
扎龙保护区丹顶鹤秋季迁徙季节行为节律的研究
2015-07-05 17:13李淑玲马杰王文锋杜微
东北农业大学学报 2015年7期
李淑玲,马杰,王文锋,杜微
扎龙保护区丹顶鹤秋季迁徙季节行为节律的研究
李淑玲1,马杰1,王文锋2,杜微2
(1.东北农业大学生命科学学院,哈尔滨150030;2.黑龙江扎龙国家级自然保护区,黑龙江齐齐哈尔161000)
采用瞬时扫描取样法观察研究扎龙保护区野生丹顶鹤和散养丹顶鹤秋季迁徙季节日行为节律和时间分配。结果表明,野生丹顶鹤在此期间,觅食行为时间分配显著高于散养丹顶鹤,差异均达极显著;散养丹顶鹤整理行为和其他行为(驱赶、鸣叫)显著高于野生丹顶鹤;成年个体两性间行为时间分配也有一定差异。观察发现,野生丹顶鹤大量觅食,是为长途迁徙储备能量;散养丹顶鹤作为不迁徙种群,整理行为和其他行为的时间分配表现出对保护区环境的适应性。野生和散养丹顶鹤家庭组中个体间行为差异体现丹顶鹤个体在家庭组中地位和职责有所不同。从野生丹顶鹤和散养丹顶鹤在秋季迁徙季节中行为节律观察,建立行为谱,可了解野生丹顶鹤和散养丹顶鹤行为差异,改进人工饲养技术,改善丹顶鹤生存环境,也为完善丹顶鹤行为生态研究提供新资料。
扎龙保护区;日活动节律;丹顶鹤;适应性;行为谱
丹顶鹤(Grus japonensis),属鹤形目,鹤科,大型珍稀涉禽,是国家Ⅰ级保护动物。在世界自然保护联盟(IUCN)红皮书中属濒危物种,在濒危野生动植物种国际贸易公约(CITES)中被列入附录一。丹顶鹤在全世界有两大种群,即日本北海道留鸟种群和东亚大陆候鸟种群[1]。现存野生丹顶鹤种群数量约2 600只[3]。在我国主要繁殖地为中国黑龙江省扎龙自然保护区,每年有400~450只繁殖群[1-3],包括人工饲养丹顶鹤不迁徙种群和野生丹顶鹤迁徙种群。
行为节律与行为时间分配是动物行为学研究重要内容,与动物能量摄取和消耗联系密切,而这些又随环境条件变化而改变[4]。动物行为节律既是野生动物自身进化适应的一部分,也是适应环境条件变化的一种体现[5]。行为时间分配是研究鸟类各种行为的重要手段,有利于对鸟类生境选择和取食之间关系的理解[6],对于丹顶鹤非繁殖季节行为研究,有助于分析丹顶鹤在不同环境中的适应策略和行为特点,也能在不同程度上了解丹顶鹤生存状况,为保护丹顶鹤提供理论依据,具有重要理论和实践意义。本研究在黑龙江扎龙国家级自然保护区进行,对野生丹顶鹤与散养丹顶鹤秋季迁徙期间各种行为节律及行为时间分配进行观察记录,通过行为观察记录数据,分析野生丹顶鹤与散养丹顶鹤非繁殖季节同环境下生存策略和行为习性,探讨野生丹顶鹤与散养丹顶鹤行为差异及生态学问题,以补充丹顶鹤非繁殖期行为学数据,为保护丹顶鹤提供科学依据。
