
人工养殖下施氏鲟Acipenser schrenckii、达氏鳇Huso dauricus及其杂交后代的繁殖特性
2016-02-07 09:43张颖曲秋芝王斌夏永涛许式见孙大江
水产学杂志 2016年3期
张颖,曲秋芝,王斌,夏永涛,许式见,孙大江
(1.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070;2.杭州千岛湖鲟龙科技股份有限公司,浙江 杭州 311700)
人工养殖下施氏鲟Acipenser schrenckii、达氏鳇Huso dauricus及其杂交后代的繁殖特性
张颖1,曲秋芝1,王斌2,夏永涛2,许式见2,孙大江1
(1.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070;2.杭州千岛湖鲟龙科技股份有限公司,浙江 杭州 311700)
2013年11月~2014年11月,测量5批次施氏鲟Acipenser schrenckii、黑龙江杂交鲟(达氏鳇Huso dauricus♀×施氏鲟♂)和达氏鳇Huso dauricus(共644尾)的体长、体质量,观察其生长、个体繁殖力、卵巢发育、卵径、卵质量和卵色等。结果表明:3种鲟雌亲鱼的体长与体质量差异显著,施氏鲟的体长和体质量显著小于杂交鲟与达氏鳇(P<0.05),而杂交鲟的体长和体质量则显著小于达氏鳇(P<0.05)。施氏鲟、杂交鲟和达氏鳇雌亲鱼的成熟系数(GSI)和繁殖力差异显著,杂交鲟的GSI和平均绝对繁殖力显著高于施氏鲟和达氏鳇(P<0.05),而施氏鲟与达氏鳇的GSI无显著性差异(P>0.05)。杂交鲟的相对繁殖力(Fw)显著高于施氏鲟与达氏鳇。3种鲟雌亲鱼产卵质量与体质量比值以10%~15%为主,该比值范围内雌性施氏鲟个体数约占同龄雌性个体的42.66%,杂交鲟为42.91%,达氏鳇为41.93%。施氏鲟和杂交鲟的卵多为棕色,而达氏鳇的卵为棕灰色。达氏鳇的卵径显著大于施氏鲟与杂交鲟。杂交鲟的绝对繁殖力、相对繁殖力及成熟系数均高于施氏鲟与达氏鳇,具有明显的杂交优势。
施氏鲟;达氏鳇;杂交种;繁殖力
鲟是我国重要的淡水经济鱼类,鲟鱼子酱价格昂贵,被比作“黑色黄金”,是国际市场的高档营养食品[1,2]。我国是世界最大的鲟养殖国,2011年我国鲟养殖产量达44 211t,占世界鲟养殖总产量的84.97%[2,3]。随着鲟养殖业的快速发展出现诸多问题,如缺乏优良选育品种、杂交品种混乱、许多养殖品种生长及抗逆性降低等,这严重制约了我国鲟产业的健康发展。苏联、德国、美国和伊朗等均培育出适合本土养殖的鲟品种,如苏联的小鳇鲟(欧鳇Huso huso♀×小体鲟Acipenser ruthenus♂)、德国的西伯利亚鲟(A.baeri)、美国的高首鲟(A.transmontanus)和匙吻鲟(Polyodon spathula)等具有生长快、性成熟早等优点[4-7]。目前急需开发一些适合我国本土养殖的鲟新品种,优化和改善我国鲟养殖品种结构。
