
湖南省部分地区棉田牛筋草(Eleusineindica)对精喹禾灵的抗性
2015-11-25 08:31刘祥英柏连阳
植物保护 2015年2期
宗 涛, 李 洁, 刘祥英, 柏连阳
(1.湖南农业大学植物保护学院, 长沙 410128; 2. 湖南省农业科学院, 长沙 410125)
湖南省部分地区棉田牛筋草(Eleusineindica)对精喹禾灵的抗性
宗 涛1, 李 洁1, 刘祥英1, 柏连阳2*
(1.湖南农业大学植物保护学院, 长沙 410128; 2. 湖南省农业科学院, 长沙 410125)
采用整株法测定了湖南省部分地区棉田牛筋草(Eleusineindica)对精喹禾灵的抗性水平,还测定了精喹禾灵对牛筋草抗(R)、感(S)种群体内GSTs及SOD、POD、CAT影响的差异。整株法测定结果显示,相对于敏感的南县种群,其他地区棉田牛筋草对精喹禾灵的抗性指数在1.6~9.7之间,其中湘阴种群最高为9.7,澧县种群最低为1.6;精喹禾灵处理2 d后,R种群GSTs活性急剧上升,在第3天达到最高,随后急剧下降,5 d之后趋于平缓。S种群GSTs活性在施药后先上升后下降。药剂处理后,R种群的SOD和POD活性在中期显著低于S种群,R、S种群的CAT活性在药后第9天有明显差异。结果表明,R种群对精喹禾灵产生抗性的原因可能是GSTs对精喹禾灵的代谢作用加强,抗氧化酶系活性的变化可能与抗性相关。
牛筋草; 精喹禾灵; 抗药性
ACCase抑制剂类药剂的作用位点单一,抗药性产生较快。自第一例抗ACCase抑制剂类除草剂禾草灵(diclofop-methyl)的瑞士黑麦草(Loliumrigidum)在澳大利亚出现后[3],抗ACCase抑制剂类除草剂的杂草先后在英国、澳大利亚、德国、法国和瑞士等国发现,至今已在36个国家发现46种抗ACCase制剂类除草剂的杂草生物型[4]。国外关于牛筋草对ACCase抑制剂类的抗性已有一些报道。Leach等报道了牛筋草对ACCase抑制剂类除草剂产生抗性是由靶标酶位点突变产生[5]。Vidal等报道了巴西大豆田的牛筋草对ACCase抑制剂类除草剂产生了抗性[6]。Osuna等报道了牛筋草对ACCase抑制剂类除草剂产生抗性,并发现了一个突变位点[7]。我国已报道日本看麦娘(AlopecurusjaponicasSteud.)、棒头草(PolypogonfugaxNeesexSteud.)、菵草[Beckmanniasyzigachne(Steud.)Fernald]对 ACCase抑制剂类产生抗性[8-10],但牛筋草对该类除草剂的抗性仍然未见报道。
靶标抗性和非靶标抗性是杂草对除草剂产生抗药性的两大类。非靶标抗性大多抗性水平不高,是指植物通过提高对除草剂的轭和作用、降解作用以及清除除草剂产生的有毒代谢产物的解毒能力,从而对除草剂产生的抗性[11]。谷胱甘肽S-转移酶(glutathione s-transferase,GSTs)是杂草对除草剂通过解毒作用产生非靶标抗性的关键酶,对于大部分除草剂在杂草体内的代谢有着重大作用[12]。研究显示双子叶杂草苘麻(Abutilontheophrasti)对莠去津(atrazine)的抗性和小子草(Phalarisminor)对异丙隆的抗性均是由于增强代谢导致[13]。