
祁连山高寒草甸两型闭锁花植物块茎堇菜的繁殖分配
2015-04-08 06:50常立博张世虎
草业科学 2015年4期
郝 楠,苏 雪,吴 琼,常立博,张世虎,孙 坤
(西北师范大学生命科学学院,甘肃 兰州730070)
繁殖分配(Reproductive Allocation,RA)是植物投入到繁殖器官的资源比例[1],不仅与环境有关,还受生物因素,如个体大小、交配系统等影响[2]。植物在不同生境下的繁殖分配模式与适合度(Fitness)密切相关[3]。因此,植物的繁殖分配方式以及如何调整繁殖分配格局来适应生存环境的变化,是生态学研究的重点之一[4]。
高寒环境具有温度低、辐射强、昆虫活动频率低等特征,特殊的环境特征直接影响植物的生长和繁殖[5],植物为了生存并适应其环境在二者之间作出权衡,以保障生存和繁衍。目前,对高寒环境下植物的繁殖分配研究很多,但大都集中在营养和繁殖的权衡及性分配方面[2,6-18],对具有混合繁殖方式植物的繁殖分配及如何在不同繁殖方式间进行权衡研究相对较少。
块茎堇菜(Viola tuberifera)为分布在青藏高原及边缘地区(海拔2 800 ~3 500 m)的多年生草本,具有独特混合繁育系统。早春6 月份形成开放花但时间很短,7 月后通过地上、地下闭锁花(Cleistogamous Flowers,CL,指从不开放的花)及鳞茎进行有性和无性繁殖,为典型的两型闭锁花植物。本研究以块茎堇菜为对象,对闭锁花时期的繁殖分配进行研究,探讨以下问题:1)具有不同繁殖方式的块茎堇菜如何权衡生长和繁殖以及在不同繁殖方式上的资源投入?2)这种权衡是否受到个体大小和海拔的影响?通过本研究以期为高寒草甸植物的生活史进化和适应机制提供依据。
1 材料与方法
1.1 研究区域概况
研究区位于天祝藏族自治县境内卡洼掌高原(100°11'11. 8″ - 102°54'7. 1″ E,37°3'29. 7″ -38°8'22.0″ N),平均海拔3 050 m,年平均气温-0.2 ~4.3 ℃,≥0 ℃积温1 300 ~2 500 ℃,相对无霜期90 ~145 d,年降水量265 ~630 mm,多集中在7―9 月,年均日照时数2 500 ~2 700 h。该地区无四季之分,仅有冷暖两季[19]。土壤以亚高山草甸土、亚高山黑钙土等为主。植物生长仅靠天然降水,无地下水供给。研究区植被以退化嵩草草甸为主,主要植物有嵩草(Kobresia myosuroides)、金露梅(Potentilla fruticosa)、锦鸡儿(Caragana sinica)、翠雀(Delphinium grandiflorum)、秦艽(Gentiana macrophylla)、棘豆(Oxytropis kansuensis)、块茎堇菜(Viola tuberifera)等。
1.2 研究方法