黑龙江杂交鲟为达氏鳇(H.dauricus♀)和施氏鲟(A.schrenckii♂)的杂交后代,在黑龙江的自然水域中有一定数量分布[8]。自2007年人工养殖的黑龙江杂交鲟性成熟后,繁殖性能明显高于其他人工养殖的鲟品种。许多养殖实践也证明黑龙江杂交鲟的生长性能较好。但有关黑龙江杂交鲟的研究报道较少,主要集中在养殖和遗传生物学等方面[9,10]。而有关达氏鳇、施氏鲟及其杂交种的繁殖性能及鱼子酱生产性能等研究还未见报道。提高鲟繁殖性能是提高鱼子酱产量和质量的重要条件。通过研究提高鲟杂交后代的繁殖力,掌握不同鲟亲鱼群体的繁殖规律,可更好地利用杂交后代的丰产性状。本文采用形态学和统计学的方法,比较分析达氏鳇、施氏鲟及其杂交种的生长、繁殖及鱼子酱生产性能,为鲟新品种选育提供参考。
1 材料与方法
1.1 材料
实验鱼随机取自杭州千岛湖鲟龙开发有限公司流水养殖网箱(6m×6m×6m),其中性成熟雌达氏鳇36尾(15+龄)、施氏鲟305尾(9+龄)、黑龙江杂交鲟(10+龄)300尾。每天测量水温2次,年养殖水温变化范围为11~32℃。每天投喂2次鲟商品鱼饲料(购于中山统一企业有限公司),投饲量为体质量的1.5%~3%。
1.2 方法
实验鱼暂养在水族箱中,先测量鲟体质量与体长,然后解剖取出性腺,测量长度和质量后,放入5%的甲醛溶液中保存。取整份卵巢计算繁殖力,完全充满卵黄、即将产出的卵母细胞用肉眼直接观察计数。
繁殖力计算公式:
成熟系数(GSI)=[卵巢质量/体质量(毛质量)] ×100%;
绝对繁殖力F=个体怀卵量;
相对繁殖力FL=绝对繁殖力/体长;
相对繁殖力Fw=绝对繁殖力/体质量(毛质量)。
生长的测定:测量不同年龄达氏鳇的全长及体质量,采用Excel 2003和SPSS 13.0软件处理统计数据。肥满度(K)=(W/L3)×100,式中W为体质量(g),L为体长(cm)。绝对体长增长量△L=L2-L1,绝对体质量增长量△W=W2-W1,式中L2和L1为初始体长和末体长,W2和W1为初始体质量和末体质量。
1.3 数据分析
所得数据采用SPSS11.5软件进行方差分析。
2 结果与分析
2.1 3种鲟雌亲鱼的生长比较
3种鲟雌亲鱼的平均体长和体质量差异显著(图1、表1),其中施氏鲟雌亲鱼的平均体长和体质量显著小于雌杂交鲟(P<0.05),而杂交鲟雌亲鱼显著小于达氏鳇雌亲鱼(P<0.05);而杂交鲟的体长和体质量则显著小于达氏鳇(P<0.05)。3种鲟雌亲鱼中,施氏鲟雌亲鱼的体质量范围以15~20kg为主,杂交鲟雌亲鱼的体质量以30~40kg为主,少部分杂交鲟的体质量超过60kg(图2)。但是,达氏鳇亲鱼的体质量均超过70kg,以90~100kg范围为主。

图1 3种鲟亲鱼的体长频率分布Fig.1 The frequency of body length in broodstock of three sturgeons