Hall等研究表明英国种群的鼠尾看麦娘(Alopecurusmyosuroides)对精噁唑禾草灵(fenoxaprop-P-ethyl)和禾草灵(diclofop-methyl)的抗性,在一定程度上是鼠尾看麦娘对这两种除草剂的代谢增强所致[14]。除草剂是一种非生物逆境,植物体内的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)可以降低干旱、低温、病、虫、盐碱及涝害等逆境对植物造成的危害[15-17]。Pyon 等研究结果显示,抗氧化酶系活性提高和还原型谷胱甘肽含量升高可能是小飞蓬(Erigeroncanadensis)对草甘膦产生抗性的原因之一[18]。但Zabalza 等研究表明氧化胁迫作用与豌豆(PisumsativumLinn.)对乙酰乳酸合成酶(ALS)抑制剂类除草剂产生的耐药性无关[19]。本文以抗性及敏感牛筋草为试材测定SOD、POD、CAT活性,为明确牛筋草抗性与氧化胁迫之间的关系,丰富杂草抗性理论,以及牛筋草的抗性治理提供科学依据。
本研究中试验材料采自湖南省洞庭湖周边的益阳、岳阳、常德三大棉区,试验选用整株测定法,检测了牛筋草对精喹禾灵的抗性水平,通过研究施药后代谢酶(GSTs)、抗氧化酶[超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)]的变化,首次探讨了牛筋草对精喹禾灵的抗性机制。
1 材料和方法
1.1 供试材料
供试药剂:50g/L精喹禾灵(quizalofop-P-ethyl)乳油,日本日产化学工业株式会社;硝酸钾(KNO3),国药集团化学试剂有限公司;还原型谷胱甘肽(GSH),Solarbio公司;聚乙烯吡咯烷酮(PVP-40),Sigma公司。
供试种子:2012年8月从湖南省部分地区连续使用精喹禾灵6~10年左右的棉田,采集到10个牛筋草种群。在南县未施过药剂的荒山中采集到相对敏感牛筋草种群。采集地点见表1。
丝黑穗病:播前种子处理,用药剂处理种子是综合防治中不可忽视的重要环节。方法有拌种、浸种和种衣剂处理三种。药剂防治必须选择内吸性强、残效期长的农药,三唑类杀菌剂拌种防治玉米丝黑穗病效果较好,大面积防效可稳定在60%~70%。
试验仪器:3WPSH-500D型生测喷雾塔,农业部南京农业机械化研究所;DK-98-IIA电热恒温水浴锅,天津泰斯特仪器有限公司;UVmini-1240型紫外分光光度计,岛津国际贸易(上海)有限公司;X-22R高速冷冻离心机,贝克曼库尔特商贸(中国)有限公司。
1.2 试验方法
1.2.1 整株测定法
取破除颖壳后的牛筋草种子,用0.125%的KNO3溶液浸泡24 h。在面积为100 cm2的塑料盆中,装入泥土与营养土2∶1混配的土壤,浇灌2次,使土壤完全湿润。将种子均匀撒到盆中,每盆约30粒,覆盖一层薄砂,再次浇水,使牛筋草种子与土壤充分接触,置于室外培养。2~3叶期时对牛筋草进行间苗,选取长势一致的牛筋草15株。于牛筋草4~5叶期,使用3WPSH-500D型生测喷雾塔进行茎叶喷雾。精喹禾灵有效剂量设置为0、3.75、7.5、15、30、60 g/hm2,所有梯度均设3个重复。施药后14 d,称量牛筋草地上部分鲜重,采用毒力回归方程y=a+bx,经SPSS(19.0)软件得出GR50。y为特定除草剂用量下所测杂草的抑制率的几率值,x为除草剂用量的对数,GR50为除草剂对牛筋草的生长抑制中量。敏感种群选用南县牛筋草,根据公式计算得出各地区的抗药性指数(resistance index,RI)。