块茎堇菜在甘肃省境内主要分布于海拔2 780 ~3 200 m 的祁连山脉退化高寒草甸区,因此2013 年8 月份闭锁花盛花期在甘肃省天祝藏族自治县卡洼掌高原,选取2 800 m(低海拔)、2 900 m(中海拔)、3 100 m (高海拔)3 个海拔梯度,设置3个样地(抓喜秀龙、抓喜双益、打柴沟),在各样地随机挖取生长良好的完整植株60 株左右(间隔大于1 m),带回实验室,除去泥土和杂质,在80 ℃烘箱中烘2 h 后用1/10 000 的电子天平对各部分进行称重。繁殖生物量为地上、地下闭锁花(果)干重;营养生物量为地上茎叶、地下根、匍匐茎、鳞茎干重;繁殖分配为繁殖生物量占总生物量的百分比;营养分配为营养生物量占总生物量的百分比;并分别计算鳞茎、地上及地下闭锁花占总生物量的百分比;总生物量反映了植株总体的资源状况和资源获取能力,不仅包括地上部分对光能获取的能力和生存竞争力,还包括地下根茎部分的营养状况和获取土壤水分、矿质元素的能力,因此本研究中个体大小以总生物量来衡量。另外,在闭锁花大量生长的采样期,前一年的无性繁殖器官鳞茎大多枯萎,数目极少。
1.3 数据处理
所有数据用SPSS 21.0 软件非参数检验中的One-Sample K-S 检验正态分布,为正态分布的用oneway ANOVA 检验居群水平上3 组数据间的差异(P=0.05);非正态则用K Independent Samples 进行比较,通过Correlate 中的Bivariate Correlations 分析两个变量之间的相关关系,用Regression 进行曲线回归分析。用Excel 2003 和SPSS 21.0 作图。
2 结果与分析
块茎堇菜居群的生物量分配格局如表1 所示。在总生物量中分配到营养器官的比例远高于繁殖器官,这可能是由于在高海拔资源有限条件下植株将更多的资源投入营养器官以保障存活。有性繁殖中,地上闭锁花分配(0.14% ~1.35%)受海拔影响变化较大(P <0.05),而地下闭锁花繁殖分配(0.75% ~0.77%)则比较稳定(P >0.05)。低海拔地上闭锁花繁殖分配高出地下闭锁花繁殖80.00%,中、高海拔下,则比地下闭锁花分别低52.63%和81.82%,可以看出,在低海拔以地上闭锁花为主要有性繁殖方式,在高海拔地区以地下闭锁花为主要有性繁殖方式。另外,在采样中发现,块茎堇菜在闭锁花繁殖盛花期仅有少数个体形成鳞茎(2 800 m 海拔下仅1 个个体具有鳞茎,2 900 m 海拔下仅2 个个体具有鳞茎,3 100 m 海拔下仅4 个个体具有鳞茎),鳞茎数目很少,其资源分配仅为0.07% ~0.12%。
2.1 繁殖分配对海拔的响应
不同海拔梯度下块茎堇菜居群的繁殖生物量表现为总生物量、繁殖生物量、地下闭锁花生物量随海拔升高而降低(表1、图1)。差异显著性分析表明,总生物量在低海拔和高海拔间差异显著(P <0.05),地上闭锁花生物量和总繁殖生物量在低海拔与中、高海拔间存在显著差异。
繁殖分配和繁殖生物量规律基本一致,但也存在微小差别。总有性繁殖分配、地上闭锁花繁殖分配随海拔升高而下降,地下闭锁花的繁殖分配则略有上升。差异显著性分析表明,总有性繁殖分配和地上闭锁花分配在低海拔和中、高海拔间差异显著(P <0.05),地下闭锁花分配在不同海拔间差异均不显著(P >0.05)。
两种闭锁花分配占总有性繁殖比例变化趋势为,地下闭锁花繁殖分配所占比重随海拔升高而明显上升(P <0.05),从35.71%升至84.62%,而地上闭锁花繁殖分配所占比例则从64. 29% 降至15.38%。另外若将繁殖分为地上(以地上闭锁花生物量计)、地下(以地下闭锁花和鳞茎生物量计)两种方式,则发现随海拔升高地下繁殖的绝对投入有所下降,但相对比例略有上升,且差异显著性分析表明,地下闭锁花繁殖分配占总有性繁殖比重随海拔