1 材料与方法
1.1 研究地状况
黑龙江扎龙国家级自然保护区(以下简称扎龙保护区)位于黑龙江省西部乌裕尔河下游齐齐哈尔市与富裕、泰来、林甸、杜尔伯特蒙古族自治县交界地域,保护区面积21万hm2。地处中温带,属寒温带大陆性季风气候,年平均气温1~3℃,最高气温36.6℃,最低气温-35.0℃,无霜期128 d,年均降水量380~470 mm,多集中于6~9月,年均蒸发量1 336 mm,为降水量的3~4倍。扎龙保护区土壤以草甸沼泽土为主,其次是潜育草甸土和碳酸盐草甸土。保护区植被主要包括草甸草原植被、草甸植被、沼泽植被、水生植被四种类型,其中高等植物有468种。扎龙湿地于1979年被批准建立省级自然保护区,1987年晋升为国家级自然保护区,1992年被首批列入国际重要湿地名录,是中国著名的以保护丹顶鹤等珍贵水禽为主的沼泽湿地类型自然保护区[7]。
1.2 研究方法
研究先于2013年9月26~30日在扎龙自然保护区进行预试验,观察确定丹顶鹤活动频繁区域及在该时期主要行为,并建立行为谱。观察固定在保护区内的望鹤楼,核心区47°10'46''N,124°06' 14''E,试验正式于2013年10月8~20日在扎龙保护区进行,期间天气多晴少云,温度2~8℃。
1.2.1 觅食行为
寻食,低头用微张开的喙尖部插入泥土或水中寻找食物;处理食物,将食物啄碎并逐个吞下;饮水,喙半开,下喙浸入水中将水舀起抬头吞咽。
1.2.2 警戒行为
张望,头部抬起并缓慢左右转动,眼睛仔细观察周围环境情况,停止正在进行的活动;鸣声恐吓,在外界刺激发生时,一只雄性丹顶鹤首先伸颈大声连续“gao~gao~”长鸣提醒其他个体。
1.2.3 运动行为
行走,双脚交替迈步前行;飞行,扇动双翼在空中飞行;跳跃,双脚用力蹬地腾空,并伴有展翅动作。
1.2.4 整理行为
理羽,用喙张合轻啄划动梳理羽毛;立羽,竖起体羽,羽毛分开;抖羽,以身体为中心,迅速左右摆动,并伴有立羽行为;还有蹭枕,抓挠,摆尾等。
1.2.5 静栖行为
卧息,将双脚折置于胸腹部下方,头向前下垂,或是将头转向后方,喙插入翅膀或搭在背上休息;单脚站立休息,单脚支撑,另一只脚收起,将头向前下垂或是转向后方,喙插入翅膀或搭在背上休息。
1.2.6 其他行为
鸣叫行为,有配偶之间的对鸣,呼唤子鹤的鸣叫,还有雄鹤在指挥飞行时发出的声音;冲突打斗行为。若遇见其他丹顶鹤,或是对威胁物警告无果,采取攻击性行为,用喙猛刺向对方,或腾空用喙和爪子攻击,并用身体压向对方;逃避行为,丹顶鹤对异物不愿采用攻击性行为,则会选择后退、惊飞、逃离,免受异物威胁。
在固定点用10×25双筒望远镜对野生和散养丹顶鹤进行全天观察,每天5:00~17:00,采用瞬时扫描取样法对研究个体进行行为观察。每5 min记录1次丹顶鹤行为,每小时扫描12次,每天扫描144次。
