
杉木第一代种子园自由授粉子代表型性状的多样性
2015-12-21 03:02欧阳磊郑仁华苏顺德方禄明
中南林业科技大学学报 2015年3期
欧阳磊,郑仁华,肖 晖,苏顺德,方禄明
杉木第一代种子园自由授粉子代表型性状的多样性
欧阳磊1a,b,郑仁华1a,b,肖 晖1a,b,苏顺德1a,b,方禄明2
杉木是我国南方最重要的乡土针叶树种之一,广泛分布于我国亚热带的南方17个省区,经长期的自然选择和人工选择,不同产地的杉木个体在表型性状方面存在不同程度的变异。项目组对不同育种世代种子园自由授粉子代表型性状的多样性进行了研究。其中,对杉木第一代种子园自由授粉子代表型性状多样性分析表明,杉木第一代种子园自由授粉子代的生长、结实、形质等表型性状存在较为丰富的变异,表型性状在家系间的差异均达显著或极显著水平。通过主成分分析和聚类分析,当距离阀值为3.5时,63个家系明显地被划分为4大类,当距离阀值为2.5时,可以进一步细分为8个亚类。第1大类包括14个家系,以来自湖北的家系为主;第2大类包括17个家系,以来自湖南和贵州的家系为主;第3大类包括17个家系,以来自广东和福建的家系为主;第4大类包括15个家系,以来自福建的家系为主。家系的分类与家系的地理分布基本一致,说明根据表型性状对杉木第一代种子园自由授粉子代家系进行分类是可行的。家系的分类将为杉木优良材料的选择、育种群体的构建、人工控制授粉以及长期遗传改良策略的制定提供参考。根据分类结果,对家系的变异系数进行合并后求平均值,发现不同类和亚类的变异系数值相差较大,不同亚类变异系数的变化范围为16.2%~24.8%。来自湖北咸宁家系的变异系数较大,来自福建三明和建阳家系的变异系数较小。
杉木;子代;表型性状;变异;多样性
林木表型表达是林木基因型与其所处立地环境条件相互作用的结果,是林木基因型在特定环境条件下的表象。至今,林木表型性状多样性与DNA分子标记多样性的研究仍是林木遗传多样性研究的主要内容,而表型多样性研究则是遗传多样性研究的前提。随着林木遗传改良的进展,林木表型性状多样性研究不断得到提高,表型性状的表现更趋复杂,利用表型性状来研究林木群体遗传多样性的难度不断增大,但利用表型性状来研究林木的遗传变异依然是最有效的方法。杉木Cunninghamia lanceolata(Lamb.)Hook是我国南方最重要的针叶用材树种之一,广泛分布于我国亚热带的南方17个省区,经长期自然选择和人工选择,不同地域的杉木群体和个体的表型性状存在着比较丰富的变异。有关杉木遗传变异规律和良种选育的报道较多,但对杉木表型性状多样性研究的报道较少[1-7]。鉴于此,笔者根据杉木第一代种子园自由授粉子代的表型性状,进行分差分析、主成分分析和聚类分析,分析子代间的多样性和亲缘关系,以期为杉木种质资源的分类、杂交制种亲本的选配和核心育种群体的建立提供依据。
1 材料与方法
1.1 试验地及参试家系
试验地位于福建省邵武卫闽国有林场(为杉木中心产区,117°40′E,27°05′N),海拔 350m,年均温17.7℃,年降水量1 767.7mm,年均相对湿度83.0%,年有效积温5 619℃。试验地前茬为天然阔叶林,立地等级为Ⅰ级。参试的杉木种子园自由授粉子代家系于1977年秋采自福建省国有来舟林业试验场的杉木嫁接种子园中,每个家系的种子采自同一无性系的3~5株生长正常的嫁接分株的混合种子,共采集63个家系的种子,并以种子园的混合种作对照,共64个处理,采用BIB设计,72个区组,9个重复,9株小区。各参试家系基本情况见表1。

表1 杉木第一代种子园自由授粉子代参试家系Table 1 Tested pedigrees of open-pollinated progeny of first generation Chinese fir breeding garden