增高明显增加,鳞茎比例虽呈上升趋势,但不明显。说明高海拔寒冷环境下,块茎堇菜在营养生长与繁殖间存在权衡关系;在地上与地下有性繁殖分开配间存在权衡关系,本研究中植株选择将更多资源投入到地下繁殖器官以保障其繁殖成功。

表1 不同海拔居群分配格局及总生物量Table 1 Allocation patterns and total biomass within different populations among altitudes
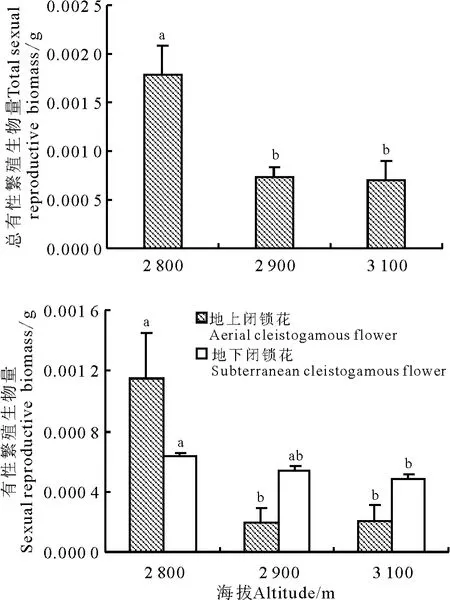
图1 不同海拔居群的繁殖投入Fig.1 Reproductive investments among different populations with different altitudes
2.2 个体大小与繁殖分配的关系
在植株的个体水平上,对其个体大小(总生物量)与繁殖分配关系的分析(图2)表明,有性繁殖分配与个体大小呈极显著负幂指数相关(r= -0.517,P=0.000),即随个体增加有性繁殖分配呈先急剧下降后缓慢持平的趋势。在有性繁殖分配中,无论是线性回归分析,还是曲线回归分析,总体上,地上闭锁花分配与个体大小均不存在明显的相关关系,因为野外观察发现,大多数个体不具有地上闭锁花,仅20 株个体存在地上闭锁花,对这部分个体地上闭锁花繁殖分配的大小依赖性分析发现,地上闭锁花繁殖分配与个体大小呈极显著负幂指数相关(r =-0.730,P=0.000)(图2);地下闭锁花繁殖分配与个体大小也呈极显著负幂指数相关(r= -0.762,P=0.000)(图2),即随着个体大小增加,地下闭锁花繁殖分配先急剧下降后趋于平缓,说明繁殖分配具个体大小依赖性,地下闭锁花的分配受个体大小制约,总体来说,地上闭锁花繁殖分配与个体大小依赖性不强,但具有地上闭锁花的20 株个体中,两者呈负幂指数相关关系。另外,由于本研究中鳞茎数目很少,无法统计分析其个体大小,有待进一步研究。
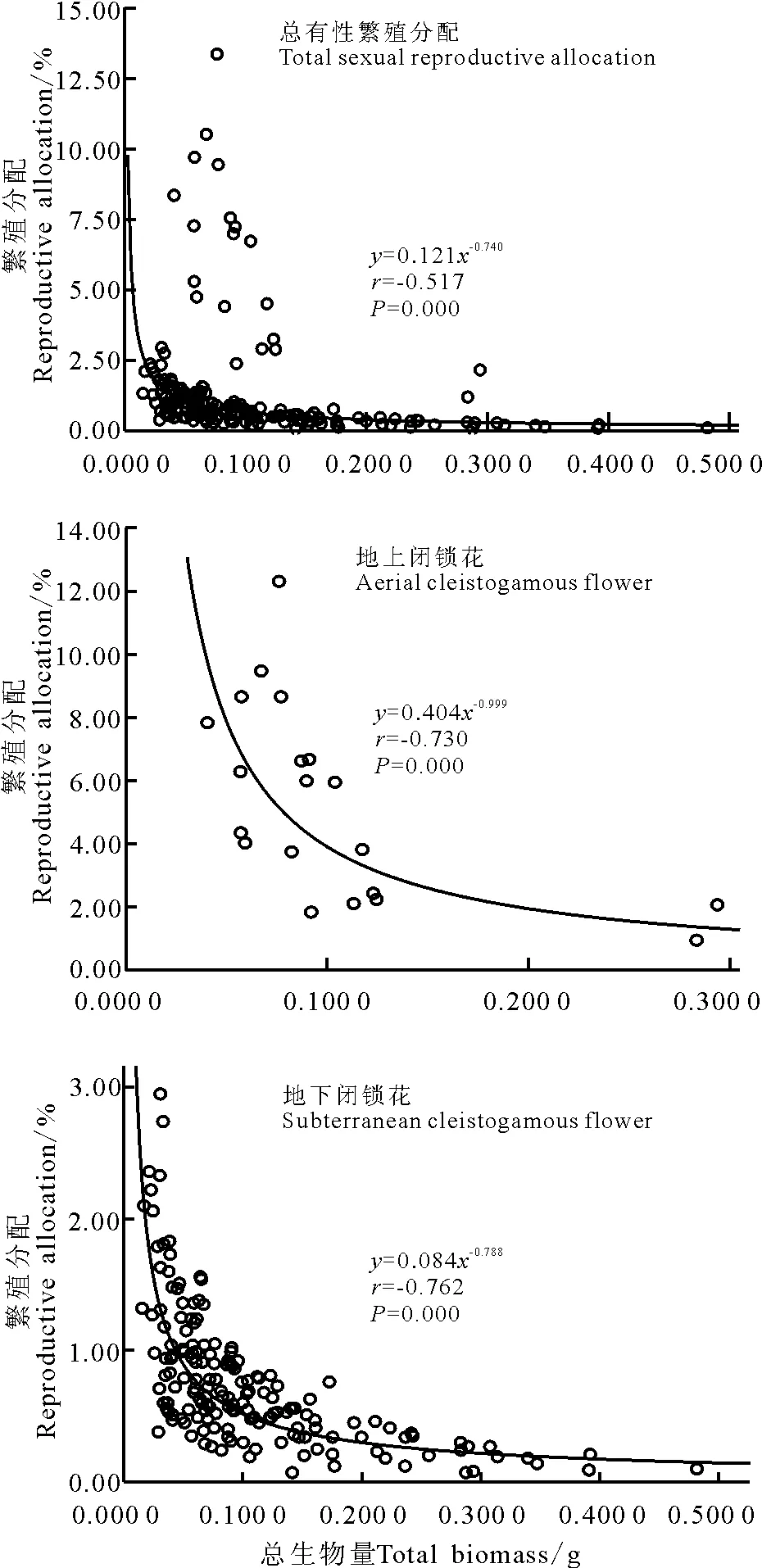
图2 个体大小与繁殖分配的关系Fig.2 The relationship between individual size and reproductive allocation
3 讨论与结论
3.1 块茎堇菜的繁殖分配
混合交配系统(Mixed-mating System)表达受外界环境变化和内部资源状态如个体大小影响[20]。关于混合交配系统维持机制的研究很多[21-26],但从繁殖分配角度研究两型闭锁花受精混合繁育系统的却很少。繁殖分配不仅是生活史进化的结果,也反映了植物对生存环境的响应[5]。在资源受限环境中,个体将自身获得的有限资源进行最优化分配以增加适合度[2]。虽然有关繁殖分配大量研究已表明,植物在有性繁殖与未来生存、营养生长或无性繁殖等功能间存在着权衡[13,27],但多年生植物这种权衡仍未得到有力证明[15]。