1.3 观察对象的选择与个体识别
在扎龙保护区内选择已配对并带有当年出生长成亚成体鹤的野生丹顶鹤和散养丹顶鹤。以腿部是否绑有脚环,和行为表现对人类活动的耐受性高低,作为区分野生丹顶鹤和散养丹顶鹤的依据。
野生丹顶鹤通常会选择人类活动较少,植被保存完好的生活环境,对人类活动因素耐受性较低,当环境有人类活动出现并遭到破坏,野生丹顶鹤会放弃该活动区域并逃离。观察时,绑有脚环,对人类活动耐受性较高的为人工饲养丹顶鹤;无脚环,对人类活动耐受性较低的为野生丹顶鹤。
雌雄丹顶鹤形态差异较小,仅从体型、外貌特征不能准确判断性别。在雌雄齐声鸣叫时,雄性会仰颈,喙指向天空,发出“gao~,gao~…”长时间单音节叫声。雌性会伸颈,喙则指向斜前方,随附“gaogao,gaogao…”短而连续的单音节鸣叫;在观察期间,雄性丹顶鹤行为活动相对家庭组其他成员较独立,会在雌性丹顶鹤与子鹤采食、静栖时,站立一旁张望警戒;在游走、飞行时,一直在最前面;遇到危险,则首当其冲。因在非繁殖季节,雄性丹顶鹤没有繁殖压力,会以更多时间保护家庭组成员领域免受其他因素干扰。通过多对成年丹顶鹤行为观察,证实雄性丹顶鹤在家庭组中的行为特点,以此作为区别雌雄丹顶鹤的主要依据,但对未配对的丹顶鹤性别区分,该方法效果不明显。
1.4 数据处理
在固定点观察期间,有多对丹顶鹤在该区域活动,选取野生丹顶鹤和散养丹顶鹤各3对,每个家庭组带有2013年出生的亚成体鹤,共观察丹顶鹤18只。于2013年10月17日固定点观察,未发现野生丹顶鹤,于是以固定点为中心向四周5 km距离,徒步搜寻野生丹顶鹤,后3 d以同样方式,加大搜寻范围,仅有零散几对野生丹顶鹤,观察记录作为数据补充,共记录各行为发生频次22 464次。对野生和散养丹顶鹤行为观察发现,在同样环境条件下,不同种群行为有鲜明特点,同种群行为模式基本相同。对观察获得的各种行为数据用Microsoft Excel 2003软件进行整理,行为节律以各时间段行为发生平均频次表示,并作混合折线图,行为时间分配以各行为发生频次占总行为频次百分比表示,节律变化使用SPSS 19.0软件,以性别年龄(雌雄幼3个水平)和生存状态(野生、散养两个水平)为固定因素,对各行为节律变化进行双因素方差分析,并用LSD后期检验。
2 结果与分析
2.1 野生丹顶鹤行为
扎龙保护区野生丹顶鹤在该时期行为时间分配见表1。其中,无论雄性、雌性还是亚成体,觅食行为所占比例均最高,达60%以上。野生丹顶鹤觅食行为占比全天均保持很高状态(见图1、3、5),其中亚成体鹤觅食行为最高,其与雄性和雌性相比较,分别达到极显著和显著差异(见表2),雄性和雌性差异不显著。
