
宁夏荒漠草原优势植物生长及生物量分配对放牧干扰的响应
2015-07-04 11:14安钰,安慧
西北植物学报 2015年2期
安 钰,安 慧
(1 宁夏农林科学院荒漠化治理研究所,银川750002;2 宁夏大学 西北退化生态系统恢复与重建教育部重点实验室/西部生态与生物资源开发联合研究中心,银川750021)
植物生物量作为草地生态系统最基本的数量特征,对认识草原生态系统结构和功能起着关键性作用[1-2]。生物量的变化与环境条件密切相关,每一种植物生物量的动态变化都影响草原群落的发生和发展,影响程度因不同植物种类而不同[3]。植物在整个生长发育阶段中,生长、繁殖和维持等功能对有限资源始终存在着竞争[4-6],植物不得不调节其生物量以权衡这些功能间的资源分配[7],在一定程度上反映了植物生长发育对环境的响应和适应,植物在不同环境下的资源分配格局与其适合度可能有关[8]。
放牧是草地利用的主要方式,能对草原植物产生一系列的影响,动物的采食和践踏影响植物的生长和耐性,同时放牧干扰是草地生态系统最重要的人为扰动因素[3,9]。植物在长期放牧过程中,已经进化出多种适应机制,在形态、生长、繁殖和资源分配等方面表现出相应变化[3]。
甘草(Glycyrrhizauralensis)为豆科多年生草本植物,根系发达,多生长于生态环境十分脆弱的干旱半干旱荒漠草原区,具有耐热、耐旱、耐寒,抗盐碱等特点,是荒漠半荒漠地区保持水土、改良土壤、防风固沙的主要植物之一[10-12]。研究多集中于资源开发、药理作用、化学成分分析、临床应用等方面[12-15]。牛心朴子(Cynanchumkomarovii)为萝摩科鹅绒藤属多年生草本植物,俗称老瓜头、芦心草等,其根系发达,耐寒,抗风沙,且繁殖力强,生长于荒漠草原和次生沙漠的半固定沙丘和荒漠流沙区等干旱地带[16]。有关荒漠植物牛心朴子的研究主要集中在化学成分,药理和药效、生物农药等方面[17],也有人认为牛心朴子是在沙化过程中,适应其环境而保存下来的沙漠生态型植物,是土地严重沙质荒漠化的指示植物之一,也是草原逆行演替过程中最后阶段的指示物种[18]。
随着宁夏荒漠草原土壤沙化、植被退化等问题的日益严重,甘草及牛心朴子在草地生态系统中的作用已越来越引起有关学者的重视。本研究选择长期适应于干旱生态环境下的耐旱沙生植物甘草、牛心朴子为研究对象,对不同放牧强度下甘草和牛心朴子的形态变化特征及各植物构件生物量分配进行了研究,旨在探索不同放牧强度对其生物量分配策略的影响机制,为探讨退化草原的恢复演替提供科学依据。
1 材料和方法
1.1 研究区自然概况
研究区位于宁夏盐池县境内东南部(37°05′~38°10′N,106°33′~107°47′E,海拔高度1 295~1 951m),年均气温8.1℃,极端最高均温34.9℃,极端最低均温-24.2℃,年均无霜期165d,年均降雨量250~350mm,其中70%以上降雨集中在6~9月,降雨年际变化率大;年蒸发量2 710mm,年均风速2.8m/s,属于典型中温带大陆性气候,是干旱与半干旱气候的过渡地带。土壤类型以灰钙土为主,其次是风沙土和黑垆土,此外有黄土,少量盐土、白浆土等[19-20]。土壤质地多为轻壤土、沙壤土和沙土,结构松散,肥力较低。