本研究表明,块茎堇菜不仅在营养与繁殖间存在权衡关系,而且在地上闭锁花和地下闭锁花有性繁殖间存在权衡,但无性与有性繁殖的这种权衡关系在本研究中不明显。高寒环境特有的块茎堇菜在较低海拔处繁殖投入比例显著高于较高海拔的繁殖分配,这是因为高海拔环境下植物为了保障存活,优先将资源分配给营养器官维持生长,从而导致繁殖投资降低。在闭锁花繁殖盛花期,鳞茎刚刚形成,植株仍以有性繁殖为主,这与珠芽蓼(Polygonum viviparum)的研究结论相符[13,16],但本研究野外观察发现,在闭锁花繁殖盛花期以后鳞茎数目会逐渐增多。另外,块茎堇菜兼具地上闭锁花和地下闭锁花,由于地上繁殖器官易受到低温、昼夜温差、辐射等不利条件影响[9],繁殖收益不如地下闭锁花,但地上闭锁花可通过蒴果开裂弹射、蚂蚁的二次搬运等方式扩散种子,传播距离较远,与母株竞争较小[28],因此块茎堇菜表现为地上闭锁花投资变化幅度较大,低海拔(2 800 m)居群对地上闭锁花投入较多。而地下闭锁花受不确定环境因素影响较小,发育比较稳定,埋藏在土壤中的种子更能躲避干扰、疾病和捕食损害[9],但后代间及后代与母体间的竞争往往较为激烈,因此,在高海拔植株选择以地下闭锁花作为主要的有性繁殖方式,说明块茎堇菜生物量分配格局存在权衡关系,通过最佳资源配置,以其特有繁殖属性适应极端环境,维持物种生存和繁衍。值得注意的是,繁殖是植物特定时期的功能,繁殖投资存在明显季节性[13],块茎堇菜开放花在早春形成,本研究仅是对其在闭锁花盛花期繁殖分配进行研究,尚需在时间尺度上开展不同季节、不同发育阶段繁殖分配变化规律的研究。
3.2 繁殖分配对海拔和个体大小的响应
随海拔梯度升高,资源可利用率与温度降低,霜冻与辐射量上升,环境条件更加恶劣,不利于植物生长繁殖[9]。资源利用假说认为无机环境资源影响植物生长发育,环境资源充足与否决定了繁殖分配高低[13]。已有研究表明,繁殖分配随海拔变化在种间和种内存在较大的差异[13],有些多年生植物繁殖分配随海拔升高而上升[10,18,27],也有随海拔升高繁殖分配下降的例子[9,29],而岩白菜(Bergenia purpurascens)则随海拔升高呈先下降后上升趋势[8]。本研究表明,块茎堇菜繁殖分配对海拔具有一定的适应规律,即随海拔升高,总有性繁殖投入及分配、地上闭锁花繁殖投入及分配明显减少;地下闭锁花投入有所下降,但相对分配略有上升,且占总繁殖分配比重上升;鳞茎繁殖投入和分配虽变化幅度不明显,但却呈上升趋势。随海拔升高,块茎堇菜的生长条件更加恶劣,植物获取资源的能力降低,植物面临较大生存危机,为保障生存,植株选择给予更多资源于营养器官,是导致繁殖绝对资源量和相对分配减少的原因。与地下繁殖器官不同的是,植物地上部分对环境变化非常敏感,随海拔升高,环境更加严酷,使得块茎堇菜地上闭锁花繁殖投入比例降低,繁殖投入及分配降低,而地下闭锁花和鳞茎繁殖分配则保持相对稳定或有所增加。可见,块茎堇菜通过对地上和地下闭锁花之间繁殖资源投入的权衡来最大限度地保证繁殖成功,提高繁殖适合度,以适应恶劣的高海拔环境。
