
受体型蛋白酪氨酸磷酸酶Q的研究进展*
2015-05-16 00:51黄永红史慧棉周晓燕罗志军徐方云南昌大学医学院病理生理学教研室生理教研室江西南昌330006
中国病理生理杂志 2015年7期
黄永红,史慧棉,周晓燕,徐 宏,罗志军△,徐方云△(南昌大学医学院病理生理学教研室,生理教研室,江西南昌330006)
Advance in receptor type protein tyrosine phosphatase Q
HUANG Yong-hong1,SHI Hui-mian1,ZHOU Xiao-yan1,XU Hong2,LUO Zhi-jun1,XU Fang-yun1(1Department of Pathophysiology,2Department of Physiology,Medical College of Nanchang University,Nanchang 330006,China.E-mail: zluo559914@ gmail.com; scb9999@ sina.com)
受体型蛋白酪氨酸磷酸酶Q的研究进展*
黄永红1,史慧棉1,周晓燕1,徐宏2,罗志军1△,徐方云1△
(南昌大学医学院1病理生理学教研室,2生理教研室,江西南昌330006)
研究表明,从基础代谢到细胞生长、分化、增殖等几乎每一细胞事件都与蛋白质酪氨酸磷酸化过程密切相关[1-4]。这一过程分别受蛋白酪氨酸激酶( protein tyrosine kinases,PTKs)和蛋白酪氨酸磷酸酶( protein tyrosine phosphatases,PTPs)共同调控。PTKs能特异地使蛋白质上的酪氨酸残基磷酸化,而PTPs能使磷酸化的酪氨酸残基去磷酸化,两者相互协调,共同调节蛋白质的磷酸化水平(图1)。一旦PTPs 与PTKs之间正常的平衡被打破,必将引起蛋白质酪氨酸磷酸化的异常,该异常可导致包括癌症在内的多种疾病发生。例如,表皮生长因子之类的受体酪氨酸蛋白激酶( receptor tyrosine kinases,RTKs)的激活和过量表达[5]、非RTKs(如Src、Bcr-Abl)的激活和过表达[6-7]都是细胞癌变的重要发病机制。
受体型蛋白酪氨酸磷酸酶Q( receptor type PTP Q,PTPRQ)是III型受体型PTPs家族的成员之一[8]。近年研究发现,PTPRQ蛋白具有多种生物学功能,与某些疾病的发生密切相关。本文将从PTPRQ基因的结构、功能及其与疾病的关系等方面进行综述,以深入理解PTPRQ在疾病中的潜在作用。
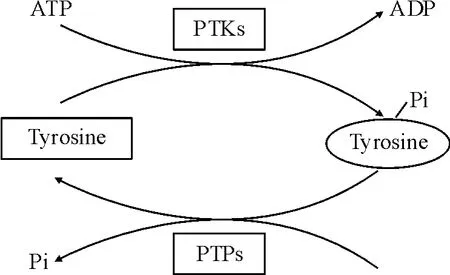
Figure 1.Reversible protein tyrosine phosphorylation.图1 可逆的蛋白酪氨酸磷酸化
1 PTPRQ基因的定位与结构
PTPRQ基因最初由Wright等[9]研究大鼠系膜增生性肾小球肾炎PTPs基因表达时发现并将其克隆。由于其在活化的系膜细胞中表达明显上调,故当时称为肾小球系膜细胞受体样PTP( receptor-like protein tyrosine phosphatase expressed by glomerular mesangial cells,rPTP-GMC1)。研究发现,鼠类rPTPGMC1基因定位于10号染色体的远端,其在染色体的位点与人类PTPRQ基因高度相似[9]。