野生丹顶鹤觅食行为在13:00~14:00出现低谷(见图1、3、5),同时野生雄性丹顶鹤警戒行为达到最高(见图2),野生雌性丹顶鹤和亚成体鹤在此时出现静栖行为,警戒行为时间分配较雄性丹顶鹤少(见图4、6)。野生雄性丹顶鹤全天警戒行为比野生雌性丹顶鹤和亚成体鹤高(见表1),差异达到极显著(见表2),野生雄性丹顶鹤静栖行为比野生雌性丹顶鹤和亚成体鹤均低(见表1),但经双因素方差分析LSD后期检验,差异性并未达到显著水平(见表2)。
雄性、雌性和亚成体运动行为未达显著差异(见表2),在12:00~13:00和15:00~16:00产生运动行为高峰(见图1、3、5),同时觅食行为和静栖行为降低。野生丹顶鹤雄性、雌性和亚成体间整理行为和其他行为全天较平均,差异不显著(见表2)。
2.2 散养丹顶鹤行为
如表1所示,扎龙保护区,散养丹顶鹤行为时间分配觅食行为最高,食源主要靠人工投食,保护区一般在8:30和15:30投食,投喂鲜鱼和鲜玉米,散养丹顶鹤觅食高峰出现在8:00~9:00、11: 00~12:00、13:00~14:00和16:00~17:00(见图1、 3、5),亚成体觅食行为比雄性和雌性高,均达到极显著差异,雄性和雌性觅食行为达差异显著(见表2)。
散养丹顶鹤雄性、雌性和亚成体警戒行为于12:00过后,逐渐上升,并在14:00~15:00达到峰值(见图2、4、6),个体间警戒行为差异均极显著(见表2),散养丹顶鹤于12:00~13:00会出现静息行为(见图2、4、6),运动行为、整理行为和其他行为只有雄性与亚成体差异显著,静栖行为差异不显著(见表2)。

图1 雄性丹顶鹤野生和散养的觅食、整理、运动行为日节律Fig.1Daily rhythm of foraging,preening and sports behaviours of the wild and the semi-captive male Grus japonensis

图2 雄性丹顶鹤野生和散养的静栖、警戒、其他行为日节律Fig.2Daily rhythm of resting,vigilance and other behaviours of the wild and the semi-captive male Grus japonensis

图3 雌性丹顶鹤野生和散养的觅食、整理、运动行为日节律Fig.3Daily rhythm of foraging,preening and sports behaviours of the wild and the semi-captive female Grus japonensis