在许多植物中存在繁殖分配的个体大小依赖性[27-29],这种依赖性归根于内在影响,即资源获取和繁殖分配在一个植株内的生理权衡[30]。因为对一个有限资源库而言,对繁殖活动投入增大,意味着对茎叶营养活动投入减少,所以植株资源获取能力下降,影响个体存活和生长[16]。大量研究表明,高海拔地区植物的繁殖分配常与个体大小呈负相关[14,16,30-31],原因可能是高寒草甸恶劣环境条件使植物生存风险增大,繁殖代价更高,激烈的竞争使得大个体倾向于营养生长并将更多资源投入到营养器官,吸收更多的光能产生有机化合物;而小个体由于自身资源有限且竞争力较弱,将更多资源投入到了繁殖生长,在严酷的环境下保障子代延续[14,31]。本研究表明,青藏高原块茎堇菜的繁殖分配与个体大小呈极显著负幂指数相关,随个体增加,繁殖分配呈先急剧减小后缓慢持平的关系,其中繁殖分配呈急剧减小的趋势与大多数研究中所得出的负相关关系一致[3,14-16,30-31]。植株为了度过严酷、多变的环境,有性繁殖由于其自身的变异性,成为恶劣条件下的优先选择,是保障植物生存下去的必备手段。本研究中即便植株个体很小,也要进行有性繁殖,才可以在高海拔环境中存活、延续子代。因此小个体的繁殖分配仍然很高,但当繁殖分配值达到一个临界点时,便不再增加,此时植物已获得足够保障繁殖的能力,所以即便个体继续增大,其繁殖分配值也不发生变化。此外,本研究还发现,总体上块茎堇菜地上闭锁花繁殖分配对个体大小的依赖性不强,但具有地上闭锁花的少数个体仍受个体大小的制约,地下闭锁花分配也存在明显的个体大小依赖性。这可能是由于块茎堇菜在内部资源受限制的情况下,选择将资源投入到更具繁殖保障功能的闭锁花中,提高对环境的适合度。
[1] 李金花,李镇清,刘振国.不同刈牧强度对冷蒿生长与资源分配的影响[J].应用生态学报,2004,15(3):408-412.
[2] Hutchings M J.Resource allocation patterns in clonal herbs and their consequences for growth[A].Bazzaz F A,Grace J.Plant Resource Allocation[M].London:Academic Press,1997.
[3] 陈学林,梁艳,齐威,苏梅,杜国桢. 一年生龙胆属植物的繁殖分配及其花大小、数量的权衡关系研究[J]. 草业学报,2009,18(5):58-66.
[4] 苏智先,张素兰,钟章成.植物生态学进展[J].生态学杂志,1998,17(4):39-46.
[5] Körner C.高山植物功能生态学[M].吴宁,罗鹏,译.北京:科学出版社,2003.
[6] 张茜,赵成章,马小丽,侯兆疆,李钰.高寒草地狼毒居群繁殖分配对海拔的响应[J].生态学杂志,2013,32(2):247-252.
[7] 苏梅,齐威,阳敏,杜国祯.青藏高原东部大通翠雀花的花特征和繁殖分配的海拔差异[J].兰州大学学报(自然科学版),2009,45(2):61-65.
[8] 王赟,胡莉娟,段元文,杨永平.岩白菜(虎耳草科)不同海拔居群的繁殖分配[J].云南植物研究,2010,32(3):270-280.
[9] 樊宝丽,孟金柳,赵志刚,杜国祯. 海拔对青藏高原东部毛茛科植物繁殖特征和资源分配的影响[J]. 西北植物学报,2008,28(4):0805-0811.