取南县牛筋草为敏感(S)种群,湘阴牛筋草为抗性(R)种群。于牛筋草的4~5叶期,用133.33 g/hm2的精喹禾灵进行茎叶喷雾,设清水为对照。施药后第1、2、3、5、7天剪取牛筋草地上部分。试验参照吴进才等的方法改进[20],称取样品0.5 g,置于预冷的研钵中,加入8 mL 0.1 mol/L的Tris-HCl缓冲液(含还原型谷胱甘肽25 mmol/L,5%PVP,pH 8.0)后冰浴研磨至匀浆,于4 000 r/min离心15 min。取上清液后,于10 000 r/min再离心10 min,得到上清液作为酶提取液(上述操作在-4℃下进行)。试验参照吴进才[20]、韩瑞娟[21]等的方法,在3 mL 0.1 mol/L的Tris-HCl缓冲液中(pH 8.0)中加入0.1 mL酶液,于水浴锅中25℃恒温10 min,再加入13 mmol/L CDNB(由0.1 mL无水乙醇配制),反应10 min后,置于分光光度计340 nm处测得A值。用摩尔消光系数9.5 mmol/(L·cm)来计算样品的蛋白活性。蛋白含量测定采用考马斯亮蓝G-250法。试验平行测定3次。
蛋白活性=1 000*A/(C*9.5)
其中A为吸光度值,C为稀释后样品的蛋白浓度mg/mL,蛋白活性单位为nmol/(min·mg)。
GSTs的相对活性为处理样品的GSTs活性与同期CK样品活性的比值。
1.2.3 超氧化物歧化酶、过氧化物酶、过氧化氢活性测定
取南县牛筋草为敏感(S)种群,湘阴牛筋草为抗性(R)种群。于牛筋草的4~5叶期,用133.33 g/hm2的精喹禾灵进行茎叶喷雾,设清水为对照。施药后第1、5、9、14天剪取牛筋草地上部分。参照Wang等、Aebi的方法[22-24]。称取牛筋草地上部分0.5 g,剪碎放入预冷的研钵中,加入5 mL 50 mmol/L预冷的磷酸缓冲液(PBS,pH 7.8)冰浴匀浆,离心20 min(4 ℃,12 000 r/min),取上清液置于-80℃冰箱待用。采用南京建成生物工程研究所提供的试剂盒进行蛋白含量和酶活性测定。
1.3 数据处理
数据处理选用软件SPSS(19.0)。得出不同地区牛筋草种群对精喹禾灵的线性回归方程、GR50、95%置信限及相关系数r。作图软件使用Sigmaplot(10.0)。
2 结果与分析
2.1 整株测定法
结果见表1。对照为南县的敏感牛筋草种群,各地区牛筋草种群均已对精喹禾灵产生不同程度的抗性,抗性指数在1.6~9.7之间。其中湘阴种群的抗性水平最高,其次是汉寿、大通湖、沅江种群。
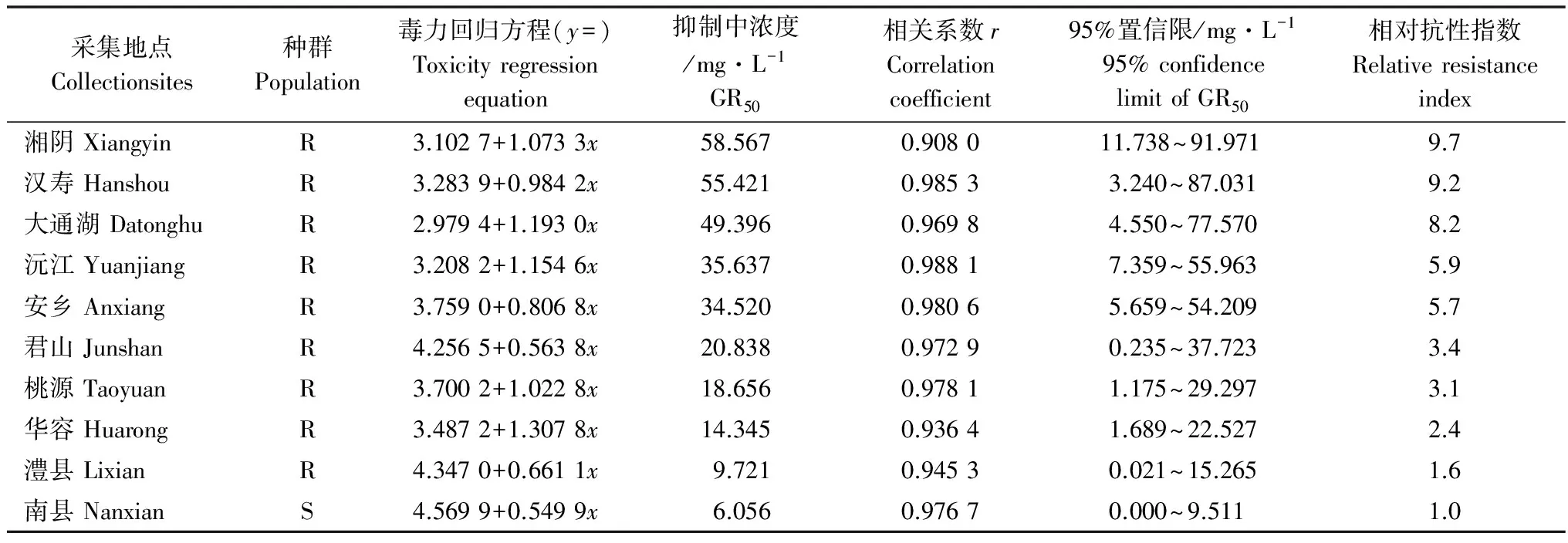
表1 不同棉田牛筋草种群对精喹禾灵的抗性水平1)
1) S为敏感种群,R为潜在抗药性种群;抗药性指数(RI)=GR50(R)/GR50(S)。
S, susceptible population; R, potential resistant population; Resistance index(RI)=GR50(R)/GR50(S)
2.2 精喹禾灵对敏感及抗性种群牛筋草GSTs活性的影响