该区植被类型有灌丛、草原、草甸、沙地植被和荒漠植被,其中灌丛、草原、沙地植被数量较大,分布广泛,群落中常见植物种类以旱生和中旱生类型为主[21]。荒漠植被类型有沙蒿(Artemisiadesertorum)、赖草(Leymussecalinus)、蒙古冰草(Agropyronmongolicum)、短花针茅(Stipabreviflora)、甘草(G.uralensis)、牛心朴子(C.komarovii)、二裂委陵菜(Potentillabifurca)、川青锦鸡儿(Caraganatibetica)、猫头刺(Oxytropisaciphylla)、和盐爪爪(Kalidiumfoliatum)等。
1.2 试验设计
试验样地设置在盐池县高沙窝镇草原资源生态监测站的围封草地及附近的退化荒漠草地。试验于2004年开始,每年5月开始放牧,10月放牧结束。由于长期放牧活动对植被群落有很大的影响,参照任继周[22]对放牧强度和草地演替阶段的划分标准,根据草地地上生物量、家畜理论采食量和草场面积及放牧时间,设置4 个处理:围封禁牧(NG,0 只/hm2)、轻度放牧(LG,0.45 只/hm2)、中度放牧(MG,1.00 只/hm2)和重度放牧(HG,1.50 只/hm2)。每个处理重复3次,共12块样地;各放牧强度样地面积为6hm2,且样地间草地发育环境及地形条件一致。
1.3 研究内容和方法
于2011年,在植物生长旺季(8月)进行野外观测及采样。在围封禁牧(NG)、轻度放牧(LG)、中度放牧(MG)和重度放牧(HG)区分别随机选取甘草和牛心朴子各15株,测定株高及地径。之后将整株植物全部挖出,取样深度30cm,在挖取过程中注意保持植物地上部与根系的自然联系,将植株带土取出放入封口袋带回实验室。在实验室内用水将泥土冲洗干净,按根、茎、叶分开,并在80℃下烘干48h,测定各器官的干重。计算根生物量比(根系生物量/植株总生物量)、茎生物量比(茎生物量/植株总生物量)、叶生物量比(叶生物量/植株总生物量)、根冠比(根系生物量/地上部分生物量)、叶茎比(叶生物量/茎生物量)。
1.4 数据处理
所有数据采用JMP 9软件进行统计分析,利用单因素方差分析(One-way ANOVA)分析不同放牧强度处理对植物生长各个指标的影响,差异显著性水平设置为α=0.05,有显著效应的用小写字母a、b、c等加以标注。
2 结果与分析
2.1 放牧干扰下甘草和牛心朴子生长特征的变化
随着放牧强度的增加,荒漠草原优势植物甘草和牛心朴子的生长特征发生了明显的变化。不同放牧强度间甘草株高和地径、牛心朴子株高差异显著(P<0.05),而牛心朴子地径在各放牧强度间差异不显著(图1)。甘草株高和地径、牛心朴子株高随着放牧强度的增加呈先升高后下降的趋势,均在轻度放牧条件下最高,重度放牧时则显著降低;而牛心朴子地径在中度放牧时达到最低值。轻度放牧条件下甘草株高和地径、牛心朴子株高较围封禁牧分别升高了20.75%、2.95%、3.22%,而重度放牧时甘草株高和地径、牛心朴子株高较围封禁牧下降了45.83%、19.84%、17.34%。