现已证实,人类PTPRQ基因定位于12q21.2,包含多个mRNA变构体,其中2个主要表达产物分别由全长PTPRQ( full length-PTPRQ,PTPRQ-FL)和截短型PTPRQ ( truncated-PTPRQ,PTPRQt) mRNA编码[10]。PTPRQ-FL mRNA全长8 066 bp,有45个外显子,编码由2 299个氨基酸组成的蛋白质,相对分子量约为255 kD。与其它III型受体PTPs类似,PTPRQ-FL蛋白自氨基端到羧基端分别由疏水性信号肽、由18个III型纤维连接蛋白( fibronectin typeⅢ,FNIII)样重复序列组成的胞外结构域、跨膜结构域和PTPs胞内催化结构域共4个结构域组成(图2)[9]。PTPRQ催化结构域的一级结构与大多已知的PTPs高度相似,所不同的是PTPs高度保守的由色氨酸( W)、脯氨酸( P)和天冬氨酸( D)组成的基序( WPD基序)中的天冬氨酸( D)被谷氨酸( E)代替了,这一点与另外3个典型PTPs成员包括PTPRU、PTPD1和HDPTP一样[11]。PTPRQt的mRNA是PTPRQ的一种组织特异性剪接形式,它编码一个只含催化结构域的胞浆蛋白质[10]。

Figure 2.Primary structure of PTPRQ protein.SP: signal peptide; TM: transmembrane.图2 PTPRQ蛋白的一级结构
最近Yu等[11]揭示了PTPRQ催化结构域的晶体结构。PTPRQ催化结构域的拓扑结构由中央1个扭曲的8链β折叠和两侧6个α螺旋组成,这种结构与PTPδ的催化结构域相似。PTPRQ催化结构域的核心由1个PTP环和1个WPE环组成。此外,靠近PTPRQ催化核心的M6环序列被打乱并形成了一侧更平的催化口袋。
2 PTPRQ的功能
目前PTPRQ的功能仍未完全清楚。现有的研究表明,PTPRQ是一种具有磷脂酰肌醇磷酸酶( phosphatidylinositol phosphatases,PIPs)和PTPs双重磷酸酶活性的受体型PTPs。它既可以通过其PIPs的催化活性调控相关信号通路,也可以通过PTPs的催化活性调控相关信号通路,进而在细胞增殖与分化、迁移等多种细胞事件中发挥作用。
2.1PTPRQ的磷酸酶活性如前所述,在PTPRQ蛋白分子结构中,不仅PTPs高度保守的WPD基序中的天冬氨酸( D)被谷氨酸( E)代替,而且靠近PTPRQ催化核心的M6环序列被打乱并形成了一侧更平的催化口袋,这使得PTPRQ对磷酸酪氨酸底物的催化活性较弱,而对磷脂酰肌醇( phosphatidylinositol,PI)底物表现出了较强的催化活性。PTPRQ这种特殊的磷酸酶活性在许多功能性实验中已得到证实。体外过表达的PTPRQ催化结构域具有广泛的PIPs活性,特别是优先对PI-3,4,5-三磷酸( PI-3,4,5-triphosphate,PIP3)的去磷酸化[8]。Yu等[11]将PTPRQ WPE基序2 847位的酸性氨基酸谷氨酸( E)突变为中性氨基酸丙氨酸( A,E2847A)或另一种酸性氨基酸谷氨酰胺( Q,E2847Q)后,发现E2847Q突变型PTPRQ对PI底物的催化活性与野生型PTPRQ相当,而E2847A突变型PTPRQ对PI底物的催化活性明显强于野生型PTPRQ,表明PTPRQ WPE基序中酸性氨基酸并非其催化PI底物所必需。