1.2 数据收集整理
试验林营建成功后每年加强抚育管理,年终开展树高、胸径、地径(离地表5cm处)、冠幅、结实量、分枝角等性状的调查。通直度分为4个等级:主干5m内通直为4,1个弯为3,2个弯为2,3个及3个弯以上为1;结实量分为4个等级:果多为4,果较多为3,中等为2,少为1;分枝角为树干上最下一轮活枝中最大枝与主干的夹角;圆满度为胸径与地径的比值;
材积计算公式为:

式中:V为材积;D为胸径;H为树高。
各年度、各性状调查观测的数据经反复核对无误后输入计算机进行统计分析。对结实、通直度等得分性状经过平方根转换后再进行分析。
1.3 分析方法
1.3.1 方差分析
以单株观测值进行单点方差分析时的线性模型为:

式中:Yijk为第i家系在第j区组中的第k个观察值;μ为群体平均值;Fi为家系的效应值;Bj为区组的效应值;(F×B)ij第i个区组第j个家系的效应值;εijk为机误。
1.3.2 主成分分析
采用主成分分析,将众多相关的表型性状指标,重新组合成一组新的相互无关的几个综合变量,同时根据实际需要从中可以取出几个较少的综合变量尽可能多地反映原来变量的信息。
1.3.3 聚类分析
利用国际通用SAS8.1软件中的聚类模块进行聚类分析,生成聚类树状图,进行分类分析。
2 结果与分析
2.1 家系表型性状变异
表2列出了杉木第一代子代家系表型性状的变异系数。由表2可知,生长性状的变异系数最大,平均值变异系数值为0.263 2,其中,材积变异系数>胸径变异系数>树高变异系数。其次是结实性状,变异系数值为0.152 5。形质性状(包括圆满度、通直度和分枝角)的变异系数最小,圆满度、通直度和分枝角的变异系数分别只有0.023 2、0.052 1和0.023 8。总体上,杉木第一代种子园自由授粉子代的生长、结实等性状存在较为丰富的变异,形质性状表现比较稳定,子代林整体表现比较圆满、通直。
2.2 家系表型性状方差分析
表3列出了杉木第一代种子园自由授粉子代家系表型性状方差分析结果,包括重复间、家系间及机误的均方以及重复间、家系间的F值。经检验,杉木第一代种子园自由授粉子代家系各表型性状中除圆满度在重复间没有达到差异显著水平外,其余性状在重复间的差异均达极显著水平,说明立地环境条件对杉木第一代种子园自由授粉子代家系的表型性状表现影响较大。在家系水平上,除2a地径、3a树高、3a地径、圆满度和通直度在家系间没有达到差异显著水平外,其余表型性状在家系间的差异均达显著或极显著水平,说明不同家系的表型性状表现存在显著差异,家系间的变异比较丰富。
2.3 家系表型性状多样性区划
2.3.1 主成分分析和聚类分析
从杉木第一代种子园自由授粉子代家系的29个表型性状主成分分析结果可知(见表4),第一个主成分的贡献率达94.11%,第二个主成分的贡献率为1.87%,第三个主成分的贡献率为1.26%,前3个主成分累计贡献率达97.24%。因此,可以得出保留3个主成分将能大部分反映原来29个表型性状变量的信息。
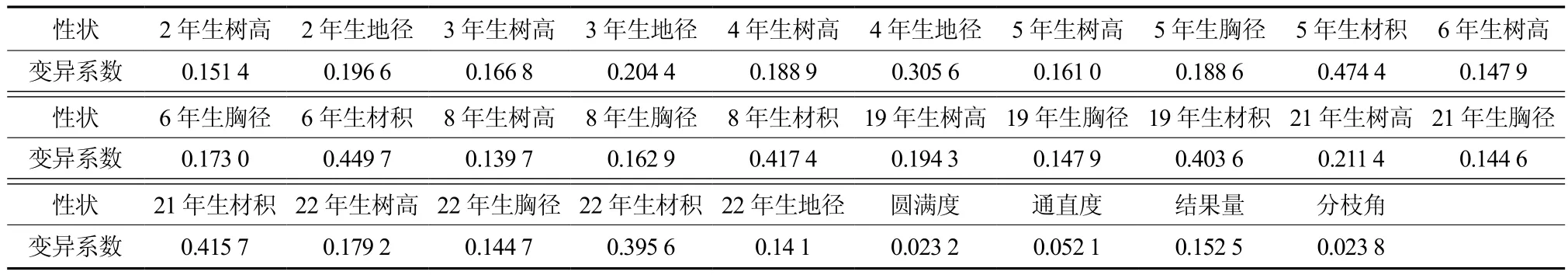
表2 杉木第一代种子园自由授粉子代家系表型性状变异系数†Table 2 Variation coefficients of phenotypic traits of first generation single parent offspring of Chinese fir

表3 杉木第一代种子园自由授粉子代家系表型性状方差分析结果Table 3 Variance analysis on phenotypic traits of first generation single parent offspring of Chinese fir