2.2 放牧干扰下甘草和牛心朴子生物量的变化
放牧干扰下,荒漠草原优势植物甘草和牛心朴子各构件的生物量均发生了不同程度的变化(图2),甘草和牛心朴子的茎生物量、叶片生物量和总生物量随着放牧强度的增加呈先升高后降低的趋势,且处理间差异性显著(P<0.05),而甘草和牛心朴子根系生物量随放牧强度的加强变化趋势不同。轻度放牧条件下甘草与牛心朴子的茎生物量、叶生物量和总生物量都达到最大值。重度放牧条件下甘草总生物量、茎生物量、叶生物量低于围封禁牧及轻度放牧。甘草根系生物量随着放牧强度的增加而下降,且处理间差异性不显著,重度放牧条件下根系生物量较围封禁牧、轻度放牧、中度放牧处理分别减小了55.6%、54.8%、44.9%。牛心朴子根系生物量呈先增加后降低趋势,且重度放牧条件下根系生物量显著低于围封禁牧、轻度放牧及中度放牧,分别降低73.6%、73.5%、76.3%。
2.3 放牧干扰下甘草和牛心朴子生物量分配变化

图1 不同放牧干扰下甘草和牛心朴子生长特征变化Fig.1 Change of plant growth of G.uralensis and C.komarovii under different grazing disturbances

图2 不同放牧干扰下甘草和牛心朴子单株生物量的变化Fig.2 Change of individual plant biomass of G.uralensis and C.komarovii under different grazing disturbances
通过对放牧干扰下荒漠草原优势植物甘草和牛心朴子各构件生物量的分配研究表明(表1),甘草和牛心朴子生物量分配的总体格局是根>叶>茎。随着放牧强度的增加,甘草根生物量比呈先升高后降低趋势,且不同处理间差异不显著;而牛心朴子根生物量比在中度放牧条件下达到最大值,且与轻度放牧、重度放牧之间差异显著(P<0.05),与围封禁牧之间差异不显著。不同放牧强度下甘草的茎生物量比差异不显著,随着放牧强度的增加呈下降的趋势;与围封禁牧区相比,轻度放牧、中度放牧、重度放牧分别降低了13.2%,32.4%,22.7%。牛心朴子茎生物量比在围封禁牧与中度放牧、轻度放牧与重度放牧之间差异不显著,而其它处理间差异显著,在重度放牧强度下达到最大值。甘草和牛心朴子叶生物量比的变化趋势一致,且处理之间差异不显著,均在重度放牧区达到最大值。
不同放牧强度下甘草和牛心朴子根冠比的变化趋势相同,均在中度放牧条件下达到最大值,在重度放牧条件下根冠比小于1。甘草根冠比在不同放牧强度下差异不显著,而牛心朴子根冠比在围封禁牧与中度放牧、轻度放牧与重度放牧处理间差异不显著,其它处理间差异显著。不同放牧强度下甘草和牛心朴子叶茎比的变化趋势不同,随着放牧强度的增加甘草叶茎比呈上升趋势,在围封禁牧与轻度放牧、中度放牧与重度放牧处理间差异不显著,而其它处理间差异显著;牛心朴子叶茎比在围封禁牧与中度放牧、轻度放牧与重度放牧处理间差异不显著,其它处理间差异显著,且牛心朴子叶茎比在围封禁牧强度下显著高于重度放牧和轻度放牧。
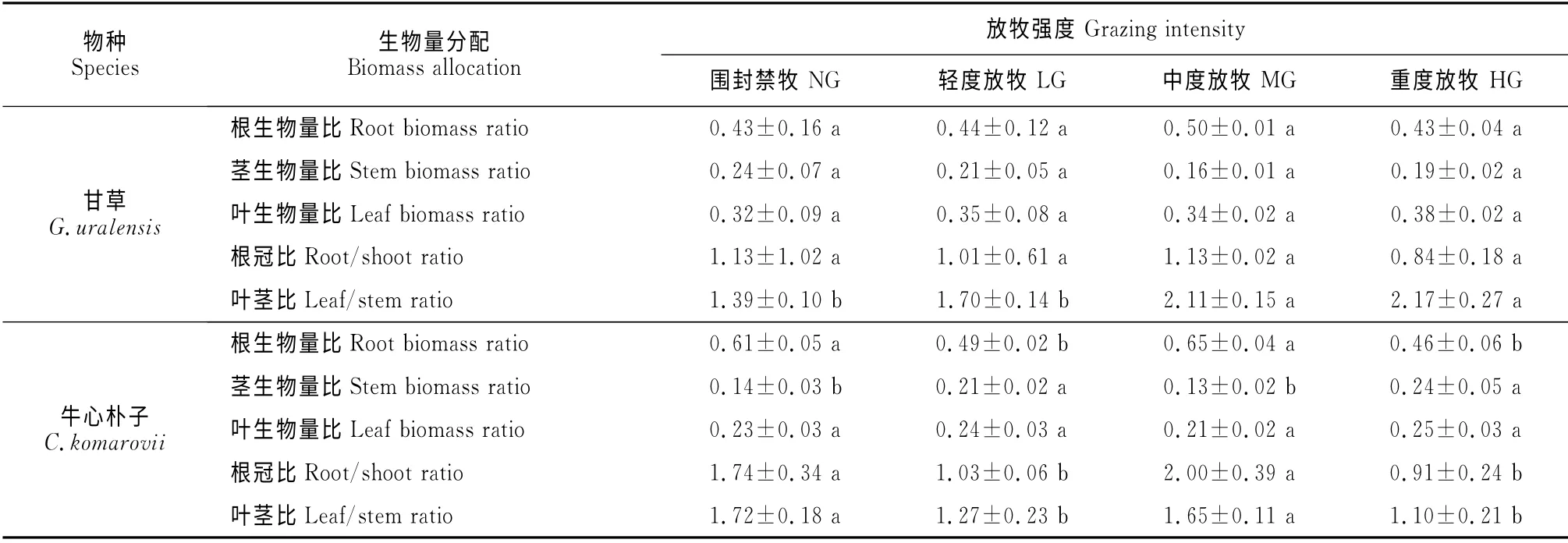