图2 3种鲟亲鱼的体质量频率分布Fig.2 The frequency of body weight in broodstock of three sturgeons

表1 3种鲟亲鱼的生长指标Tab.1 The growth indices of Amur sturgeon(Acipenser schrenckii),kaluga sturgeon(Huso dauricus)and their hybrid(kaluga sturgeon♀×Amur sturgeon♂)
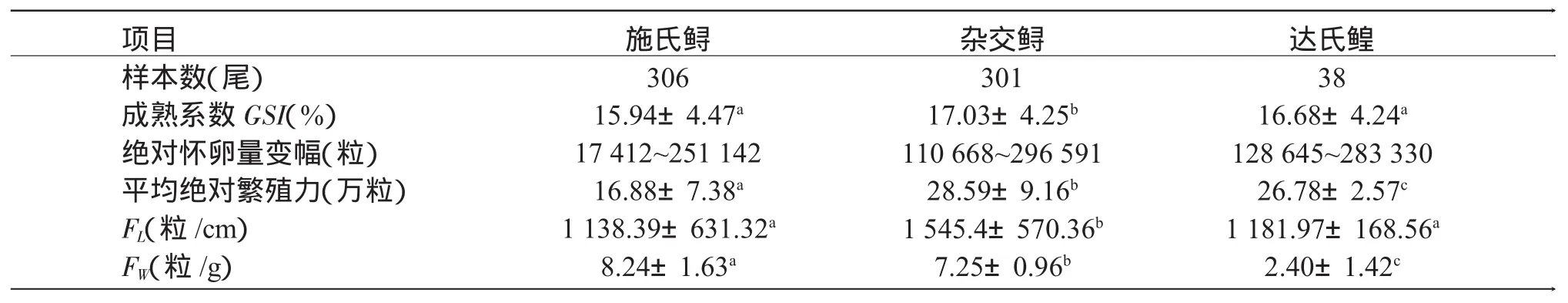
表2 3种鲟亲鱼的生物学指标和个体繁殖力Tab.2 The biological indices and individual fecundity of Amur sturgeon(Acipenser schrenckii),kaluga sturgeon(Huso dauricus)and their hybrid(kaluga sturgeon♀×Amur sturgeon♂)

表3 3种鲟亲鱼的卵径及卵质量Tab.3 The egg diameter and egg weight of Amur sturgeon(Acipenser schrenckii),kaluga sturgeon(Huso dauricus)and their hybrid(kaluga sturgeon♀×Amur sturgeon♂)
2.2 3种鲟雌亲鱼的个体繁殖力
3种鲟雌亲鱼的成熟系数和繁殖力差异显著(表2),其中杂交鲟的GSI和平均绝对繁殖力显著高于施氏鲟和达氏鳇(P<0.05),而施氏鲟与达氏鳇的GSI无显著性差异(P>0.05)。3种鲟亲鱼中,施氏鲟的平均绝对繁殖力最低,显著低于杂交鲟和达氏鳇(P<0.05)。而杂交鲟的绝对繁殖力最高,为28.59万粒,显著高于施氏鲟的16.88万粒和达氏鳇的26.78万粒(P<0.05)。施氏鲟的平均相对繁殖力FL为1 138.39粒/cm,低于杂交鲟的1 545.4粒/cm和达氏鳇的1 181.97粒/cm。杂交鲟的相对繁殖力FW为7.25粒/g,显著低于施氏鲟的8.24粒/g,高于达氏鳇的2.40粒/g(P<0.05)。相对繁殖力FL以杂交鲟为最高,相对繁殖力FW以施氏鲟为最高。
2.3 3种鲟亲鱼个体繁殖力与体长、体质量的关系
回归分析表明3种鲟雌亲鱼的个体繁殖力与体长和体质量呈二次函数关系,回归方程式分别为:
F施氏鲟=0.021W2-0.61W+6.86(R2=0.46),
F施氏鲟=0.0049 L2-1.38L+99.33(R2=0.37);
F杂交鲟=292.96W2-19819W+586935(R2=0.57),