抗性牛筋草种群在药剂处理前的GSTs活性为121.31 nmol/(min·mg),敏感牛筋草种群则为74.18 nmol/(min·mg),抗性种群GSTs活性高于敏感种群。图1中相对活性结果均为各天施药后GSTs活力/抗性或敏感种群空白对照GSTs活力。结果显示,精喹禾灵133.33 g/hm2处理2 d后,抗性牛筋草种群GSTs活性急剧上升,在第3天达到最高,随后急剧下降,5 d之后趋于平缓。敏感种群GSTs活性在施药后1 d开始上升,2 d之后开始下降,第3、5天基本无变化,随即又上升。抗性种群在第2、3、5、7天的GSTs活性波动较大,变化急剧;敏感种群GSTs活性波动较小,变化相对平缓,说明抗性种群受药剂的影响较大。

图1 抗性及敏感种群茎叶组织GSTs活性变化Fig.1 The activity variation of GSTs in resistant and susceptible plants

图2 抗性及敏感种群茎叶组织SOD活性变化Fig.2 Changes in SOD activity in leaf and shoot extracts of resistant (R) and susceptible (S) Eleusine indica
2.3 精喹禾灵对敏感及抗性种群牛筋草SOD、POD、CAT活性的影响
图2结果显示,用133.33 g/hm2精喹禾灵处理牛筋草后,抗性种群和敏感种群SOD活性变化在14 d内没有显著性差异。但敏感种群的SOD相对活性均高于同期的抗性种群,且在第5、9、14天有显著性差异。
图3结果显示,用133.33 g/hm2精喹禾灵处理牛筋草后,抗性种群和敏感种群POD活性变化在14 d内均没有显著性差异,但抗性种群的POD相对活性均低于同期敏感种群,且在第5、9天有显著性差异。

图3 抗性及敏感种群茎叶组织POD相对活性变化Fig.3 Changes in POD activity in leaf and shoot extracts of resistant (R) and susceptible (S) Eleusine indica
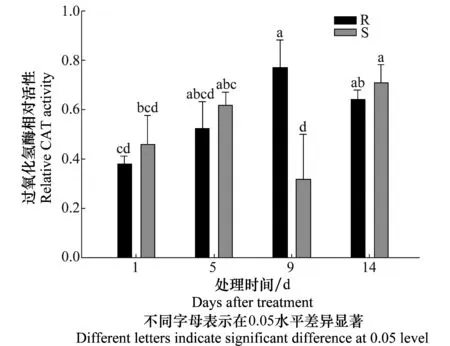
图4 抗性及敏感种群茎叶组织CAT相对活性变化Fig.4 Changes in CAT activity in leaf and shoot extracts of resistant (R) and susceptible (S) Eleusine indica
植物体内的过氧化氢酶可以清除过氧化氢,是植物体内酶促防御系统的重要部分。植物组织中CAT活性和H2O2含量与植物自身的抗逆性紧密关联。CAT试验结果见图4。用精喹禾灵133.33 g/hm2处理牛筋草后,第9天敏感种群CAT活性显著性下降,达到最低,而抗性牛筋草种群CAT活性达到峰值,且与敏感牛筋草种群CAT有显著性差异。敏感种群CAT活性在第14天有显著性上升。第1、5、14天抗性与敏感种群CAT活性没有显著性差异。
3 讨论