表1 不同放牧干扰下甘草和牛心朴子生物量分配的变化Table 1 Changes of biomass allocation of G.uralensis and C.komarovii under different grazing disturbances
3 讨 论
当前有很多关于植物功能特征对放牧响应的研究[23-24],植物功能性特征为适应放牧地环境的变化而发生一系列的改变,如植物高度和叶面积等明显变小[24-25]。本研究中,不同放牧强度间甘草株高和地径、牛心朴子株高差异显著(P<0.05),在重度放牧条件下显著降低;而牛心朴子地径在中度放牧时达到最低值,且不同处理间差异不显著。
放牧干扰对青藏高原高寒草甸、灌丛草甸植物群落特征研究表明,随着放牧强度的增加,地上生物量呈显著降低趋势[26],内蒙古典型草原、沙质草地植物地下生物量随着放牧强度的增大呈现降低趋势[27-28],而小蒿草草甸植物地下生物量随着放牧强度的增加而增加[29]。本研究中荒漠草原植物甘草和牛心朴子的茎生物量、叶片生物量和总生物量随着放牧强度的增加呈先升高后降低的趋势,且处理间差异性显著(P<0.05);甘草和牛心朴子根系生物量随放牧强度的加强变化趋势不同,甘草根系生物量随着放牧强度的增加而下降,牛心朴子根系生物量呈先增加后降低趋势,这表明轻度的放牧干扰能够使植物总生物量及地上、地下生物量增加,但过度放牧将导致植物总生物量及地上、地下生物量降低,这是植物对放牧干扰的一种响应与适应。
植物根、茎、叶等器官的生物量分配格局反映了植物对放牧干扰的生态适应对策变化,植物总是通过调整自身的生长方式和资源分配来适应环境[3,8,30]。有研究表明,在不同胁迫下,植物通过调节光合产物在地上、地下部分的分配,体现不同的适应性[31]。在本研究中,甘草根系生物量分配在不同放牧强度下差异不显著,牛心朴子根系生物量分配在中度放牧条件下达到最大值,且与轻度放牧、重度放牧之间差异显著(P<0.05),与围封禁牧之间差异不显著。随着放牧强度的增加,甘草的茎生物量比呈下降的趋势,叶生物量比呈上升趋势,即在放牧干扰下,资源被优先分配给光合器官——叶片,这与任海彦等[25]研究结果基本一致。牛心朴子的茎生物量和叶生物量变化趋势一致,均随着放牧强度增加呈先增加后下降的趋势,而叶茎比则随着放牧强度的增加呈先减小后增加的趋势。这说明植物生物量在地上、地下不同器官的分配因植物种而异,体现了一定表型可塑性,这种差异反映出植物生态适应策略的不同。
[1]陈佐忠,汪诗平.中国典型草原生态系统[M].北京:科学出版社,2000:49-66.
[2]ZHAO B B(赵彬彬),NIU K CH(牛克昌),DU G ZH(杜国祯).The effect of grazing on above-ground biomass allocation of 27plant species in an alpine meadow plant community in Qinghai-Tibetan Plateau[J].ActaEcologicaSinica(生态学报),2009,29(3):1 596-1 606(in Chinese).
