
Wx基因对灌浆期小麦叶片POD活性的影响
2015-10-13 21:08孔治有等
湖北农业科学 2015年18期
关键词:小麦
孔治有等


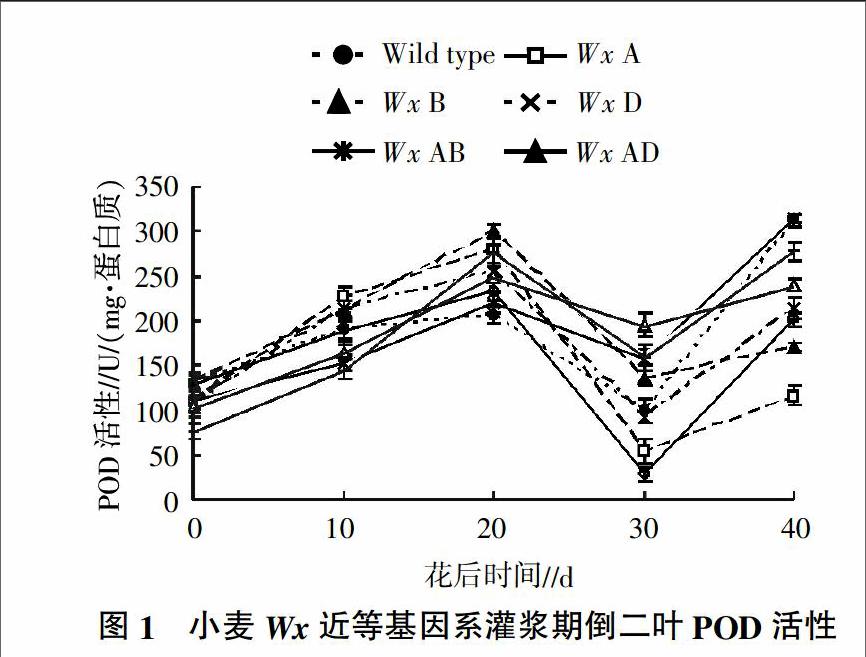
摘要:通过对8个Wx近等基因系灌浆期间旗叶和倒二叶POD活性进行测定,研究Wx基因对小麦(Triticum aestivum Linn.)叶片POD活性的影响,结果表明,Wx-B1基因的缺失对小麦叶片POD活性有增加作用,而Wx-A1和Wx-D1基因的缺失作用则相反,表明WxB型小麦可能对逆境的抗性最强;POD活性随灌浆时间的延长先增加(到花后20 d达到峰值)后降低、最后又增加,同时期旗叶的POD活性显著高于倒二叶,反映了植株叶片的生理状态及活性氧的清除能力强弱。
关键词:小麦(Triticum aestivum Linn.);Wx近等基因系;POD活性;灌浆期
中图分类号:S512.01 文献标识码:A 文章编号:0439-8114(2015)18-4401-03
DOI:10.14088/j.cnki.issn0439-8114.2015.18.005
植物生长过程中会产生活性氧类(AOS),过量的AOS则会对植物造成氧化胁迫,使细胞产生细胞水平和分子水平上的不可逆损伤[1]。植物体中最重要的抗氧化系统是抗氧化酶类,包括抗坏血酸过氧化物酶(APX)、谷胱甘肽还原酶(GR)、过氧化氢酶(CAT)及超氧化物歧化酶(SOD)等。过氧化物酶(Peroxidase,POD)是广泛存在于各种动、植物和微生物体内的一类氧化酶,催化由过氧化氢参与的各种还原剂的氧化反应:RH2+H2O2→2H2O+R。根据催化底物特性可将POD分为愈创木酚POD、谷胱甘肽POD和抗坏血酸POD等[2]。POD作为植物细胞内重要的组成部分,具有许多非常重要的生理功能,如参与活性氧代谢、木质素和木栓层的形成、生长素的降解以及其他(如谷胱甘肽、NADH、DTT、草酰乙酸、氢醌、酪氨酸)酚类化合物的氧化过程。除了与植物正常代谢和生长发育相关的结构型POD外,很大部分POD的合成属于诱导表达型,这已为大量的胁迫处理试验和在分子水平上的基因调控研究证据证实。在上述的POD生理功能中,多数与某种或多种胁迫作用导致细胞膜的损伤和破坏,细胞的空间结构被打破,以及损伤信号的转导等一系列生理生化变化有关[3]。陈立松等[4]在对荔枝进行的水分胁迫试验中发现,随着水分胁迫程度的增加,荔枝叶片中POD活性增加,而且抗旱性较强的品种上升的幅度大于抗旱性较弱的品种。吴伯千等[5]发现茶树遭受水分胁迫时,水分胁迫过程有一个临界强度,耐旱性强的茶树品种其POD活性在一定范围内相对较低,超过这一范围则维持相对较高的活性水平。
六倍体小麦中的Waxy蛋白质受3个不同的Wx基因编码,分别位于染色体臂7AS(Wx-A1)、4AL(Wx-B1)和7DS(Wx-D1)上[6-9],Wx基因的缺失、突变或遗传表达障碍会使胚乳中直链淀粉的含量减少和支链淀粉含量增加,全部缺失的小麦胚乳表现为糯性而不含直链淀粉[10-15]。关于Wx基因对小麦粉品质及食品品质的影响已有较多研究,但目前尚无Wx基因对其他生理生化特性的影响的报道。本研究以小麦Wx近等基因系为材料,探讨Wx基因对小麦灌浆期叶片POD活性的影响,以期为将来小麦Wx基因的深入研究提供参考。
1 材料与方法
1.1 材料
8个小麦Wx近等基因系(宁麦14背景)种植于云南农业大学校内试验基地(表1),于花后每隔10 d取样测定叶片POD活性。
1.2 方法