Yu等[11]进一步将PTPRQ中M环中2 785位的谷氨酸( E)突变为丙氨酸( A,E2785A),结果发现双重突变型( E2785A/E2847A) PTPRQ和双重突变型( E2785A/ E2847Q) PTPRQ对PI底物的催化活性明显降低,表明PTPRQ M环中一侧变平的催化口袋在靶向PI底物的催化活性方面非常重要。但另有研究发现[10],当PTPRQ中WPE基序突变为WPD基序后PTPRQ重新获得PTPs的催化活性而丧失其PIPs的催化活性,表明WPE基序亦对PTPRQ的PIPs活性非常关键。
虽然上述研究表明PTPRQ具有PIPs和PTPs双重磷酸酶活性,但进一步研究发现PTPRQ的磷酸酶活性可因PTPRQ亚细胞定位的特定模式不同而不同,或靶向PI底物,或靶向磷酸酪氨酸底物,或不具有特异性限制。例如,在活化的大鼠系膜细胞中PTPRQ蛋白主要定位于细胞胞浆,其催化活性并不具有特异性限制[10]。体外过表达胞内催化结构域的PTPRQ具有较弱的PTPs活性[8]。而在人肾小囊脏层上皮细胞即足细胞中PTPRQ蛋白以跨膜蛋白形式定位于基底细胞膜,其特异性地靶向PI底物[8]。此外,在内耳的感光毛细胞中PTPRQ也是以跨膜蛋白形式存在于其顶膜,特异性地靶向PI底物[12]。这些研究表明PTPRQ蛋白表达具有跨膜和胞浆2种调节性表达模式,不同的表达模式决定了磷酸酶活性的不同。现已证实,PTPRQ蛋白表达模式的不同源于PTPRQ基因转录启动子和剪接方式的不同。5’端启动子启动受体样跨膜PTPRQ蛋白的表达,而位于跨膜与近膜结构域之间的启动子则启动只含催化结构域的胞浆PTPRQ蛋白表达[10]。然而,如何调节PTPRQ基因这种启动子和剪接方式的细胞与种属特异性来获得PIPs和PTPs活性,这一机制尚不清楚。
2.2PTPRQ的生物学功能研究表明,PTPRQ与细胞的增殖、凋亡、分化与迁移等多种生物学行为有关。Wright等[9]研究发现,在大鼠实验性系膜增生性肾小球肾炎模型中,系膜细胞损伤后出现系膜细胞迁移与增殖即系膜细胞活化时伴随PTPRQ mRNA表达的明显上调;而且,当系膜细胞迁移与增殖期结束后伴随PTPRQ表达的消失;这些研究提示PTPRQ具有调节肾小球损伤反应过程中系膜细胞增殖与迁移的作用。另外,从遗传性角度来看,果蝇的受体型PTPs如DLAR[13]、DPTP10D[14]和DPTP69D[15]等是重要的细胞迁移与轴突导向调节子也印证了PTPRQ具有调节细胞迁移的作用。Oganesian等[8]研究发现,体外过表达PTPRQ胞内催化结构域能抑制人神经胶质瘤U87MG和U373MG细胞的增殖,诱导细胞G0/G1阻滞,促进细胞的凋亡,同时发现PTPRQ的这种调控细胞生长的作用主要是凭借其PIPs活性来降低细胞内PIP3的水平,进而负性调控蛋白激酶B ( protein kinase B,PKB/Akt)信号通路,而并非通过抑制PI3K的结合和活化来影响PKB/Akt信号通路,这一点与肿瘤抑制因子第10号染色体上缺失的与张力蛋白同源的磷酸酶基因( phosphatase and tensin homologue deleted on chromosome 10,PTEN)极其相似。Jung等[16]在研究PTPs介导人骨髓源性间充质干细胞( mesenchymal stem cells,MSCs)分化为脂肪细胞的过程中发现,PTPRQ的mRNA表达在该过程中出现明显下调;进一步研究发现,体外过表达PTPRQ胞内催化结构域可抑制MSCs分化为成熟的脂肪细胞,而且也证实PTPRQ通过其PIPs活性调节细胞内PIP3的水平而影响PKB/Akt信号通路,进而调控MSCs分化为脂肪细胞。
