12更适合应用于玉米-大豆套作模式。有研究表明,作物95%以上的干物质来源于光合同化物,光合速率和光吸收面积是决定干物质形成的两个最重要方面[46]。本研究发现,套作环境中大豆干物质积累下降可能主要是由LAI下降导致的LDM、SDM和RDM降低所引起(表1), NDK在套作环境下较低的叶面积指数可能是导致其生物量下降程度较大的主要原因。此外,玉米遮荫导致的大豆幼苗根冠比(Root/Shoot)下降(图6)则可能意味着套作环境中碳同化物更多的被分配到地上部来最大程度地获取光照资源[47]。
表1 玉米遮荫条件下各参数变化程度间的相关性分析 (两品种的平均值), 变化程度表示为: (RI-M)/M, *代表P<0.05, **代表P<0.01
Table 1 Correlation among maize shading induced relative changes in gas exchange parameters, chlorophyll a fluorescence parameters and biomass parameters in two soybean varieties(means of two soybean varieties), Changes are expressed as percent differences between RI plants and M plants, (RI-M)/M, before Pearson’s correlation test. Asterisks denote significant correlation, * <0.05, ** <0.01.

指标Index叶绿素bChlb总叶绿素Chls叶片氮TotalNitrogeninleaf叶片碳TotalCarboninleaf叶面积指数Leafareaindex光合速率Pn气孔导度Gs胞间CO2Ci蒸腾速率TrPSⅡ光化学效率Fv/Fm茎干重Stemdrymass叶干重Leafdrymass根干重Rootdrymass生物量Biomass根冠比Root/Shoot叶绿素bChlb1总叶绿素Chls0.2561叶片氮TotalNitrogeninleaf0.110.3181叶片碳TotalNitrogeninleaf0.3820.519*0.1741叶面积指数Leafareaindex0.4490.623**0.661**0.3531光合速率Pn-0.0180.225-0.0080.764**0.1081气孔导度Gs-0.181-0.0190.0530.4790.1320.654**1胞间CO2Ci0.2490.598**0.3480.120.608*-0.3-0.0791蒸腾速率Tr-0.063-0.1340.1760.2960.2330.4520.783**-0.1831PSⅡ光化学效率Fv/Fm0.190.140.1830.1390.2290.1840.149-0.2320.4481茎干重Stemdrymass0.4190.645**0.4380.070.655**-0.362-0.5150.413-0.501*0.0411叶干重Leafdrymass0.4370.691**0.536*0.2070.797**-0.19-0.3620.414-0.3420.0960.963**1根干重Rootdrymass0.4380.699**0.4670.230.688**-0.228-0.410.426-0.4610.0210.983**0.969**1生物量Biomass0.4470.676**0.4780.1640.733**-0.265-0.4310.425-0.4240.060.987**0.991**0.987**1根冠比Root/Shoot-0.1550.542*-0.1330.323-0.0490.1260.1680.318-0.181-0.1350.1610.0770.2270.1141
4 结论
玉米-大豆套作模式中高位作物(玉米)遮荫会引起低位作物(大豆)幼苗光合作用显著下降,具体表现为:叶片Pn、Gs、Tr、LAI、TOC、SDM、LDM、RDM和BIOMASS显著下降;而与光吸收相关的叶片光合色素Chla、Chlb、Chls以及TON含量却显著上升。光合速率Pn的下调主要是由气孔限制和CO2同化过程中的能量不足共同引起,而干物质积累的降低则主要是由LAI降低所引起。LAI、SDM、LDM、RDM和BIOMASS可作为生产和育种方面评价大豆品种耐荫性差异的良好指标。耐荫性品种ND12更适合应用于西南地区玉米-大豆套作模式中,深入研究其遗传和生理生化特性对选育更适合于玉米-大豆套作模式的大豆品种具有重要的意义。
致谢:感谢许宏博士对本文撰写的帮助。
[1] Li L, Li S M, Sun J H, Zhou L L, Bao X G, Zhang H G, Zhang F S. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(27): 11192- 11196.
[2] Lithourgidis A S, Dordas C A, Damalas C A, Vlachostergios D N. Annual intercrops: an alternative pathway for sustainable agriculture. Australian Journal of Crop Science, 2011, 5(4): 396- 410.
[3] Agegnehu G, Ghizaw A, Sinebo W. Yield potential and land-use efficiency of wheat and faba bean mixed intercropping. Agronomy for Sustain Development, 2008, 28(2): 257- 263.