图4 雌性丹顶鹤野生和散养的静栖、警戒、其他行为日节律Fig.4Daily rhythm of resting,vigilance and other behaviours of the wild and the semi-captive female Grus japonensis
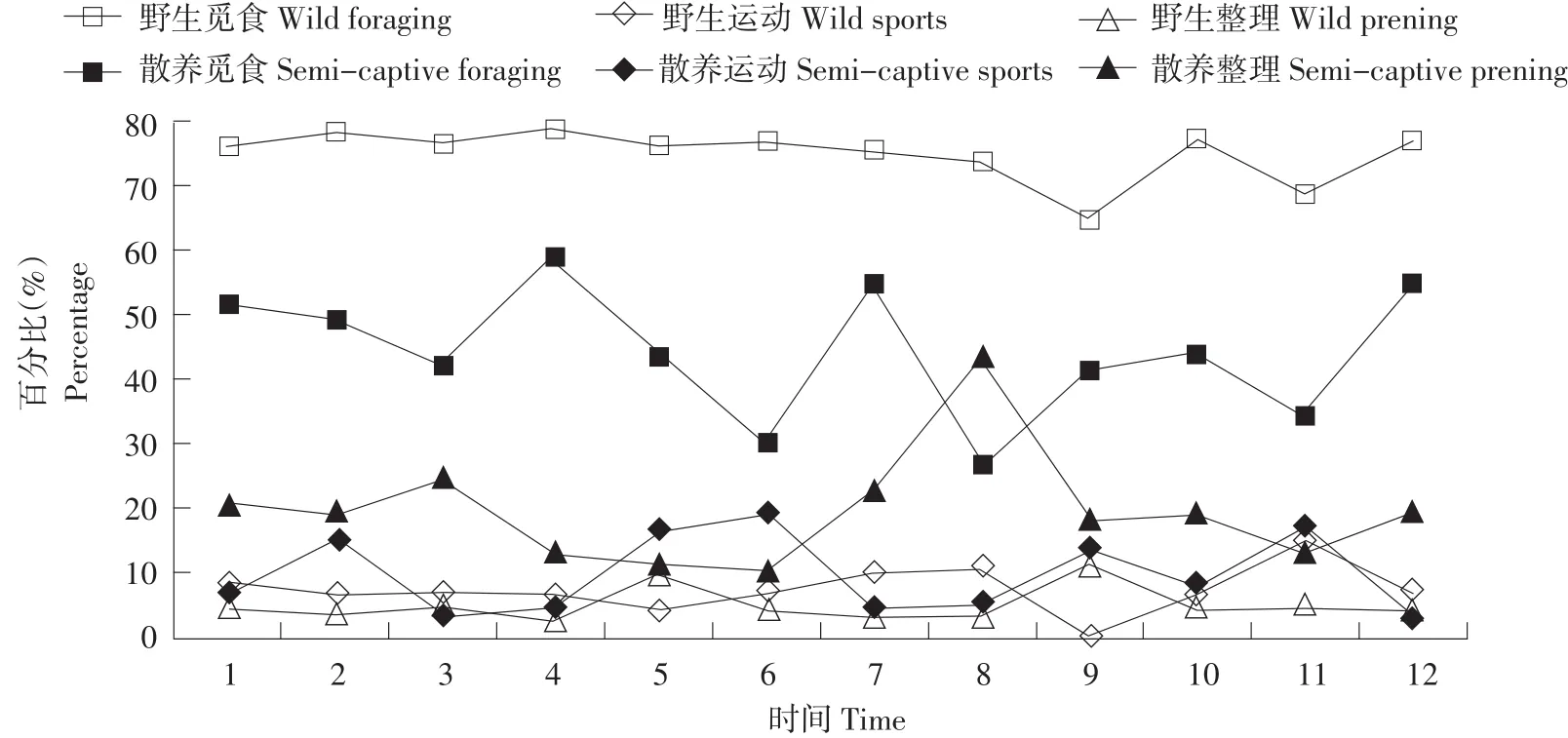
图5 亚成体鹤野生和散养的觅食、整理、运动行为日节律Fig.5Daily rhythm of foraging,preening and sports behaviours of the wild and the semi-captive sub-mature Grus japonensis
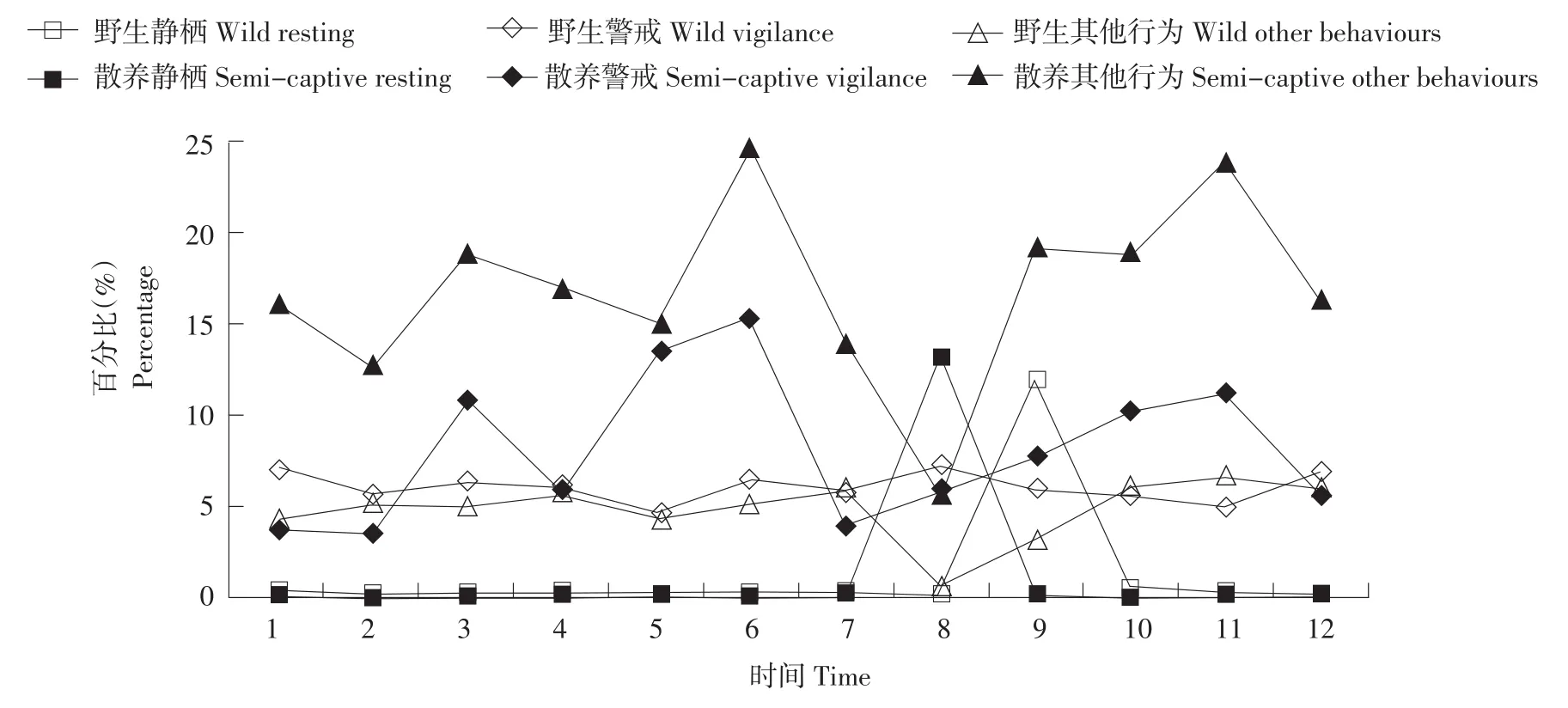
图6 亚成体鹤野生和散养的静栖、警戒、其他行为日节律Fig.6Daily rhythm of resting,vigilance and other behaviours of the wild and the semi-captive sub-mature Grus japonensis
2.3 野生丹顶鹤和散养丹顶鹤行为比较
以丹顶鹤生存状态和年龄性别为固定因素,各行为双因素方差分析见表1。
由表1可知,野生丹顶鹤与散养丹顶鹤觅食行为、运动行为、警戒行为、其他行为均差异极显著(P<0.01),整理行为差异显著(P<0.05),静栖行为无显著差异(P>0.05)。
对双因素方差所得结果作LSD后期检验,以性别年龄和生存状态作两两比较(见表2)。
由表2可知,觅食行为(野生♂-散养♂P<0.01,野生♀-散养♀P<0.01,野生亚成体-散养亚成体P<0.01)差异均极显著,运动行为只有野生雄性和散养雄性具极显著差异,警戒行为只有野生雌性和散养雌性具极显著差异外,整理行为和其他行为野生和散养丹顶鹤各性别年龄间均差异极显著。
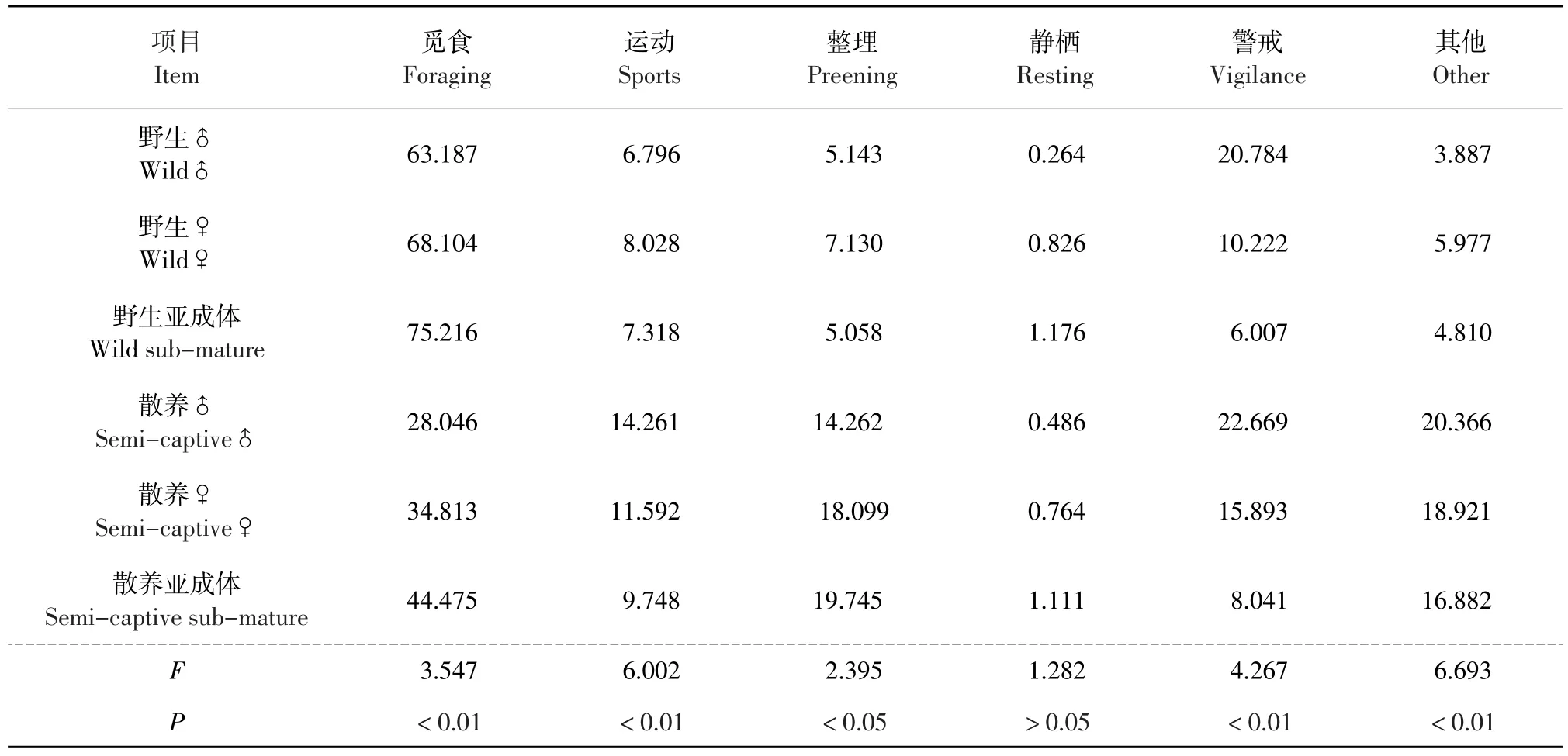
表1 野生丹顶鹤和散养丹顶鹤行为时间分配及双因素方差分析Table 1Time budget of the wild and the semi-captive Grus japonensis and two-way ANOVA
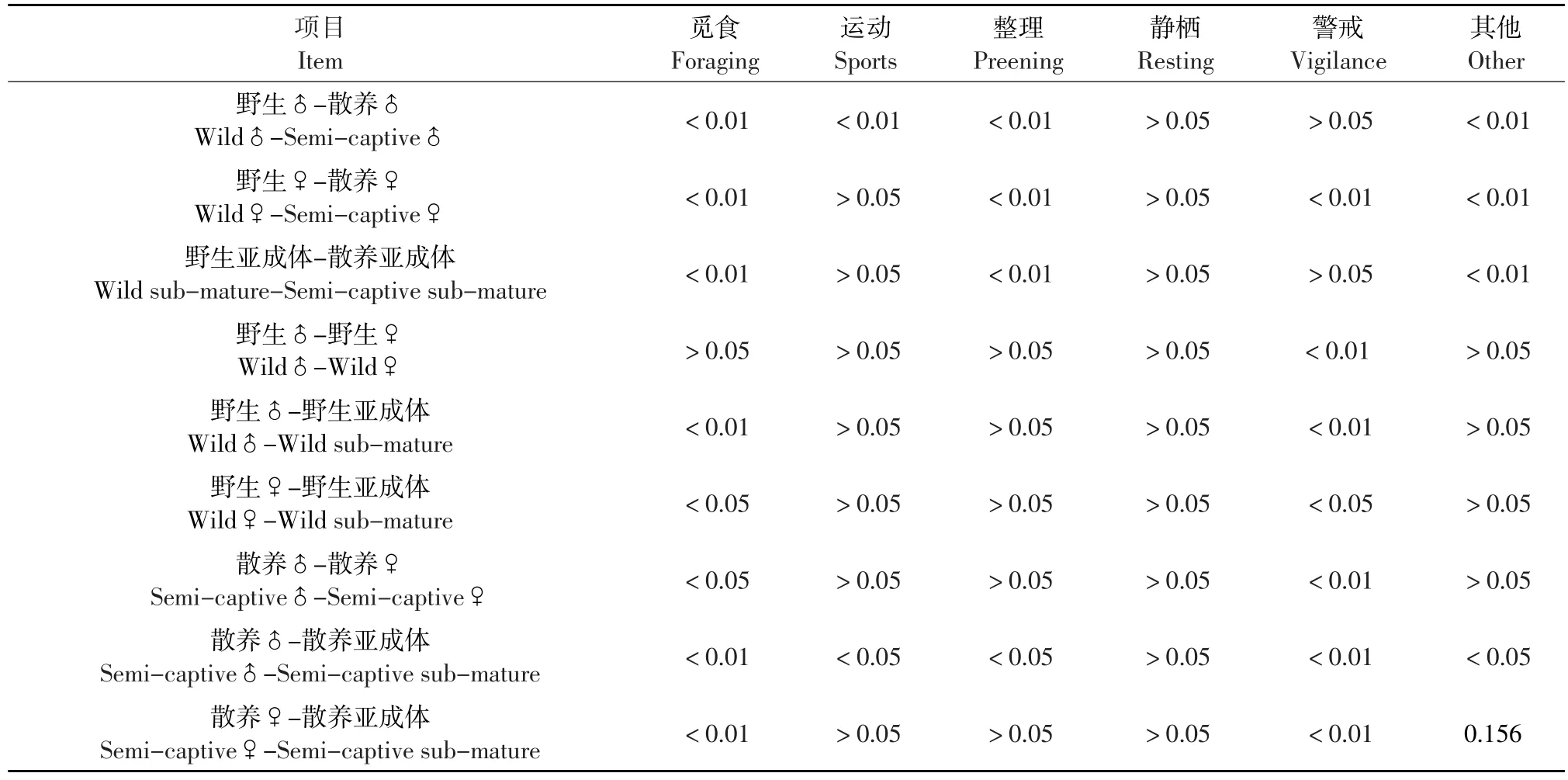
表2 野生丹顶鹤和散养丹顶鹤行为双因素方差分析LSD后期检验Table 2LSD post hoc multiple comparisons of two-way ANVOA of time budget of the wild and the semi-captive Grus japonensis