[3]LI J H(李金花),LI ZH Q(李镇清).Clonal morphological plasticity and biomass allocation pattern ofArtemisiafrigidaandPotentilla acaulisunder different grazing intensities[J].ActaPhytoecologicaSinica(植物生态学报),2002,26(4):435-440(in Chinese).
[4]HU J CH(胡继超),JIANG D(姜 东),CAO W X(曹卫星),etal.Effect of short-term drought on leaf water potential,photosynthesis and dry matter partitioning in paddy rice[J].ChineseJournalofAppliedEcology(应用生态学报),2004,15(1):63-67(in Chinese).
[5]LI H(李 红),YANG Y F(杨允菲).Effect of restorative measures on quantitative characters of reproduction forLeymuschinensispopulation in the degenerated grassland[J].ChineseJournalofAppliedEcology(应用生态学报),2004,15(5):819-823(in Chinese).
[6]WU CH(吴 楚),FAN ZH Q(范志强),WANG ZH Q(王政权).Effect of phosphorus stress on chlorophyll biosynthesis,photosynthesis and biomass partitioning pattern ofFraxinusmandchuricaseedlings[J].ChineseJournalofAppliedEcology(应用生态学报),2004,15(6):935-940(in Chinese).
[7]张大勇.理论生态学研究[M].北京:高等教育出版社,2000:32-38.
[8]LI J H(李金花),LI ZH Q(李镇清),LIU ZH G(刘振国).Growth and resource allocation pattern ofArtemisiafrigidaunder different grazing and clipping intensities[J].ChineseJournalofAppliedEcology(应用生态学报),2004,15(3):408-412(in Chinese).
[9]WANG SH P(汪诗平).Grazing resistance of rangeland plants[J].ChineseJournalofAppliedEcology(应用生态学报),2004,15(3):517-522(in Chinese).
[10]ZHANG J(张 继),YAO J(姚 健),DING L(丁 兰),etal.Advancement of research on the utilization ofGlycyrrhiza[J].Grassland andTurf(草原与草坪),2000,89(2):12-17(in Chinese).
[11]KONG H(孔 红),YAN X Y(闫训友),SHI ZH X(史振霞).Progress research onGlycyrrhiza[J].NorthernHorticulture(北方园艺),2007,(7):70-72(in Chinese).
[12]WANG ZH L(王照兰),DU J C(杜建材),YU L Q(于林清).The utilization value,current research and problems ofGlycyrrhiza[J].GrasslandofChina(中国草地),2002,24(1):73-76(in Chinese).
[13]BAI H,LI W,KOIKE K,etal.A novel biflavonoid from roots ofGlycyrrhizauralensiscultivated in China[J].Chemical&PharmaceuticalBulletin,2003,51(9):1 095-1 097.
[14]MA C J,LI G S,ZHANG D L,etal.One step isolation and purification of liquiritigenin and isoliquiritigenin fromGlycyrrhizauralensisRisch.using high-speed counter-current chromatography[J].JournalofChromatographyA,2005,1 078(1-2):188-192.
[15]LUO J Y(罗婧莹),HUANG X(黄 熙),CHEN D H(陈德华),etal.Observation of therapeutic efficacy of stronger neo-minophagen C combined with salvia miltiorrhiza injection on treating systemic sclerosis[J].TheChineseJournalDermatovenereology(中国皮肤性病学杂志),2009,23(7):411-412(in Chinese).
[16]DUAN H J(段海婧),HAN T(韩 婷),WU X L(吴秀丽),etal.Separation and identification of endophytic fungi from desert plantCynanchumkomarovii[J].ChinaJournalofChineseMateriaMedica(中国中药杂志),2013,38(3):325-330(in Chinese).
[17]CHEN SH SH(陈绍淑),HE SH H(何生虎).Progress on chemical constituents,bioactivity,development and utilization ofCynanchum komarovii[J].ProgressinVeterinaryMedicine(动物医学进展),2006,27(7):46-50(in Chinese).