从开花后第0、10、20、30和40 d取样,每个基因型每次取5个单株(即5次重复)的旗叶和倒二叶用于测定。POD活性采用愈创木酚法[16]进行测定。每个基因型重复5次,以SAS9.0进行统计分析。
2 结果与分析
在叶位、基因型和取样时间3个处理因素中,除基因型与取样时间互作外,POD活性受任何单一因素或两因素互作的影响,均达到了极显著水平(表2)。
8个基因型中,Wx B的POD活性最高,Wx ABD次之,以Wx A、Wx D、Wx BD和Wx AB最低(表3),Wx B和Wx ABD与Wx A、Wx D和Wx AB的POD活性达到显著或极显著水平。
在各取样时间中,POD活性以花后20 d最高,花后40 d次之,初开花时(花后0 d)最低(表4),相互之间差异达极显著水平(花后10 d与花后30 d之间差异未达极显著水平)。
随灌浆时间的延长,8个Wx基因型倒二叶POD活性先逐渐增加,到花后20 d达到最大,此后急剧降低,在花后30 d达最低值,然后又迅速升高(图1)。尽管所有基因型的POD活性变化趋势一致,但活性水平有较大差异,其中WxB在花后前20 d的POD活性最大,但此后降低幅度较大,回升后也只处于中等水平;Wild type在灌浆开始阶段POD活性相对不高,但灌浆结束时增加较快,高于其他基因型。
随灌浆时间的延长,各Wx基因型旗叶POD活性增加变化趋势与倒二叶基本一致,开花后即急剧增加,在花后20 d达到最大值,此后迅速降低,但自降低后POD活性增加幅度极小,基本处于平稳状态(图2)。在所有基因型中,WxB初开花时POD活性不高,但增加迅速,到花后10 d起即高于其他基因型;Wild type在初开花时活性最高,在花后10 d后仅次于WxB,花后30 d POD活性降低又高于其他基因型。
3 小结与讨论
Wx-B1基因的缺失对小麦叶片POD活性有增加作用,而Wx-A1和Wx-D1基因的缺失作用则相反。POD活性随灌浆时间的延长先增加(到花后20 d达到峰值)后降低、最后又增加。同时期旗叶的POD活性显著高于倒二叶。
植物体内会不断产生并清除活性氧,为防止自由基的伤害,植物体会产生一些活性物质以维持正常的生理功能,POD就是其中一种清除活性氧的保护酶。POD以过氧化氢为电子受体催化底物氧化,主要是将H2O2水解,从而对细胞起保护作用[17]。POD活性的强弱反映了植物抗逆境及衰老的能力。
本试验中,叶位、基因型和取样时间均对POD活性产生显著影响,其中Wx-B1基因的缺失可提高POD活性水平,而缺失Wx-A1和Wx-D1基因则会降低叶片POD活性。在8个近等基因系中,Wx B型小麦对逆境的抗性最好。
在整个灌浆期中,花后20 d所有基因型的POD活性均达到最高,此时灌浆速度最快、可能产生的活性氧也随之增多,从而导致清除活性氧的酶包括POD活性也随之增强;此后POD活性逐渐降低,与灌浆活动逐渐减弱有关;而在灌浆的30~40 d时POD活性有明显增加趋势,则可能是叶片加速衰老导致活性氧急剧增加引起的清除系统应急性增强。
旗叶POD活性在花后各阶段均高于倒二叶,表明旗叶在整个灌浆过程中的生理活动均强于倒二叶,内部产生的活性氧也相对更多、导致清除系统的作用也相对更强;旗叶POD活性与倒二叶在灌浆过程中的变化基本一致,惟一区别在于旗叶POD活性在花后30~40 d增加很少或基本保持稳定,表明此时旗叶功能仍保持一定的活力,而倒二叶的生理功能相对更为衰弱。
参考文献:
[1] 秦小琼,贾士荣.植物抗氧化逆境的基因工程(综述)[J].农业生物技术学报,1997,5(1):14-21.