3 讨论
3.1 野生丹顶鹤与散养丹顶鹤行为差异
野生丹顶鹤在该时期大量觅食,为越冬长途迁徙储存能量,这与中华秋沙鸭越冬迁徙策略很相似[8],因为迁徙飞行行为是高耗能行为方式。而散养丹顶鹤在此期间日行为时间分配未能表现出夏候鸟应有特征,因保护区人工定时定点投喂鲜鱼和鲜玉米,使散养丹顶鹤不用花费更多时间寻找食物。长时间人工投喂模式,使散养丹顶鹤形成对人工投食的依赖,并失去迁徙行为。冬季来临,扎龙保护区湖水河流冻结,部分地区有积雪覆盖,野生丹顶鹤寻找食物困难,则促使其寻找有利环境维持生存。因而,食物缺乏成为丹顶鹤迁徙的主要原因之一。
警戒行为同性别年龄组间比较,发现野生丹顶鹤要比散养丹顶鹤低,原因是散养丹顶鹤所处环境,有游客、饲养员、摄影爱好者等,人为干扰因素较多。野生丹顶鹤生活环境远离人类活动频繁区域,对野生丹顶鹤造成的影响较少。与警戒行为相联系的其他行为(驱赶、鸣叫)的时间分配,野生丹顶鹤较散养丹顶鹤少很多,并表现极显著差异(见表1、2)。
野生丹顶鹤在越冬地越冬期间喜集群生活,迁徙前集群是夏候鸟迁徙的标志[9],本试验期内,未发现野生丹顶鹤集群迹象,活动均以家庭组为单位,家庭组之间距离很远。此间白枕鹤、东方白鹳、白头鹤等夏候鸟种群已开始集群,白枕鹤以4~5个家庭组组成一个群体,在野生丹顶鹤活动区域觅食、静栖。说明扎龙保护区野生丹顶鹤迁徙前集群条件与其他夏候鸟有差异。另外,因观察方法和技术局限、湿地环境复杂性和鸟类活动随机性等因素影响,未能及时观察到野生丹顶鹤集群,也未见扎龙保护区野生丹顶鹤秋季迁徙的相关报道。冬季临近时,扎龙保护区气温明显降低,在固定观察点,仍有野生丹顶鹤出没。说明扎龙保护区野生丹顶鹤迁徙时间可能相对其他夏候鸟较晚,集群迅速,并在短时间内便开始迁徙。散养丹顶鹤该时期活动也是以家庭组为单位,具有领域行为。但不喜欢集群,表现为当散养丹顶鹤同时出现在活动领域交错区内时,则会出现鸣叫和驱赶行为,以示意领域范围。散养丹顶鹤领域行为和其繁殖期领域警戒行为模式相同[10],并将此行为模式延续至越冬期。散养丹顶鹤该行为模式体现出对保护区环境的适应。
关于运动行为,野生丹顶鹤和散养丹顶鹤均会用一定时间飞行。野生丹顶鹤飞行运动一般出现在12:00~13:00和15:00~16:00(见图1、3、5),飞行时间2~3 min,高度20~30 m,由雄性丹顶鹤领头,雌性丹顶鹤和子鹤跟随,排成斜“一”字或“人”字[8],雄性丹顶鹤边飞边鸣叫。散养丹顶鹤也有飞行运动,分别发生在6:00~7:00,10:00~11:00,15:00~16:00(见图1、3、5),飞行时间1~2 min,高度5~15 m,也是由雄鹤领头,雌鹤和子鹤跟随其后,也是排列成斜“一”字或“人”字。