[4] Metwally A A, Shafik M M, EI-Habbak K E, Abdel-Wahab S I. Yield and land equivalent ratio of intercropped soybean with maize under different intercropping patterns and high population densities. Egyptian Journal of Agronomy, 2009, 31(2): 199- 222.
[5] Pypers P, Sanginga J M, Kasereka B, Walangululu M, Vanlauwe B. Increased productivity through integrated soil fertility management in cassava-legume intercropping systems in the highlands of Sud-Kivu, DR Congo. Field Crops Research, 2011, 120(1): 76- 85.
[6] Horwith B. A role for intercropping in modern agriculture. BioScience, 1985, 35(5): 286- 291.
[7] Leihner D E, Ruppenthal M, Hilger T H, Castillo J A F. Soil conservation effectiveness and crop productivity of forage legume intercropping, contour grass barriers and contour ridging in cassava on Andean Hillsides. Experimental Agriculture, 1996, 32(3): 327- 338.
[8] Fustec J, Lesuffleur F, Mahieu S, Cliquet J B. Nitrogen rhizodeposition of legumes // Sustainable Agriculture Volume 2. Netherlands: Springer, 2010: 869- 881.
[9] Zhu Y Y, Chen H R, Fan J H, Wang Y Y, Li Y, Chen J B, Fang J X, Yang S S, Hu L P, Leung H, Mew T W, Teng P S, Wang Z H, Mundt C C. Genetic diversity and disease control in rice. Nature, 2000, 406(6797): 718- 722.
[10] Saucke H, Ackermann K. Weed suppression in mixed cropped grain peas and false flax (Camelinasativa). Weed Research, 2006, 46(6): 453- 461.
[11] Shennan C. Biotic interactions, ecological knowledge and agriculture. Philosophical Transactions of the Royal Society B: Biological Sciences, 2008, 363(1492): 717- 739.
[12] Power A G. Ecosystem services and agriculture: tradeoffs and synergies. Philosophical Transactions of the Royal Society B: Biological Sciences, 2010, 365(1554): 2959- 2971.
[13] Tilman D, Cassman K G, Matson P A, Naylor R, Polasky S. Agricultural sustainability and intensive production practices. Nature, 2002, 418(6898): 671- 677.
[14] Rusinamhodzi L, Corbeels M C, Nyamangara J, Giller K E. Maize-grain legume intercropping is an attractive option for ecological intensification that reduces climatic risk for smallholder farmers in central Mozambique. Field Crops Research, 2012, 136: 12- 22.
[15] Amossé C, Jeuffroy M H, David C. Relay intercropping of legume cover crops in organic winter wheat: Effects on performance and resource availability. Field Crops Research, 2013, 145: 78- 87.
[16] 杨文钰, 杨峰, 雍太文. 我国间套作大豆研究方向和发展对策研讨会纪要. 大豆科技, 2011, (1): 35- 36.
[17] Li L, Sun J H, Zhang F S, Li X L, Rengel Z, Yang S C. Wheat/maize or wheat/soybean strip intercropping: II. Recovery or compensation of maize and soybean after wheat harvesting. Field Crops Research, 2001, 71(3): 173- 181.
[18] Reynolds P E, Simpson J A, Thevathasan N V, Gordon A M. Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada. Ecological Engineering, 2007, 29(4): 362- 371.
[19] Zhang L, Van der Werf W, Bastiaans L, Zhang S, Li B, Spiertz J H J. Light interception and utilization in relay intercrops of wheat and cotton. Field Crops Research, 2008, 107(1): 29- 42.
[20] Peng X B, Zhang Y Y, Cai J, Jiang Z M, Zhang S X. Photosynthesis, growth and yield of soybean and maize in a tree-based agroforestry intercropping system on the Loess Plateau. Agroforestry Systems, 2009, 76(3): 569- 577.
[21] 黄承建, 赵思毅, 王龙昌, 王季春, 赵勇, 蔡叶茂, 滕艳, 杨国才. 马铃薯/玉米套作对马铃薯品种光合特性及产量的影响. 作物学报, 2013, 39(2): 330- 342.
[22] Huang D, Wu L, Chen J R, Dong L. Morphological plasticity, photosynthesis and chlorophyll fluorescence ofAthyriumpachyphlebiumat different shade levels. Photosynthetica, 2011, 49(4): 611- 618.