此外,研究显示,PTPRQ在内耳的发育和毛细胞的分化成熟过程中发挥重要作用。Goodyear等[12]研究发现,胚胎13.5 d时PTPRQ首先表达于小鼠前庭的毛细胞,胚胎17.5 d时表达于耳蜗基底的毛细胞,而且PTPRQ的表达伴随至前庭和耳蜗毛细胞分化成熟的整个过程;在PTPRQ敲除转基因小鼠模型中研究发现,突变的前庭毛细胞缺乏轴连接器,且PTPRQ功能性缺失可导致前庭毛细胞小鼠耳蜗基底部毛细胞进行性缺失,继而影响小鼠的高频听力。这些研究表明,PTPRQ是耳蜗毛细胞发育成熟、轴连接器形成和高频听毛细胞长时存活所必须的PTPs。此外,Nayak等[17]研究发现至少有3种不同PTPRQ亚型表达于鸡内耳毛细胞。其中,2种亚型表达于发育成熟的鸡内耳毛细胞,1种亚型表达于未发育成熟的鸡内耳毛细胞。
3 PTPRQ与疾病
3.1PTPRQ与失聪如前所述,PTPRQ在内耳的发育和毛细胞的分化成熟过程中发挥重要作用。如果PTPRQ基因突变或蛋白表达异常可能影响内耳的发育和毛细胞的分化成熟,从而导致听力相关性疾病的发生。目前已有多项研究表明,PTPRQ基因突变与人类听力疾病密切相关[18-21]。Shahin等[18]首次报道了PTPRQ突变与人类遗传性失聪密切相关。该研究团队对有血缘关系的失听患者和听力正常的亲属DFNB84基因座染色体12q21 12.5 Mb纯合子区域的失聪候选基因测序时发现,所有失聪患者均存在PTPRQ基因纯合子无义突变( c.1285C→T),进而导致翻译在429位密码子处提前终止,而类似情况并未见于听力正常的亲属。Schraders等[20]报道,常染色体隐性非综合征性失聪患者DFNB84基因座存在PTPRQ纯合子突变,序列分析研究发现在非遗传性的患者中存在PTPRQ基因的无义突变,而在遗传性患者中的存在PTPRQ基因的错义突变,而且发现错义突变位点位于PTPRQ FNIII结构域,无义突变产生了不含跨膜部分和PTP结构域而仅含小部分FNIII结构域的截短PTPRQ蛋白。最近,复旦大学Sang等[21]在常染色体隐性非综合征性失聪患者DFNB84基因家族中鉴定了新的PTPRQ杂合子突变,该杂合子突变引起了移码突变并产生了截短PTPRQ蛋白。这些研究均表明,PTPRQ基因极有可能是一个新的失聪相关性基因,但PTPRQ通过何种机制导致患者失聪还需深入研究。
3.2PTPRQ与肿瘤肿瘤被普遍认为是由多个控制细胞增殖和生存的基因突变所致,这些原癌基因突变成永久激活的癌基因;或抑癌基因发生失活突变,致使其编码的蛋白产物失去活性。这些突变的综合效应导致细胞的生长失去控制,形成癌变的细胞。许多研究表明PTPs与PTKs协调调控蛋白的磷酸化水平,在细胞增殖、分化及凋亡等多种生命活动过程中发挥作用。现有研究显示PTPs在肿瘤发生中扮演着双重角色,它们中的一些成员具有抑制细胞生长的功能,其失活突变可以导致细胞内酪氨酸磷酸化的增加,使控制细胞增殖的信号转导通道持续性激活,从而使细胞生长失控,最终可以导致癌变。例如,PTEN基因异常广泛存在于前列腺癌、肺癌、胃癌和乳腺癌等多种人类恶性肿瘤中,负性调控PKB/Akt等信号通路而抑制肿瘤细胞的生长和侵袭转移[22-23]。然而,一些PTPs可以是癌基因产物,其活性增强可促进细胞增殖而导致癌变。例如,PTPN11基因编码的非受体型PTP蛋白Src同源酪氨酸磷酸酶2( Src homology phosphotyrosyl phosphatase 2,SHP2)在白血病[24]、乳腺癌[25]等肿瘤中存在异常活化而被认为是原癌基因,其可活化生存相关信号通路如有丝分裂原活化蛋白激酶( mitogen-activated protein kinase,MAPK)通路。