野生丹顶鹤飞行是为长途迁徙做准备,飞行多以乘风滑行姿势居多,研究表明滑行姿势飞行和飞行排列的队形,均可有效减少迁徙飞行所耗能量[11]。散养丹顶鹤飞行也有类似滑行姿势和排列队形,说明散养丹顶鹤具备长途迁徙飞行能力,而散养丹顶鹤飞行多用于领土巡视和规避游客骚扰,或对领地里其他鹤进行驱赶,因此,运动行为差异也可大致区分野生丹顶鹤和散养丹顶鹤。
3.2 丹顶鹤个体间行为差异
从表1家庭组中个体间行为比较发现,不论是野生丹顶鹤还是散养丹顶鹤,雄性觅食行为时间比同家庭组其他成员要少,而警戒行为时间高于家庭组其他成员;雌性整理行为时间多于雄性;子鹤一直跟随雌鹤,觅食行为为主要行为。这些差异显示丹顶鹤个体在家庭组中职责不同,雄性丹顶鹤主要起警戒保护作用,雌性丹顶鹤主要负责照顾亚成体鹤,子鹤则在父母的保护和照顾下迅速生长发育。
丹顶鹤行为研究,涉及到雌雄个体间行为比较[12-15],但尚无明确的雌雄丹顶鹤区分标准。对丹顶鹤雌雄识别,从形态上很难区分,可用分子生物学方法[16],该方法非常准确,适用于各年龄段各种群,但过程繁琐、取样困难。从动物行为学角度,也能准确、快速区分丹顶鹤性别。通过本试验研究,从野生和散养丹顶鹤家庭组个体间行为比较中,发现两者间存在显著差异,因而,可辨别雌雄丹顶鹤,但此方法只针对已配对丹顶鹤种群,对亚成体鹤和幼鹤区分效果不明显。
3.3 保护管理建议
本试验中野生与散养丹顶鹤生活环境相似,但行为具显著差异。扎龙保护区人工饲养丹顶鹤,通过驯化而散放,使其适应自然环境变化,繁衍生息,逐渐摆脱人工饲喂依赖而融入野生种群,补充野生丹顶鹤种群数量,达到扩大野生迁徙种群目的。但人工野化放飞成功率不高,而散养雄性丹顶鹤与野生雌性丹顶鹤配对,雌鹤变成扎龙保护区繁殖地不迁徙种群成员的情况时有发生,说明雄性丹顶鹤在已配对家庭组中占有主导地位。散养雌性丹顶鹤与野生雄性丹顶鹤配对成功,人工散养雌性丹顶鹤是否跟随野生雄性丹顶鹤迁徙越冬,春季再迁回繁殖种群,补充野生丹顶鹤种群数量,仍待进一步调查研究。
对保护区散养丹顶鹤管理,可适当逐渐减少直至停止投食,改变其人工投食依赖,使其能够自行觅食;更改投食点,使散养丹顶鹤觅食地点远离人类活动频繁区域,改变散养丹顶鹤生活模式并逐步向野生生活模式过渡,使散养丹顶鹤野化成为可能。
[1]Hiroki T,Yukari T,Moe K,et al.Changes of mercury contamina⁃tion in red-crowned cranes,Grus japonensis,in East Hokkaido,Ja⁃pan[J].Arch Environ Contam Toxicol,2012,63:153-160.