[23] Dai Y J, Shen Z G, Liu Y, Wang L L, Hannaway D, Lu H F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content ofTetrastigmahemsleyanumDiels et Gilg. Environmental and Experimental Botany, 2009, 65(2/3): 177- 182.
[24] Makoi J H J R, Chimphango S B M, Dakora F D. Photosynthesis, water-use efficiency and δ13C of five cowpea genotypes grown in mixed culture and at different densities with sorghum. Photosynthetica, 2010, 48(1): 143- 155.
[25] 刘悦秋, 孙向阳, 王勇, 刘音. 遮荫对异株荨麻光合特性和荧光参数的影响. 生态学报, 2007, 27(8): 3457- 3464.
[26] 牟会荣, 姜东, 戴廷波, 荆奇, 曹卫星. 遮荫对小麦旗叶光合及叶绿素荧光特性的影响. 中国农业科学, 2008, 41(2): 599- 606.
[27] 眭晓蕾, 毛胜利, 王立浩, 张宝玺, 张振贤. 弱光条件下辣椒幼苗叶片的气体交换和叶绿素荧光特性. 园艺学报, 2007, 34(3): 615- 622.
[28] 王竹, 杨文钰, 吴其林. 玉/豆套作荫蔽对大豆光合特性与产量的影响. 作物学报, 2007, 33(9): 1052- 1057.
[29] 宋艳霞, 杨文钰, 李卓玺, 于晓波, 郭凯, 向达兵. 不同大豆品种幼苗叶片光合及叶绿素荧光特性对套作遮荫的响应. 中国油料作物学报, 2009, 31(4): 474- 479.
[30] 刘卫国, 蒋涛, 佘跃辉, 杨峰, 杨文钰. 大豆苗期茎秆对荫蔽胁迫响应的生理机制初探. 中国油料作物学报, 2011, 33(2): 141- 146.
[31] Arnon D I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase inBetaVulgaris. Plant Physiology, 1949, 24(1): 1- 15.
[32] 张行峰. 实用农化分析. 北京: 化学工业出版社, 2005: 191- 191.
[33] 邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2000: 121- 122.
[34] Rice S A, Bazzaz F A. Growth consequences of plasticity of plant traits in response to light conditions. Oecologia, 1989, 78(4): 508- 512.
[35] Pastur G M, Lencinas M V, Peri P L, Arena M. Photosynthetic plasticity ofNothofaguspumilioseedlings to light intensity and soil moisture. Forest Ecology and Management, 2007, 243(2/3): 274- 282.
[36] 姚允聪, 王绍辉, 孔云. 弱光条件下桃叶片结构及光合特性与叶绿体超微结构变化. 中国农业科学, 2007, 40(4): 855- 863.
[37] Franck N, Vaast P. Limitation of coffee leaf photosynthesis by stomatal conductance and light availability under different shade levels. Trees, 2009, 23(4): 761- 769.
[38] Pengelly J J L, Sirault X R R, Tazoe Y, Evans J R, Furbank R T, von Caemmerer S. Growth of the C4dicotFlaveriabidentis: photosynthetic acclimation to low light through shifts in leaf anatomy and biochemistry. Journal of Experimental Botany, 2010, 61(14): 4109- 4122.
[39] 贾士芳, 李从锋, 董树亭, 张吉旺. 弱光胁迫影响夏玉米光合效率的生理机制初探. 植物生态学报, 2010, 34(12): 1439- 1447.
[40] Buykey K O, Wells R. Response of soybean photosynthesis and chloroplast membrane function to canopy development and mutual shading. Plant Physiology, 1991, 97(1): 245- 252.
[41] Zhang Y J, Xie Z K, Wang Y J, Su P X, An L P, Gao H. Light intensity affects dry matter, photosynthesis and chlorophyll fluorescence of oriental lily. The Philippine Agriculture Scientist, 2011, 94(3): 232- 238.
[42] 周治国, 孟亚利, 施培. 苗期遮荫对棉苗茎叶结构及功能叶光合性能的影响. 中国农业科学, 2001, 34(5): 519- 525.
[43] Valladares F, Gianoli E, Gómez J M. Ecological limits to plant phenotypic plasticity. New Phytologist, 2007, 176(4): 749- 763.
[44] 任万军, 杨文钰, 张国珍, 朱霞, 樊高琼, 徐精文. 弱光对杂交稻氮素积累、分配与子粒蛋白质含量的影响. 植物营养与肥料, 2003, 9(3): 288- 293.