现有研究显示,PTPRQ与肿瘤之间有着一定的联系。如前所述,体外过表达PTPRQ胞内催化结构域能抑制人神经胶质瘤U87MG和U373MG细胞的增殖,诱导细胞G0/G1阻滞,促进细胞的凋亡[8]。Laczmanska等[26]研究发现,散发性结直肠癌患者癌组织中存在PTPRQ基因的异常扩增,提示PTPRQ在散发性结直肠癌可能是起着癌基因的作用。而人类蛋白质表达图集( The Human Protein Atlas)的资料显示,在肺癌、结肠癌、前列腺癌和乳腺癌等多种类型的肿瘤组织存在PTPRQ的异常改变。这提示PTPRQ在这些肿瘤中可能起着某种作用。如前所述,PTPs家族成员可以是癌症促进因子,也可以肿瘤抑制因子。那么,PTPRQ在肿瘤中到底是癌症促进因子还是肿瘤抑制因子,亦或二者兼而有之,在不同的特定环境下发挥不同的作用?这些问题的阐明对于理解PTPRQ在疾病中尤其是肿瘤发病中的作用具有极其重要的意义。
3.3PTPRQ与其它疾病动物实验中显示PTPRQ可通过促进系膜细胞增殖与迁移而参与系膜增生性肾小球肾炎的发生[9]。但目前尚缺乏PTPRQ与人类肾小球疾病之间关系的直接证据。体外研究[16]发现,人MSCs分化为脂肪细胞的过程中PTPRQ mRNA表达在该过程中出现明显下调,且过表达PTPRQ胞内催化结构域可抑制MSCs分化为成熟的脂肪细胞。该研究提示PTPRQ可能参与肥胖症的发生。
4 存在的问题与展望
综上所述,PTPRQ是作为受体型PTPs家族成员之一,其编码的蛋白质具有PIPs和PTPs双重特异性磷酸酶作用,尤其是其PIPs活性通过负性调控PKB/ Akt信号通路,在细胞的增殖、凋亡和分化等生物学行为中发挥了重要作用。此外,PTPRQ与一些疾病如失听及肿瘤性疾病有着密切的联系,如PTPRQ基因在遗传性听力疾病中存在突变; PTPRQ功能性缺失可影响PTPRQ-/-转基因小鼠耳蜗基底部毛细胞的发育,继而影响小鼠的高频听力等;在结直肠癌、肺癌、乳腺癌等多种肿瘤组织中存在PTPRQ的异常改变。但事实上,PTPRQ及其与疾病之间关系的研究目前尚处在起步阶段,还有许多的研究有待进行,包括PTPRQ生物学功能、PTPRQ相关信号通路、PTPRQ致患者失聪的发病机制及PTPRQ在肿瘤发病中的确切作用及其机制等一系列问题都亟待阐明。这些问题的解决不仅有助于我们对PTPRQ功能及其作用机制有更深入的理解,同时也将为疾病的靶向治疗发现新的作用靶点。
[参考文献]
[1]张丽景,金成艳,王果元,等.蛋白酪氨酸磷酸酶SHP-2 对AngⅡ刺激的心肌成纤维细胞增殖的影响[J].中国病理生理杂志,2012,28( 2) : 244-248.
[2]Bogdanova OV,Kot LI,Lavrova KV,et al.Modulation of protein tyrosine phosphorylation in gastric mucosa during re-epithelization processes[J].World J Biol Chem,2010,1( 11) : 338-347.