[2]Su L Y,Zou H F.Status,threats and conservation needs for the continental population of the red-crowned grane[J].Chinese Bird, 2012,3(3):147-164.
[3]赵越,李晓民,王令刚.中国丹顶鹤研究现状及保护进展[J],野生动物,2013,34(6):358-360.
[4]Nielsen E T.Relation of behavioral activity rhythms to the chang⁃es of day and night,a revisionof views[J].Behaviour,1984,89(3): 147-173.
[5]周晓禹,王晓明,姜振华.贺兰山石鸡越冬期昼间行为时间分配及活动规律[J].东北林业大学学报,2008,36(5):45-46.
[6]Belovshy G E,Slade J B.Time budget of grassland herbivres,body siaze similarities[J],Oecologia(Berlin).1986,70(1):53-62.
[7]赵旭.扎龙国家级自然保护区水环境评价与实地资源可持续发展研究[J].水资源研究,2006.27(3):22-25.
[8]易国栋,杨志杰,刘宇,等.中华秋沙鸭越冬行为时间分配及日活动节律[J].生态学报,2010,30(8):222-223.
[9]吕士成,陈浩,刘中权.越冬期丹顶鹤集群行为研究[J].中国生物圈保护区,1996(6):6-9.
[10]赵越,李淑玲.扎龙保护区人为干扰对繁殖期丹顶鹤的行为影响[J].东北农业大学学报,2012.43(6):92-95.
[11]吕士成,陈卫华.环境因素对丹顶鹤越冬行为的影响[J].野生动物,2006(6):18-20.
[12]马逸清,李晓民.丹顶鹤研究[M].上海:上海科技教育出版社,2002. [13]仇褔臣,刘胜龙,蔡勇军.散养丹顶鹤的繁殖行为[J].东北林业大学学报,2002,30(9):81-82.
[14]田秀华,石全华,吕士成,等.半散放丹顶鹤越冬日活动行为[J].生态学杂志,2006,25(9):1077-1081.
[15]计子娟,李淑玲,马建华,等.散养与野生丹顶鹤孵卵期行为比较[J].动物学杂志,2008,43(6):62-67.
[16]张红霞,郑维平,包文斌,等.丹顶鹤(Grus japonensis)性别鉴定的分子标记方法[J].农业生物技术学报,2008,16(3):417-420.
LI Shuling1,MA Jie1,WANG Wenfeng2, DU Wei2(1.School of Life Sciences,Northeast Agricultural University,Harbin 150030,China;2. Heilongjiang Zhalong National Nature Reserve,Qiqihar Heilongjiang 161000,China)
The activity rhythm and the time budget in fall-migrating season of the wild and the semi-captiveGrus japonensiswere studied by using instantaneous scan method on Zhalong Reserve.The results showed that the time distribution of foraging behaviour of the wildGrus japonensiswas more than the semi-captive,the preending and the other behaviour(banishing and screaming)of the semi-captiveGrus japonensiswere higher than the wild;the time budget of behaviour of male and female adults had some differences.The research was found that the wild cranes had a lot of foraging,was the long distance migration by north to south,reserve energy;the semi-captive,as a not migratory species,time distribution of preening and other behaviours,showed the adaptability to the environment of the reserve.The wild and the semi-captiveGrus japonensisbehavioural differences between individuals in the family group,which indicated that the status ofGrus japonensisin the family group,and bore the responsibility.From the wild and the semi-captiveGrus japonensisin the fall-migrating season rhythm observation of behaviour,set upbehaviour spectrum,the wild and the semi-captiveGrus japonensisbehaviour differences,thus,we can improve the technique of artificial breeding and theGrus japonensissurvival environment,as well as improve the crane behaviour research to provide new information.
Zhalong Reserve;daily activity rhyhm;Grus japonensis;adaptability;behaviour spectrum
Q959.7+26
A
1005-9369(2015)07-0062-08
时间2015-7-9 14:42:33[URL]http://www.cnki.net/kcms/detail/23.1391.S.20150709.1442.005.html
李淑玲,马杰,王文锋,等.扎龙保护区丹顶鹤秋季迁徙季节行为节律的研究[J].东北农业大学学报,2015,46(7)∶62-69.
Li Shuling,Ma Jie,Wang Wenfeng,et al.Behavoiral rhythm research of theGrus japonensison Zhalong Reserve during the fall-migrating season[J].Journal of Northeast Agricultural University,2015,46(7)∶62-69.(in Chinese with English abstract)
2014-12-16
黑龙江省自然科学基金资助项目(C200921)
李淑玲(1965-),女,教授,博士,研究方向为动物行为与繁殖生态。E-mail:hljshulingli@163.com
Behavoiral rhythm research of theGrus japonensison Zhalong Reserve during the fall-migrating season
猜你喜欢
伙伴(2020年4期)2020-05-11
小哥白尼(军事科学)(2018年9期)2018-12-08
小哥白尼(军事科学)(2018年8期)2018-09-12
小哥白尼(军事科学)(2018年6期)2018-09-10
读者(2018年14期)2018-06-28
小哥白尼(军事科学)(2018年1期)2018-05-25
青年文学家(2017年28期)2017-11-28
青年文学家(2016年34期)2017-03-31
学苑创造·A版(2016年10期)2016-11-19
环境与生活(2016年6期)2016-02-27