表4 杉木第一代种子园自由授粉子代表型性状主成分分析结果Tablt 4 Principal component analysis on phenotypic traits of Chinese fir first generation progeny
在主成分分析的基础上进行聚类分析。根据杉木第一代种子园自由授粉子代家系2年生树高等29个表型性状进行UPGMA聚类分析,当距离阀值为3.5时,可将参试的63个家系明显的划分为4大类(见图1)。从图1可知,湖北咸宁1、湖北咸宁2、湖北咸宁3、湖北咸宁4、湖北咸宁5、湖北咸宁7、湖北咸宁8、湖北咸宁9、湖北咸宁10、湖北咸宁14、湖北咸宁45、湖北咸宁46、湘校3和湘校4形成第一大类,第一大类主要为来自湖北的家系(占85.7%);湖北咸宁11、湘校5、湘广1、湘广7、湘广9、湘排329、湘排360、湘排403、贵州锦屏4、贵州锦屏8、贵州锦屏优9、贵州锦屏10、贵州锦屏13、贵州锦屏40、贵州锦屏45、贵州锦屏48和贵州锦屏54形成第二大类,第二大类主要为来自湖南和贵州的家系(占94.1%);贵州锦屏62、广东2、广东5、广东7、广东10、赣州10、建44、建优58、建64、建111、建19、顺53、浦城5、崇安15、三45、三优54和三优67形成第三大类,第三大类主要为来自广东和福建的家系(占88.2%);广东6、建105、建106、建优107、建1、建26、顺41、顺54、顺58、浦城19、三优7、三12、三17、三优21和三25形成第四大类,第四大类主要为来自福建的家系(占93.3%)。

图1 杉木第一代种子园自由授粉子代家系表型性状聚类Fig.1 Dendrogram of phenotypic traits of Chinese fi r fi rst generation progeny
2.3.2 表型多样性划分的特点
对杉木第一代种子园自由授粉子代家系间平均变异系数按4个大类和8个亚类(距离阀值为2.5)进行合并后求均值(见表5)。由表5可知,变异系数最大的是Ⅰ-1(24.8%)和Ⅰ-2(22.3%),其次是Ⅱ-2(21.1%),往下依次是Ⅱ-1(18.7%)>Ⅳ-1(17.7%)>Ⅲ-2(17.2%)>Ⅲ-1(16.9%)Ⅳ-2(16.2%)。归类后发现,杉木第一代种子园自由授粉子代家系间存在丰富的遗传变异,不同亚类群的遗传变异系数相差较大,变化范围16.2%~24.8%。来自湖北咸宁子代家系的变异系数较大,来自福建三明和建阳子代家系的变异系数较小。
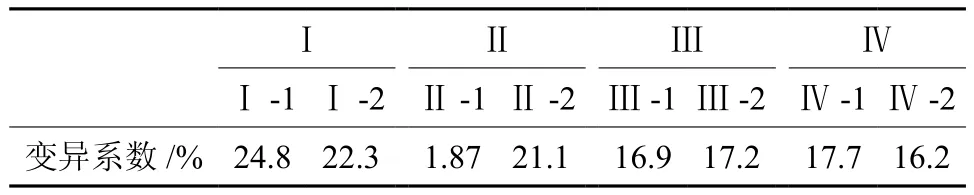
表5 杉木第一代种子园自由授粉子代家系变异系数归类分析Table 5 Classification analysis on variation coefficients of first generation single parent offspring of Chinese fir
3 结论与讨论
表型性状多样性是生物多样性的基础,掌握植物表型多样性及其变化规律,对于植物资源的保护及可持续利用具有十分重要的意义[8-10]。不同杉木第一代种子园自由授粉子代造林后,表型性状产生了丰富的变异。林木表型表达是林木基因型与其所处立地环境条件相互作用的结果,是林木基因型在特定环境条件下的表象。表型变异越大,可能存在的遗传变异越大。根据杉木第一代种子园自由授粉子代丰富的变异情况,可以推断其改良前景是广阔的。因此,在杉木的遗传改良工作中,杉木表型性状多样性研究和优良材料的选择应受到重视。