[18]MI H L(米海莉),XU X(许 兴),LI SH H(李树 华),etal.Physiological response and adapting mechanism ofCynanchumkomarociiseedlings under drought stress[J].AgriculturalResearchintheAridAreas(干旱地区农业研究),2002,20(4):11-16(in Chinese).
[19]XU D M(许冬梅),WANG K(王 堃).Characteristics of soil microbe in southern ecotone of Mu Us sandy land[J].JournalofDesert Research(中国沙漠),2007,27(5):805-808(in Chinese).
[20]YANG Y(杨 越),HASI(哈 斯),SUN B P(孙保平),etal.Effects of different vegetation restoration types on soil nutrients in southern edge of Mu Us sandy land[J].ChineseAgriculturalScienceBulletin(中国农学通报),2012,28(10):37-42(in Chinese).
[21]ZHANG K B(张克斌),LI R(李 瑞),HOU R P(侯瑞萍),etal.Study on plant diversity of different control measures of desertification in Yanchi County,Ningxia[J].ScienceofSoilandWaterConservation(中国水土保持科学),2004,2(4):66-72(in Chinese).
[22]任继周.草业科学研究方法[M].北京:中国农业出版社,1998:42-48,207-211.
[23]CINGOLANI A M,POSSE G,COLLANTES M B.Plant functional traits,herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands[J].JournalofAppliedEcology,2005,42(1):50-59.
[24]DIAZ S,LAVOREL S,McINTYRE S,etal.Plant trait responses to grazing-aglobal synthesis[J].GlobalChangeBiology,2007,13(2):313-341.
[25]REN H Y(任海彦),ZHENG SH X(郑淑霞),BAI Y F(白永飞).Effects of grazing on foliage biomass allocation of grassland communities in Xilin River Basin,Inner Mongolia[J].ChineseJournalofPlantEcology(植物生态学报),2009,33(6):1 065-1 074(in Chinese).
[26]REN Q J(仁青吉),WU G L(武高林),REN G H(任国华).Effect of grazing intensity on characteristics of alpine meadow communities in the eastern Qinghai-Tibetan Plateau[J].ActaPrataculturaeSinica(草业学报),2009,18(5):256-261(in Chinese).
[27]WANG Y F(王艳芬),WANG SH P(汪 诗平).Influence of different stocking rates on belowground biomass in Inner Mongolia steppe[J].ActaAgrestiaSinica(草地学报),1999,7(3):198-203(in Chinese).
[28]ZHAO H L(赵哈林),ZHAO X Y(赵学勇),ZHANG T H(张铜会),etal.Study on damaged process of the vegetation under grazing stress in sandy grassland[J].ActaEcologicaSinica(生态学报),2003,23(8):1 505-1 511(in Chinese).
[29]WANG Q L(王启兰),CAO G M(曹广民),WANG CH T(王长庭).The impact of grazing on the activities of soil enzymes and soil environmental factors in alpineKobresiapygmaeameadow[J].PlantNutritionandFertilizerScience(植物营养与肥料学报),2007,13(5):856-864(in Chinese).
[30]XILIN T Y(锡林图雅),XU ZH(徐 柱),ZHENG Y(郑 阳).Influence of different stocking rates on underground biomass and net primary productivity onStipakryloviisteppe in Inner Mongolia[J].ChineseJournalofGrassland(中国草地学报),2009,31(3):26-29(in Chinese).
[31]WANG R Z,GAO Q,CHEN Q S.Effects of climatic change on biomass and biomass allocation inLeymuschinensis(Poaceae)along the North-east China Transect(NECT)[J].JournalofAridEnvironments,2003,54(4):653-665.
猜你喜欢
中老年保健(2021年9期)2021-08-24
电脑报(2020年12期)2020-06-30
看历史(2018年1期)2018-02-07
中成药(2018年1期)2018-02-02
中国民族医药杂志(2017年7期)2017-12-21
农民致富之友(2017年15期)2017-08-20
晚报文萃(2016年7期)2017-01-10
中国果业信息(2015年10期)2015-01-23
中医研究(2014年2期)2014-03-11
中成药(2014年10期)2014-02-28