F杂交鲟=0.0082L2-2.84L+252.51(R2=0.52);
F达氏鳇=-5114.3W2+105216W-456332(R2=0.65),
F达氏鳇=30.52L2-6 057.5L+323 741(R2=0.58)。
2.4 3种鲟亲鱼的卵径、卵质量、卵色及产卵质量
3种鲟亲鱼的卵色分布频率、卵径及卵质量见图3和表3。施氏鲟和杂交鲟的卵主要以棕色为主,而达氏鳇的卵则以棕灰色为主。施氏鲟卵呈棕色、棕灰色、灰黄色、黑棕色和棕黄色5种颜色,杂交鲟的卵有棕色、棕灰色、灰黄色、黑棕色、黑灰色、棕黄色和棕黑色7种颜色,而达氏鳇的卵则呈棕色、棕灰色、灰黄色和棕黄色4种颜色。达氏鳇的平均卵径显著大于施氏鲟与杂交鲟,分别为3.32mm、3.09mm和2.92mm。达氏鳇的平均单粒卵质量大于施氏鲟和杂交鲟,但无显著性差异。
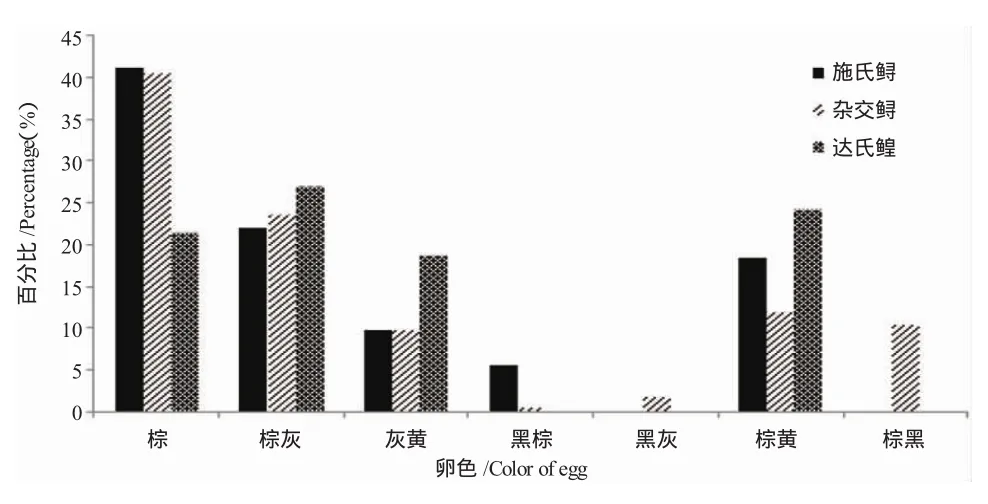
图3 3种鲟亲鱼卵子的颜色分布频率Fig.3 The frequency of egg color weight in broodstock of three sturgeons
3种鲟雌亲鱼中产卵质量与体质量比的个体比例分布规律各不相同(图4)。3种鲟雌亲鱼的产卵质量与体质量比值以10%~15%为主,其中雌施氏鲟的个体数占同龄雌性个体数的42.66%,而杂交鲟和达氏鳇分别占42.91%和41.93%。施氏鲟雌亲鱼中,产卵质量与体质量比值为5%~10%的雌性个体约占同龄雌性个体的35.84%,而杂交鲟占23.18%,达氏鳇不足17%。雌达氏鳇群体中,以产卵质量与体质量比值为15%~20%的雌性个体最多,约占同龄雌性达氏鳇的38.71%,杂交鲟次之为22.14%,施氏鲟最小为9.89%。

图4 3种鲟亲鱼产卵质量与体质量比的分布频率Fig.4 Thefrequency ofratio ofegg weightto in broodstock of three sturgeons body
3 讨论
3.1 雌亲鱼繁殖力的比较
鱼类繁殖力为物种或种群对环境的适应特征,是遗传和环境共同作用的结果,具有种属的特性[11-13]。鲟为一次产卵类型鱼类,分为春、秋季产卵两种类型,一生多次产卵,个体繁殖力巨大[14-16]。国内外有关鲟个体繁殖力的研究多数集中在野生鲟,而养殖鲟的报道较少,仅王斌等[17]报道了5种养殖鲟怀卵质量的差异。研究表明,野生中华鲟A. sinensis的怀卵量变动范围为20~130万粒[15],野生铲鲟Scaphirhynchus platorynchus为5~17万粒[18]。体长为105~241cm的野生施氏鲟个体的相对繁殖力为11.28粒/g,而体长为82.7~88.6cm的野生铲鲟的个体繁殖力变动范围为2.7~20.3粒/g,而野生中华鲟的相对怀卵量为2.99粒/g[14-19]。不同种鲟的怀卵量差异具有种属特异性[17]。本研究中,养殖杂交鲟的怀卵量变幅为11 068~29 591粒,高于施氏鲟和达氏鳇,这可能与种属特性有关,表明杂交鲟的繁殖力强[12,17]。