Gressel对稻田除草剂抗性风险进行分级,参考了除草剂的作用靶标、选择压、持效期、使用历史等。马晓渊综合国内实际情况,对国内水旱田常用除草剂的抗性风险进行了分级,其中ACCase抑制剂类除草剂属于高风险级别[25],报道一般连续使用6~10年就会产生抗性[26]。整株法测定结果表明:湖南省部分地区棉田牛筋草已对精喹禾灵产生不同程度的抗性,其中,湘阴牛筋草种群抗性水平相对较高,当地使用精喹禾灵防除棉田杂草已将近10年,在长期的选择压力下产生了抗性,这与当地反映采用精喹禾灵防除牛筋草失效的情况相一致。未采取除草剂的混用和轮用措施的情况下,农户若继续增大精喹禾灵用药量和用药次数,则会使该药剂对牛筋草选择压力继续增加,导致抗药性迅速发展,增加抗性风险。要制定合理的化学防除方案,首先必须明确各地棉田牛筋草对除草剂的抗性水平。本研究针对湖南省洞庭湖棉区益阳、岳阳、常德的棉田牛筋草进行了抗药性水平测定,今后应当进一步扩大疑似抗性牛筋草种群的采集区域。
本研究中的湘阴抗性种群牛筋草和南县敏感种群牛筋草的GSTs活性存在较大的差异,施用药剂后,抗性和敏感种群GSTs活性在前期均有显著性上升,但在第2~7天内,抗性种群GSTs相对活性受药剂影响大,有显著性变化,而敏感种群GSTs相对活性变化趋势不明显。试验表明,牛筋草对精喹禾灵的抗药性与GSTs活性密切相关。牛筋草对精喹禾灵产生抗性的重要原因之一就是GSTs对精喹禾灵代谢能力的提高。Western blot研究结果显示看麦娘体内GSTs活性增强与ACCase的轭合使噁唑禾草灵被解毒,从而导致了看麦娘对噁唑禾草灵产生抗性[27]。研究指出在鼠尾看麦娘的敏感生物型中发现了比抗AOPP类除草剂生物型中更低的GSTs活性[27-28]。
许长成等比较了两个水稻品种,敏感品种‘桂朝2号’比抗百草枯品种‘Lemont’比具有较低的抗逆活性[29]。吴进才等研究认为水稻叶鞘中SOD活性的变化反映了水稻植株对除草剂的抵御能力[30]。Choudhary等研究显示受到重金属胁迫的藻青菌(Spirulinaplatensis-S5)SOD活性显著上升,认为是藻青菌的一种自我保护反应[31]。抗性种群与敏感种群的CAT活性差异在第9天达到最大,可能是抗性种群受到药剂胁迫后,启动自身保护系统避免受到伤害。植物抗氧化酶系活性的提高是抵抗逆境的方式之一,而本研究中药剂处理之后,抗性种群SOD相对活性在第5、9、14天显著低于敏感种群,POD相对活性第5、9天显著性低于敏感种群,与前人研究相悖,SOD和POD活性变化对牛筋草抗性的产生是否有影响,还有待于进一步研究。
本研究仅从代谢抗性和抗氧化酶系活性方面研究了抗性产生的机理,但抗性的产生是否涉及编码ACCase的基因的突变还有待于进一步研究。
[1] 李扬汉.中国杂草志[M]. 北京:中国农业出版社, 1998: 1222.
[2] Lee L J, Ngim J. A first report of glyphosate-resistant goosegrass (Eleusineindica(L.) Gaertn.) in Malaysia [J]. Pest Management Science, 2000, 56(4): 336-339.
[3] Heap I M, Knight R. The occurrence of herbicide cross resistance in a population of annual ryegrass,Loliumrigidum, resistant to diclofop-methyl [J]. Australian Journal of Agricultural Research, 1986, 37(2): 149-156.