[3]Horvat-Brocker A,Reinhard J,Illes S,et al.Receptor protein tyrosine phosphatases are expressed by cycling retinal progenitor cells and involved in neuronal development of mouse retina[J].Neuroscience,2008,152( 3) : 618-645.
[4]Laczmanska L,Sasiadek MM.Tyrosine phosphatases as a superfamily of tumor suppressors in colorectal cancer[J].Acta Biochim Pol,2011,58( 4) : 467-470.
[5]Koskensalo S,Louhimo J,Hagstrom J,et al.Concomitant tumor expression of EGFR and TATI/SPINKL associates with better prognosis in colorectal cancer[J].PLoS One,2013,8( 10) : e76906.
[6]Anbalagan M,Moroz K,Ali A,et al.Subcellular localization of total and activated Src kinase in African American and Caucasian breast cancer[J].PLoS One,2012,7 ( 3) : e33017.
[7]Ren R.Mechanisms of BCR-ABL in the pathogenesis of chronic myelogenous leukaemia[J].Nat Rev Cancer,2005,5( 3) : 172-183.
[8]Oganesian A,Poot M,Daum G,et al.Protein tyrosine phosphatase RQ is a phosphatidylinositol phosphatase that can regulate cell survival and proliferation[J].Proc Natl Acad Sci U S A,2003,100( 13) : 7563-7568.
[9]Wright MB,Hugo C,Seifert RA,et al.Proliferating and migrating mesangial cells responding to injury express a novel receptor protein-tyrosine phosphatase in experimental mesangial proliferative glomerulonephritis[J].J BiolChem,1998,273( 37) : 23929-23937.
[10]Seifert RA,Coats SA,Oganesian A,et al.PTPRQ is a novel phosphatidylinositol phosphatase that can be expressed as a cytoplasmic protein or as a subcellularly localized receptor-like protein[J].Exp Cell Res,2003,287 ( 2) : 374-386.
[11]Yu KR,Kim YJ,Jung SK,et al.Structural basis for the dephosphorylating activity of PTPRQ towards phosphatidylinositide substrates[J].Acta Crystallogr D Biol Crystallogr,2013,69( Pt 8) : 1522-1529.
[12]Goodyear RJ,Legan PK,Wright MB,et al.A receptorlike inositol lipid phosphatase is required for the maturation of developing cochlear hair bundles[J].J Neurosci,2003,23( 27) : 9208-9219.
[13]Prakash S,McLendon HM,Dubreuil CI,et al.Complex interactions amongst N-cadherin,DLAR,and Liprin-alpha regulate Drosophila photoreceptor axon targeting[J].Dev Biol,2009,336( 1) : 10-19.
[14]Lee HK,Cording A,Vielmetter J,et al.Interactions between a receptor tyrosine phosphatase and a cell surface ligand regulate axon guidance and glial-neuronal communication[J].Neuron,2013,78( 5) : 813-826.
[15]Desai C,Purdy J.The neural receptor protein tyrosine phosphatase DPTP69D is required during periods of axon outgrowth in Drosophila[J].Genetics,2003,164( 2) : 575-588.
[16]Jung H,Kim WK,Kim DH,et al.Involvement of PTPRQ in differentiation during adipogenesis of human mesenchymal stem cells[J].Biophys Biochem Res Commun,2009,383( 2) : 252-257.
[17]Nayak G,Goodyear RJ,Legan PK,et al.Evidence for multiple,developmentally regulated isoforms of Ptprq on hair cells of the inner ear[J].Dev Neurobiol,2011,71 ( 2) : 129-141.
[18]Shahin H,Rahil M,Rayan AA,et al.Nonsense mutation of the stereociliar membrane protein gene PTPRQ in human hearing loss DFNB84[J].J Med Genet,2010,47 ( 9) : 643-645.
[19]Goodyear RJ,Jones SM,Sharifi L,et al.Hair bundle defects and loss of function in the vestibular end organs of mice lacking the receptor-like inositol lipid phosphatase PTPRQ[J].J Neurosci,2012,32( 8) : 2762-2772.