鱼类的繁殖力与其年龄、营养条件及栖息水域等因素有关[13]。刘鉴毅等[14]研究表明,随着栖息环境的变化葛洲坝下野生中华鲟的繁殖力下降。与野生鱼相比,养殖鲟的饲养条件较好,在人工繁殖过程中发现,多数养殖鲟的性腺中含有大量的脂肪,而野生鲟的卵巢中脂肪含量很低,因此养殖鲟的繁殖力低于同种同规格的野生鲟的繁殖力。本研究中,3种养殖鲟的怀卵量均低于野生中华鲟,但高于野生铲鲟[15,19]。对圆筒体型黄鳝Monopterus albus的研究表明,黄鳝的个体绝对繁殖力与体长相关,这与其他一些鱼类是相似的;而与体质量不相关,与一般鱼类不同[12]。本研究中,除达氏鳇的个体繁殖力与体质量的相关系数高于体长外,杂交鲟和施氏鲟的个体繁殖力与体长和体质量的关系基本相同。王斌等[17]研究表明,鲟科鱼类的产卵质量随体质量的增加而增加。
鲟个体大、性成熟晚、繁殖力巨大,种群的增殖潜力小,为典型的k型选择鱼类,自然水体的种群遭遇破坏后,一般不易恢复[20]。近些年,野生鲟资源急剧衰退,鲟资源的保护应该依据其生殖特点进行人工增殖。本研究中的施氏鲟和杂交鲟均为黑龙江中的土著鲟种类,应通过保护其产卵场、划定繁殖保护期等手段和措施进行保种[20,21]。
3.2 鲟的种间杂交
杂交是一种有效的品种改良方法,在动物的新种制备和遗传改良中有重要作用[22,23]。杂交能够有效地将双亲的遗传物质组合,改变杂交后代的遗传和表型[24],改良杂交后代的养殖性状、产量和抗逆性等[22-25]。与大多数鱼类不同,鲟形目鱼类种间甚至属间的杂交现象极为普遍,且大部分杂交种可育,产生三杂交后代[26]。迄今,国外报道的鲟杂交种有:小鲟鳇(欧鳇H.huso♀×小体鲟A.ruthenus♂)、西伯利亚鲟 A.baeri×小体鲟、俄罗斯鲟 A. gueldenstaedtii×小体鲟,和俄罗斯鲟×欧洲鳇等。国内养殖的鲟杂交种以施氏鲟×西伯利亚鲟和施氏鲟×达氏鳇为主,此外还包括少量的施氏鲟与其他鲟的杂交种,如俄罗斯鲟、小体鲟等[4,27-30]。鲟的种间杂交可改善其生长、外部形态及繁殖性能等[2,4,28-30]。本研究中,雌性杂交鲟的绝对繁殖力、成熟系数和产卵质量与体质量的比值(10%~15%),均高于施氏鲟和达氏鳇,具有明显的繁殖杂交优势。但雌性杂交鲟(欧鳇♀×小体鲟♂)的繁殖性能如怀卵量、成熟系数等要好于亲本[4]。
[1]孙大江,曲秋芝,张颖,等.中国的鲟鱼养殖[J].水产学杂志,2011,24(4):67-70.
[2]张颖,刘晓勇,曲秋芝,等.达氏鳇、施氏鲟及其杂交种(施氏鲟♂×达氏鳇♀)形态差异与判别分析[J].淡水渔业,2012,42(6):27-32.
[3]孙大江,张颖,马国军.鲟鱼子酱的生产与国际贸易概况[J].水产学杂志,2014,27(1):1-7.