[4] Heap I. International survey of herbicide resistant weeds[J/OL].(2015-03-09).http:∥www.weedscience.org/,2014.
[5] Leach G E, Devine M D, Kirkwood R C, et al. Target enzyme-based resistance to acetyl-coenzyme A carboxylase inhibitors inEleusineindica[J]. Pesticide Biochemistry and Physiology, 1995, 51(2): 129-136.
[6] Vidal R A, Portes E S, Lamego F P, et al. Resistência deEleusineindicaaos inibidores de ACCase [J]. Planta Daninha, 2006, 24(1): 163-171.
[7] Osuna M D, Goulart I, Vidal R A, et al. Resistance to ACCase inhibitors inEleusineindicafrom Brazil involves a target site mutation [J]. Planta Daninha, 2012, 30(3): 675-681.
[8] Mohamed I A, Li R, You Z, et al. Japanese foxtail (Alopecurusjaponicus) resistance to fenoxaprop and pinoxaden in China [J]. Weed Science, 2012, 60(2): 167-171.
[9] Tang W, Zhou F Y, Chen J, et al. Resistance to ACCase-inhibiting herbicides in an Asia minor bluegrass (Polypogonfugax) population in China [J]. Pesticide Biochemistry and Physiology, 2014, 108: 16-20.
[10]Li Lingxu, Bi Yaling, Liu Weitang, et al. Molecular basis for resistance to fenoxaprop-p-ethyl in American sloughgrass (BeckmanniasyzigachneSteud.) [J]. Pesticide Biochemistry and Physiology, 2013,105:118-121.
[11]李洁, 宗涛, 刘祥英, 等. 湖南省部分地区棉田牛筋草(Eleusineindica)对高效氟吡甲禾灵的抗药性[J]. 棉花学报, 2014, 26(3): 279-282.
[12]苏少泉. 除草剂在植物体内的代谢与选择性及使用[J]. 现代农药, 2003, 2(6): 14-17.
[13]Anderson M P, Gronwald J W. Atrazine resistance in a velvetleaf (Abutilontheophrasti) biotype due to enhanced glutathioneS-transferase activity [J]. Plant Physiology, 1991, 96(1): 104-109.
[14]Hall L M, Moss S R, Powles S B. Mechanisms of resistance to aryloxyphenoxypropionate herbicides in two resistant biotypes ofAlopecurusmyosuroides(blackgrass): herbicide metabolism as a cross-resistance mechanism [J]. Pesticide Biochemistry and Physiology, 1997, 57(2): 87-98.
[15]张朝贤, 倪汉文, 魏守辉, 等. 杂草抗药性研究进展[J]. 中国农业科学, 2009, 42(4): 1274-1289.
[16]Powles S B, Yu Q. Evolution in action:plants resistant to herbicides [J].Annual Review of Plant Biology, 2010, 61: 317-347.
[17]Choudhary M, Jetley U K, Abash K M, et al. Effect of heavy metal stress on proline, malondialdehyde, and superoxide dismutase activity in the cyanobacteriumSpirulinaplatensis-S5 [J]. Ecotoxicology and Environmental Safety, 2007, 66 (2): 204-209.
[18]Pyon, J Y, Piao R Z, Roh S W, et al. Differential levels of antioxidants in paraquat-resistant and susceptibleErigeroncanadensisbiotypes in Korea [J]. Weed Biology and Management, 2004, 4: 75-80.
[19]Zabalza A, Gaston S, Sandalio L M, et al. Oxidative stress is not related to the mode of action of herbicides that inhibit acetolactate synthase [J]. Environmental and Experimental Botany, 2007, 59 (2): 150-159.
[20]吴进才, 许俊峰, 冯绪猛, 等. 稻田常用农药对水稻3个品种生理生化的影响[J]. 中国农业科学, 2003, 36(5): 536-541.
[21]韩瑞娟.日本看麦娘对高效氟吡甲禾灵抗性机理的研究[D].南京:南京农业大学,2010.