依据子代家系的表型性状特征,可将63个参试子代家系划为4个不同的大类,进一步可细分为8个不同的亚类。从分类结果来看,第1大类包括14个家系,以来自湖北的家系为主(12个),另2个家系来自湖南;第2大类包括17个家系,以来自湖南和贵州的家系为主(16个),另1个家系来自湖北;第3大类包括17个家系,以来自广东和福建的家系为主(15个),另2个家系分别来自贵州和江西;第4大类包括15个家系,以来自福建的家系为主(14个),另1个家系来自广东。家系的分类与家系的地理来源基本一致,说明根据家系的表型性状对杉木第一代种子园自由授粉子代家系进行分类是可行的。家系的分类将为杉木优良材料的选择、育种群体的构建、人工控制授粉等提供参考。
杉木作为我国南方最重要的针叶用材树种之一,广泛分布于我国亚热带的南方17个省区,不同产地的杉木个体在表型性状方面存在不同程度的变异,经长期自然选择和人工选择,在叶片、叶色、开花、结实、种子、球果、干型、材性、生长速度等各项指标上存在较大的变异,蕴含着丰富的群体/个体的多样性差异。自上世纪六十年代以来,我国开始杉木遗传改良研究,杉木遗传改良研究水平一直处于国内领先地位,以种子园、种质资源库库和子代林等多种方式保存了大量丰富的育种资源。如何评价和利用现存的杉木育种资源是我国现阶段杉木遗传改良的重要任务之一,具有承前启后的重要意义。本研究着重从表型性状方面分析杉木第一代种子园自由授粉子代表型性状的多样性,尚未涉及不同子代的其它性状包括同工酶、蛋白质和DNA分子标记等的多样性,虽相关研究已有少量报道[11-18],但仍有待不断深入和系统研究。
[1] 施季森,叶志宏,翁玉榛.杉木生长与材性联合遗传改良研究[J].南京林业大学学报,1993,17(1):1-8.
[2] 郑仁华.杉木种子园自由授粉子代遗传变异及优良遗传型选择[J].南京林业大学学报,2006,30(1):8-12.
[3] 韦如萍,胡德活,郑会全,等.杉木优树生长性状和材质性状的研究[J].中南林业科技大学学报,2013,33(2):28-33.
[4] 王润辉,胡德活,郑会全,等.杉木2.5代种子园开花物候遗传变异分析[J].西南林业大学学报,2013,33(4):25-29.
[5] 王润辉,胡德活,郑会全,等.杉木无性系生长和材性变异及多性状指数选择[J].林业科学,2012,48(3):45-50.
[6] 齐 明,何贵平,李恭学,等.杉木不同水平试验林的遗传参数估算和高世代育种的亲本评选[J].东北林业大学学报,2011,39(5):4-8.
[7] 陈孝丑.杉木耐瘠薄速生优树子代测定及早期选择[J].林业科技开发,2013,27(4):55-57.
[8] 姚淑均,张守攻,王军辉,等.滇楸花部性状的表型多样性研究[J].中南林业科技大学学报,2013,33(10):20-24.
[9] 张强英,张启翔,程堂仁.榆叶梅野生群体表型多样性的研究[J].中南林业科技大学学报,2012,32(5):156-160.
[10] 文亚峰,韩文军,吴 顺.植物遗传多样性及其影响因素[J].中南林业科技大学学报,2010,30(12):80-86.
[11] 黄敏仁,陈道明,施季森,等.杉木种源酯酶同工酶地理分布研究[J].南京林业大学学报,1986,3:31-35.
[12] 易能君,施季森,王明庥.杉木群体遗传多样性和多位点遗传结构[J].生物多样性,1996,4(3):153-159.
[14] 陈伯望,洪菊生,施行博.杉木和秃杉群体的叶绿体微卫星分析[J].林业科学,2000,36(3):46-51.
[15] 李 梅,施季森,甘四明,等.杉木杂交亲本分子遗传变异与子代生长相关的研究[J].林业科学研究,2001,14(1):35-40.