[20]Schraders M,Oostrik J,Huygen PLM,et al.Mutations in PTPRQ are a cause of autosomal-recessive nonsyndromic hearing impairment DFNB84 and associated with vestibular dysfunction[J].Am J Hum Genet,2010,86( 4) : 604-610.
[21]Sang Q,Mei H,Kuermanhan A,et al.Identification of a novel compound heterozygous mutation in PTPRQ in a DFNB84 family with prelingual sensorineural hearing impairment[J].Mol Genet Genomics,2015,290( 3) : 1135-1139.
[22]Leonardo S,Arkaitz C,Pier PP.Tenets of PTEN tumor suppression[J].Cell,2008,133( 3) : 403-414.
[23]Raftopoulou M,Etienne-Manneville S,Self A,et al.Regulation of cell migration by the C2 domain of the tumor suppressor PTEN[J].Science,2004,303( 5661) : 1179-1181.
[24]Tartaglia M,Niemeyer CM,Fragale A,et al.Somatic mutations in PTPN11 in juvenile myelomonocytic leukemia,myelodysplastic syndromes and acute myeloid leukemia[J].Nat Genet,2003,34( 2) : 148-150.
[25]Zhou X,Coad J,Ducatman B,et al.SHP2 is up-regulated in breast cancer cells and in infiltrating ductal carcinoma of the breast,implying its involvement in breast oncogenesis[J].Histopathology,2008,53( 4) : 389-402.
[26]Laczmanska I,Karpinski P,Kozlowska J,et al.Copy number alterations of chromosomal regions enclosing protein tyrosine phosphatase receptor-like genes in colorectal cancer[J].Pathol Res Pract,2014,210( 12) : 893-896.
[ABSTRACT]Receptor type protein tyrosine phosphatase Q ( PTPRQ) is an unusual protein tyrosine phosphatase that has intrinsic dephosphorylating activity for various phosphatidylinositiol and phospho-tyrosine substrates,especially the phosphatidylinositol activity.Recent data show that PTPRQ has an important role in various biological processes and is associated with some diseases.In this article,the structure and function of PTPRQ and the relationship between PTPRQ and diseases were briefly summarized.
Advance in receptor type protein tyrosine phosphatase Q
HUANG Yong-hong1,SHI Hui-mian1,ZHOU Xiao-yan1,XU Hong2,LUO Zhi-jun1,XU Fang-yun1
(1Department of Pathophysiology,2Department of Physiology,Medical College of Nanchang University,Nanchang 330006,China.E-mail: zluo559914@ gmail.com; scb9999@ sina.com)
[关键词]受体型蛋白酪氨酸磷酸酶Q;失聪;肿瘤;细胞增殖
[KEY WORDS]Protein tyrosine phosphatase,receptor type Q; Hearing loss; Neoplasms; Cell proliferation
[中图分类号]R363
[文献标志码]A
doi:10.3969/j.issn.1000-4718.2015.07.034
通讯作者△Tel: 0791-86360556; E-mail:罗志军zluo559914@ gmail.com;徐方云scb9999@ sina.com
*[基金项目]江西省自然科学基金资助项目( No.20151BAB205002) ;南昌大学博士后启动基金助项目( No.06301055)
[收稿日期]2015-01-19[修回日期]2015-05-19
[文章编号]1000-4718( 2015)07-1340-05
猜你喜欢
中国耳鼻咽喉颅底外科杂志(2022年6期)2023-01-06
作物学报(2022年3期)2022-01-22
天津医科大学学报(2021年4期)2021-08-21
中国现代药物应用(2020年10期)2020-06-18
中华耳科学杂志(2020年6期)2020-01-08
分析化学(2018年7期)2018-09-17
中成药(2017年12期)2018-01-19
中华耳科学杂志(2018年6期)2018-01-16
中成药(2017年8期)2017-11-22
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