[45] Kennedy S, Black K, O′Reilly C, DhubháinN. The impact of shade on morphology, growth and biomass allocation inPiceasitchensis,Larix×eurolepisandThujaplicata. New Forest, 2007, 33(2): 139- 153.
[46] Niinemets Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecological Research, 2010, 25(4): 693- 714.
[47] Schall P, Lödige C, Beck M, Ammer C. Biomass allocation to roots and shoots is more sensitive to shade and drought in European beech than in Norway spruce seedlings. Forest Ecology and Management, 2012, 266: 246- 253.
Photosynthetic responses of soybean (Glycinemax) seedlings to shading caused by maize in an intercropping system
SU Benying1, SONG Yanxia2, CHEN Shengbin3, YANG Wenyu1,*
1KeyLaboratoryofCropEcophysiologyandFarmingSysteminSouthwestChina,MinistryofP.R.China,CollegeofAgriculture,SichuanAgricultureUniversity,Chengdu611130,China2AgriculturalTechnologyPromotionCenterofAnyue,Ziyang642350,China3NanjingInstituteofEnvironmentalSciences,MinistryofEnvironmentalProtection,Nanjing210042,China
Maize-soybean relay intercropping pattern, with highly economic, ecological and environmental significance, is widely applied in southwestern China. However, growth of soybean seedlings may be depressed by maize shade in this cropping pattern. The objective of this paper was to study the photosynthetic responses of soybean seedlings to maize shading in relay intercropping system in terms of morphological plasticity and photosynthetic parameters compared with mono-cropping system. A field experiment was conducted to compare gas exchange, chlorophyll fluorescence parameters, pigments contents, leaf carbon and nitrogen contents, leaf area index and dry matter accumulation of two soybean cultivars in two cropping patterns, i.e., soybean mono-cropping and maize-soybean relay intercropping. Compared with the mono-cropped seedlings, soybean seedlings intercropped with maize had significantly lower net photosynthetic rate (Pn)(-37.9%), transpiration rate (Tr) (-42.4%) and stomatal conductance (Gs) (-54.2%) due to low available light, but there was non-significant difference in two soybean varieties. The intercellular carbon dioxide concentration (Ci) andFv/Fmwere not changed significantly, in comparison with the seedlings under mono-cropping. Additionally, reductions in photosynthetic rate induced by maize shading was correlated significantly with stomatal conductance. The maize shading-induced depressions of photosynthetic rate may be attributed to stomatal limitation and energy deficiency in CO2assimilation process. Leaf carbon contents, leaf weight, leaf area index, root dry weight, total biomass and root/shoot ratio was also reduced because of maize shading, and significant difference was found between the two soybean varieties, which indicated that Nangdou12 might be a better cultivar to be relay intercropped with maize in Southwest China. The correlation analysis indicated that the relative reduction in biomass was caused by the decline in leaf area index rather thanPn. Although automatic regulation of light-harvesting efficiency in terms of total nitrogen contents, total chlorophyll contents and chlorophyll b contents was significantly improved in soybean seedlings in relay intercropping condition, it could not compensate the decline of light interception caused by the reduced light interception area. Therefore, the growth parameters may be more important index for assessing the adaptation of soybean cultivars to low light condition in intercropping system than ecophysiological parameters merely at unit leaf level.
intercropping; shading; gas exchange; pigments contents; dry matter accumulation
国家自然科学基金(31171476, 31201169); 科技部973项目(2011CB100402); 农业部公益性行业科研专项(201203096)
2013- 07- 03;
2014- 05- 30
10.5846/stxb201307031833
*通讯作者Corresponding author.E-mail: wenyu.yang@263.net
苏本营, 宋艳霞, 陈圣宾, 杨文钰.大豆幼苗对套作玉米遮荫环境的光合生理生态响应.生态学报,2015,35(10):3298- 3308.
Su B Y, Song Y X, Chen S B, Yang W Y.Photosynthetic responses of soybean (Glycinemax) seedlings to shading caused by maize in an intercropping system.Acta Ecologica Sinica,2015,35(10):3298- 3308.
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
阅读(科学探秘)(2020年8期)2020-11-06
四川蚕业(2020年3期)2020-07-16
新疆农业科学(2020年1期)2020-02-14
作文新天地(2019年25期)2019-11-27
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
新疆农垦科技(2016年2期)2016-08-21
中国麻业科学(2015年5期)2015-12-28
食品工业科技(2014年6期)2014-05-10