[10] 王一峰,岳永成.青藏高原东缘不同海拔波缘风毛菊资源分配及花部特征对种子数目和质量的影响[J].植物生态学报,2014,38(4):366-374.
[11] 徐波,王金牛,石福孙,高景,吴宁.青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应[J].植物生态学报,2013,37(3):187-196.
[12] 李冰,刘左军,赵志刚,胡春,任红梅,伍国强.海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响[J].草业学报,2013,22(1):10-19.
[13] 何亚平,段元文,费世民,刘建全,杨慧玲.青藏高原天山报春高寒湿地居群的花期资源分配[J].应用与环境生物学报,2008,14(2):180-186.
[14] 陶冶,张元明.准格尔荒漠6 种类短命植物生物量分配与异速生长关系[J].草业学报,2014,23(2):38-48.
[15] 刘左军,杜国祯,陈家宽.不同生境下黄帚橐吾(Ligularia virgaurea)个体大小依赖的繁殖分配[J].植物生态学报,2002,26(1):44-50.
[16] 赵志刚,杜国祯,任青吉.5 种毛茛科植物个体大小依赖的繁殖分配和性分配[J].植物生态学报,2004,28(1):9-16.
[17] 赵玉红,魏学红,苗彦军,其美,扎旺. 藏北高寒草甸不同退化阶段植物群落特征及其繁殖分配研究[J]. 草地学报,2012,20(2):221-228.
[18] 索南措,王一峰,李梅,岳永成,李毛先.青藏高原东缘常见种长毛风毛菊的繁殖分配[J].生态学杂志,2013,32(6):1433-1438.
[19] 天祝藏族自治县县志编篡委员会.天祝县志[M].兰州:甘肃民族出版社,1994.
[20] Ranua V A,Weinig C.Mixed-mating strategies and their sensitivity to abiotic variation in Viola lanceolata L.(Violaceae)[J].The Open Ecology Journal,2010,3(1):83-94.
[21] Oakley C G,Moriuchi K S,Winn A A.The maintenance of outcrossing in predominantly selfing species:Ideas and evidence from cleistogamous species[J].Annual Review of Ecology Evolution and Systematics,2007,38:437-457.
[22] Kalisz S,Vogler D W.Benefits of autonomous selfing under unpredictable pollinator environments[J].Ecology,2003,84:2928-2942.
[23] Masuda M,Yahara T,Maki M.Evolution of floral dimorphism in a cleistogamous annual,Impatiens nolitangere L. occurring under different environmental conditons[J].Ecological Research,2004,19:571-580.
[24] Goodwillie C,Kalisz S,Eckert C G.The evolutionary enigma of mixed mating systems in plants:Occurrence,theoretical explanations,and empirical evidence[J].Annual Review of Ecology Evolution and Systematics,2005,36:47-79.
[25] Winn A A,Moriuchi K S.The maintenance of mixed mating by cleistogamy in the perennial violet Viola septemloba(Violaceae)[J].The Open Ecology Journal,2009,96(11):2074-2079.
[26] Forrest J,Thomson J D.Pollen limitation and cleistogamy in subalpine Viola praemorsa[J].Botany,2008,86:511-519.
[27] 王一峰,李梅,李世雄,郭杰,陈玉萍,王瑞雪.青藏高原东缘星状风毛菊生殖分配对海拔的响应[J]. 植物生态学报,2012,36(11):1145-1153.
[28] 巩明明,苏雪,孙坤.青藏高原特有植物块茎堇菜地上地下结果性及其散布机制[J].草业科学,2009,26(4):16-19.
[29] 赵方,杨永平.中华山蓼不同海拔居群的繁殖分配研究[J].植物分类学报,2008,46(6):830-835.
[30] Pickering C M.Size-denpendent reproduction in Australian alpine Ranunculus[J].Australian Journal of Botany,1994,76:43-50.
[31] Reekie E G. An explanation for size-dependent reproductive allocation in Plantago major[J]. Canadian Journal of Botany,1998,76:43-50.
猜你喜欢
新农村(浙江)(2022年8期)2022-08-01
现代苏州(2022年5期)2022-03-21
老年博览·上半月(2021年12期)2021-12-21
今日农业(2021年15期)2021-11-26
花卉(2021年15期)2021-08-17
种子(2019年7期)2019-08-13
现代园艺(2017年19期)2018-01-19
奇闻怪事(2016年11期)2017-02-14
IT时代周刊(2015年9期)2015-11-11
云南中医学院学报(2015年2期)2015-07-31