[4]Ishihara M,Tokui B,Abe T,et al.Seasonal changes in oocyte maturational competence,ovulatory competence and gonadotropin sensitivity of ovarian follicles in hybrid sturgeon,bester(Huso huso×Acipenser ruthenus)[J].Journal ofApplied Ichthyology,2014,30(6):1141-1148.
[5]AdrioF,Rodriguez-Moldes I and Anadon R.Distribution of glycine immunoreactivity in the brain of the Siberian sturgeon(Acipenser baeri):comparison with gamma-aminobutyric acid[J].Journal ofComparative Neurology,2011,519(6):1115-1142.
[6]SeunghyungL,Yichuan W,Silas S O H,et al.Development ofoptimumfeedingrate model for white sturgeon(Acipenser transmontanus)[J].Aquaculture,2014,433:411-420.
[7]Patterson J T,Mims S D and Wright R A.Effects of body mass and water temperature on routine metabolism of American paddlefish Polyodon spathula[J].Journal ofFish Biology,2013,82(4):1269-1280.
[8]宋聃,张颖,麦丽坎,等.饥饿和不同饵料对杂交鲟(达氏鳇(♀)×施氏鲟(♂))仔稚鱼生长和消化酶活性的影响[J].淡水渔业,2015(4):76-81.
[9]李斌,杨军,黄建华,等.三种微生态制剂对杂交鲟(达氏鳇♀×史氏鲟♂)生长性能及免疫机能的影响[J].淡水渔业,2014(1):78-83.
[10]高宇,袁改玲,李大鹏,等.杂交鲟和匙吻鲟HSP70 cDNA克隆与序列分析[J].华中农业大学学报,2010,29(1): 85-89.
[11]庹云.袁河花鱼骨个体繁殖力研究[J].四川动物,2013,32(3):375-379.
[12]杨代勤,陈芳,刘百韬,等.黄鳝产卵类型及繁殖力的研究[J].湖北农学院学报,1994,14(3):40-44.
[13]张觉民.黑龙江鲟鱼的体长与体重繁殖力关系[J].自然资源研究,1983(2):53-55.
[14]刘鉴毅,危起伟,陈细华,等.葛洲坝下中华鲟繁殖生物学特性及其人工繁殖效果[J].应用生态学报,2007,8(6):1397-1402.
[15]George S G,Slack W T and Hoover J J.A note on the fecundity of pallid sturgeon[J].J Appl Ichthyol,2012,28: 512-515.
[16]四川省长江水产资源调查组.长江鲟鱼类生物学和人工繁殖研究[M].成都:四川科学技术出版社,1988.
[17]王斌,彭涛,夏永涛,等.5种养殖鲟鱼怀卵差异及周年繁殖[J].动物学杂志,2011,46(3):109-116.
[18]Henry C J and Ruelle R.A study of pallid sturgeon and shovelnose sturgeon reproduction[M].U S Fish and Wildlife Service,1992:19.
[19]Keenlyne KD,Grossman E Mand Jenkins LG.Fecundity of the pallid sturgeon[J].Trans Am Fish Soc,1992,121: 139-140.
[20]叶富良,陈刚.3种鲟科鱼类生活史类型的研究[J].湛江水产学院学报,1996,16(1):1-4.
[21]Bryan J L,Wildhaber ML,Papoulias D M,et al.Estimation of gonad volume,fecundity,and reproductive stage of shovelnose sturgeon using sonography and endoscopy with application to the endangered pallid sturgeon[J].J Appl Ichthyol,2007,23:411-419.
[22]Barton N H.The role of hybridization in evolution[J]. Mol Ecol,2001,10:551-568.
[23]MalletJ.Hybridspeciation[J].Nature,2007,446:279-283.
[24]王静,肖军,曾鸣,等.白鲫×红鲫杂交后代的遗传变异[J].中国科学:生命科学,2015,45(4):371-380.
[25]Seehausen O.Hybridization and adaptive radiation[J]. Trends Ecol Evol,2004,19:198-207.
[26]郭向贺,董颖,胡红霞,等.鲟鱼种质鉴定方法研究进展[J].生物技术通报,2014(2):56-63.