[2] ASADA K.Ascorbate peroxidase-a hydrogen peroxide-scavenging enzyme in plants[J].Physiol Plant,1992,85:235-241.
[3] 田国忠,李怀方,裘维蕃.植物过氧化物酶研究进展[J].武汉植物学研究,2001,19(4):332-344.
[4] 陈立松,刘星辉.水分胁迫下荔枝叶片过氧化物酶和IAA氧化酶活性的变化[J].武汉植物学研究,2002,20(2):131-136.
[5] 吴伯千,潘根生.茶树对水分胁迫的生理生化反应[J].浙江农业大学学报,1995,21(5):451-456.
[6] ANISWORTH C,TARVISM,CLARK J. Isolation and analysis of cDNA clone encoding the small subunit of ADP-glucose pyrophosphorylase from wheat[J].Plant Molecular Biology,1993, 23: 23-33.
[7] NAKAMURA T,YAMAMORI M,HIRANO H,et al. Identification of three Wx proteins in wheat(Triticum aestivum L.)[J].Biochemical Genetics,1993,31(1-2):75-86.
[8] YAMAMORI M,ENDO T R.Variation of starch granule proteins and chromosome mapping of their coding genes in common wheat[J]. Theor Appl Genet,1996,93:275-281.
[9] SUN C, PUTHIGAE S,STAFFAn A,et al.The two genes encoding starch-branching enzymes Ⅱa and Ⅱb are differentially expressed in barley[J].Plant Physiology,1998,118:37-49.
[10] 陈新民.糯小麦(Waxy Wheat)研究进展[J].麦类作物学报,2000,20(3):82-85.
[11] 邓万洪,晏本菊,任正隆.糯性小麦研究进展[J].麦类作物学报,2007,27(1):166-171.
[12] 刘爱峰,宋建民,赵振东,等.糯小麦配粉对面团流变学特性和面包烘烤品质的影响[J].中国农业科学,2004,37(6):902-907.
[13] 李 春,宋广芝,田纪春.糯小麦及其Waxy基因的研究进展[J].中国农学通报,2007,23(7):257-262.
[14] 梁荣奇,张义荣,刘广田,等.小麦淀粉品质改良的综合标记辅助选择体系的建立[J].中国农业科学,2002,35(3):245-249.
[15] 孙彩玲,田纪春,翟红梅,等.糯性小麦育种的早期选择[J].山东农业科学,2006(3):15-17.
[16] 覃 鹏.转基因SOD高表达烟草抗旱性研究[D].昆明:云南大学,2003.
[17] 邵世光,阎斌伦,许云华,等.Cd2+对条斑紫菜的胁迫作用[J].河南师范大学学报(自然科学版),2006,34(2):113-l16.
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2021年12期)2021-11-28
今日农业(2021年8期)2021-11-28
今日农业(2021年8期)2021-11-28
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
河北农机(2020年10期)2020-12-14
今日农业(2020年13期)2020-08-24
作文小学中年级(2020年4期)2020-06-11
今日农业(2019年11期)2019-08-13