[22]Wang S H, Yang Z M, Yang H, et al. Copper-induced stress and antioxidative responses in roots ofBrassicajunceaL. [J]. Botanical Bulletin of Academia Sinica, 2004, 45(3):203-212.
[23]Wang Yousheng, Yang Zhimin. Nitric oxide reduces aluminum toxicity by preventing oxidative stress in the roots ofCassiatoraL. [J]. Plant and Cell Physiology, 2005, 46(12): 1915-1923.
[24]Aebi H. Catalaseinvitro[J]. Methods in Enzymology, 1984, 105: 121-126.
[25]马晓渊. 农田杂草抗药性的发生为害, 原因与治理[J]. 杂草科学, 2002 (1): 5-9.
[26]Devine M D. Mechanisms of resistance to acetyl‐coenzyme A carboxylase inhibitors: a review [J]. Pesticide Science, 1997, 51(3): 259-264.
[27]Cummins I, Moss S, Cole D J, et al. Glutathione transferases in herbicide‐resistant and herbicide‐susceptible black‐grass (Alopecurusmyosuroides) [J]. Pesticide Science, 1997, 51(3): 244-250.
[28]Tal A, Romano M L, Stephenson G R, et al. Glutathione conjugation: a detoxification pathway for fenoxaprop-ethyl in barley, crabgrass, oat, and wheat [J]. Pesticide Biochemistry and Physiology, 1993, 46(3): 190-199.
[29]许长成, 樊继莲. 水稻对百草枯 (Paraquat)和一些环境胁迫交叉抗性[J]. 作物学报, 1996, 22(3): 358-361.
[30]吴进才, 刘井兰, 沈迎春, 等. 农药对不同水稻品种SOD活性的影响[J]. 中国农业科学, 2002, 35(4): 451-456.
[31]Choudhary M, Jetley U K, Abash K M, et al. Effect of heavy metal stress on proline, malondialdehyde, and superoxide dismutase activity in the cyanobacteriumSpirulinaplatensis-S5 [J]. Ecotoxicology and Environmental Safety, 2007, 66 (2): 204-209.
Resistance of Eleusine indica to quizalofop-P-ethyl in some cotton fields in Hunan Province
Zong Tao1, Li Jie1, Liu Xiangying1, Bai Lianyang2
(1. College of Plant Protection, Hunan Agricultural University, Changsha 410128, China;2.Hunan Academy of Agricultural Sciences, Changsha 410125, China)
In order to evaluate the resistance ofEleusineindicapopulations to quizalofop-P-ethyl, whole-plant bioassay was carried out and the differences in GSTs, SOD, POD and CAT of the resistant (R) and susceptible (S) populations to quizalofop-P-ethyl were preliminarily compared. The results of whole-plant bioassay showed that the populations ofE.indicahad different levels of resistance to quizalofop-P-ethyl, and the highest resistance index (RI) was 9.7, which was found in Xiangyin County. The activity of R population increased immediately, with a peak value of 2.6 times of the control, while the S population increased at first and then declined, with a peak value of 2.2 times. The SOD and POD activities of R population were significantly lower than that of S population in the medium term. The CAT activity of R population was significantly higher than that of S population after 9 days. The results indicated that the increased metabolic activity may be one of the resistance mechanisms to quizalofop-P-ethyl inE.indica. Oxidative stress may be related to the mode of action of quizalofop-P-ethyl.
Eleusineindica; quizalofop-P-ethyl; resistance
2014-01-06
2014-04-22
公益性行业(农业)科研专项(201303031);湖南省自然科学基金项目(14JJ3096)
S 481.4
A
10.3969/j.issn.0529-1542.2015.02.010
* 通信作者 E-mail:bailianyang2005@aliyun.com
猜你喜欢
今日农业(2019年14期)2019-09-18
世界农药(2019年3期)2019-09-10
世界农药(2019年3期)2019-09-10
农药科学与管理(2019年10期)2019-04-20
今日农业(2019年15期)2019-01-03
中学时代(2018年6期)2018-07-19
作文周刊·七年级读写版(2016年36期)2017-03-03
大作文(2016年5期)2016-05-14
杂草学报(2015年2期)2016-01-04
三月三(2015年11期)2015-11-16