[27]Jähnichen H,Kohlmann Kand Rennert B.Juvenile growth of Acipenser ruthenus and 4 different sturgeon hybrids[J].Journal ofApplied Ichthyology,2007,15(4/5):248-249.
[28]曲秋芝,孙大江,宋苏祥,等.人工养殖史氏鲟仔鱼和幼鱼的试验[J].水产学杂志,1997,10(2):72-75.
[29]Lin Luo,Tieliang Li,Wei Xing,et al.Effects of feeding rates and feedingfrequencyon the growth performances of juvenile hybrid sturgeon,Acipenser schrenckii Brandt♀×A.baeriBrandt♂[J].Aquaculture,2015,448:229-233.
[30]Amiri B M,Maebayashi M,Adachi S,et al.Testicular development and serumsexsteroid profiles duringthe annual sexual cycle of the male sturgeon hybrid,the bester[J].J Fish Biol,1996,48:1039-1050.
ComparisonofReproductiveCharacteristicsAmongAmurSturgeon(Acipenserschrenckii), KalugaSturgeon(Husodauricus)andTheirHybrid(KalugaSturgeon♀×AmurSturgeon♂)
ZHANG Ying1,QU Qiu-zhi1,WANG Bin2,XIA Yong-tao2,XU Shi-jian2,SUN Da-jiang1
(1.Heilongjiang River Fisheries Research Institute,Chinese Academy of Fishery Sciences,Harbin 150070,China; 2.Hangzhou Qiandaohu Xunlong Sci-tech Co.Ltd.,Hangzhou 311700,China)
The growth,fecundity,egg color and egg size were studied in broodstock of Amur sturgeon(Acipenser schrenckii),kaluga sturgeon(Huso dauricus)and their hybrid(kaluga sturgeon♀×Amur sturgeon♂)from November 2013 to November 2014.The results showed that there were significant differences in growth in body length and body weight of three sturgeon broodstocks(P<0.05).The Amur sturgeon broodstock had significantly less body length and body weight than kaluga sturgeon and hybrid sturgeon did(P<0.05).There were significantly higher gonadosomatic index(GSI)and absolute fecundity in the hybrid sturgeon than those in kaluga sturgeon and Amur sturgeon(P<0.05),without significant difference between kaluga sturgeon and Amur sturgeon(P>0.05). The hybrid sturgeon had higher relative fecundity per gram than Amur sturgeon and kaluga sturgeon did.Among the broodstocks of three sturgeons,the ratio of egg weight to body weight was primarily within about 10%~15%,accounting for about 42.66%in Amur sturgeon,about 42.91%in hybrid sturgeon,and about 41.93%in kaluga sturgeon.In addition,Amur sturgeon and hybrid sturgeon had brown eggs in color,and the eggs of kaluga sturgeon showed brown gray in color.The kaluga sturgeon had larger eggs in diameter than Amur sturgeon and the hybrid did.The findings indicated that the hybrid sturgeon had higher fecundity than Amur sturgeon and kaluga sturgeon did,showing significant heterosis in fecundity.
Amur sturgeon;kaluga sturgeon;hybrid;fecundity
S965.215
A
1005-3832(2016)03-0025-05
2016-01-22
农业部公益性行业科研专项(201203055);国家自然基金青年基金(31201971).
张颖(1977-),女,博士,副研究员,从事鱼类繁殖生理研究.E-mail:juletzhang@hotmail.com
孙大江,研究员.E-mail:13845059350@163.com
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
海洋渔业(2021年1期)2021-03-18
水生生物学报(2021年1期)2021-02-04
科学养鱼(2020年7期)2020-12-20
渔业致富指南(2020年6期)2020-12-19
当代水产(2018年8期)2018-11-02
猪业科学(2018年4期)2018-05-19
渔业致富指南(2017年8期)2017-02-02
猪业科学(2016年1期)2016-03-30