[16] 李 梅,施季森,李发根,等.杉木第2代育种群体优良基因型的RAPD遗传变异[J].林业科学,2007,43(12):50-55.
[17] 齐 明.杉木远交亲本群体遗传多样性研究[J].植物研究,2008,23(8):299-303.
[18] 杨玉玲,马祥庆,张木清.不同地理种源杉木的分子多态性分析[J].热带亚热带植物学报,2009,17(2):183-189.
Phenotypic genetic diversity of open-pollinated progenies of fi rst generation Chinese fi r seed orchard
OUYANG Lei1a,b,ZHENG Ren-hua1a,b,XIAO Hui1a,b,SU Shun-de1a,b,FANG Lv-ming2
(1a.Key Laboratory of Timber Forest Breeding and Cultivation for Mountainous Areas in Southern China;b.State Forestry Administration Engineering Research Center of Chinese Fir,Fujian Academy of Forestry,Fuzhou 350012,Fujian,China;2.Jiangle National Forest Farm of Fujian,Jiangle 353300,Fujian,China)
The phenotypic diversity of the fi rst generation of Chinese fi r seed orchard pollination progeny was investigated.The results were as follows.There were signi fi cant or very signi fi cant differences among progenies in growth traits,seed production and tree form traits,and the phenotypic variation was abundant.Through principal component analysis and cluster analysis,sixty three families were obviously divided into four main groups when the distance threshold was 3.5,and were subdivided into eight sub-groups when the distance threshold was 2.5.The fi rst main group included 14 families,which mainly came from Hubei province;the second main group included 17 families,which mainly came from Hunan and Guizhou provinces;the third main group included 17 families,which came from Guangdong and Fujian provinces;the fourth main group includes 15 families,which came from Fujian provinces.The classi fi cation of families was in some degree consistent with the geographical distribution of families,which indicated it is feasible for the classi fi cation of the fi rst generation Chinese fi r seed orchard pollinated progenies based on phenotypic traits.Pedigree classi fi cation would be useful for genetic material selection,breeding populations building,artificial hybrid seeding and long-term genetic improvement strategy.According to the classi fi cation results,there were lager differences among main groups and sub-groups in the averages of the variation coef fi cients,and the variation coef fi cients ranged from 16.2%~24.8%.The variation coef fi cient families from Xianning of Hubei were more larger than others,and Sanming and Jianyang of Fujian were relatively small.
Cunninghamia lanceolata;progeny;phenotypic traits;variation;diversity
(1.福建省林业科学研究院,a.国家林业局南方山地用材林培育重点实验室;b.国家林业局杉木工程技术研究中心,福建 福州 350012;2.福建省将乐国有林场,福建 将乐 353300)
S791.27
A
1673-923X(2015)03-0022-05
10.14067/j.cnki.1673-923x.2015.03.005
2014-01-10
福建省林木种苗科技攻关三期、四期项目“杉木速生优质高产新品种定向选育研究与应用”(闽林科[2009]4号、[2013]1号);福建省自然科学基金项目“杉木第三代种质资源遗传多样性分析及核心种质构建研究”(2012J01097);福建省科技重大专项专题“杉木优质种苗繁育技术及人工林高效培育技术”(2012NZ0001-1);“十二五”农村领域国家科技计划专题(2012BAD01B0201);福建省森林培育与林产品加工利用重点实验室项目
欧阳磊,高级工程师,博士
郑仁华,教授级高工,博士,E-mail:zrh08@126.com
欧阳磊,郑仁华,肖 晖,等.杉木第一代种子园自由授粉子代表型性状的多样性[J].中南林业科技大学学报,2015,35(3):22-26.
[本文编校:吴 毅]
猜你喜欢
浙江林业科技(2022年4期)2022-07-16
当代水产(2022年1期)2022-04-26
文萃报·周五版(2021年4期)2021-09-13
辽宁林业科技(2021年3期)2021-07-08
山西水土保持科技(2020年2期)2020-12-22
东方少年·布老虎画刊(2020年3期)2020-06-09
家教世界·创新阅读(2018年9期)2018-12-17
大江南北(2016年8期)2016-02-27
浙江林业(2015年4期)2015-12-21
中国火炬(2012年9期)2012-07-25
