
黄海中部近岸产卵场日本鳀卵子大小的时空变化❋
2016-03-24 09:18李秀梅叶振江李增光胡海生于洪亮
中国海洋大学学报(自然科学版) 2016年2期
关键词:时空变化
李秀梅, 叶振江❋❋, 李增光, 胡海生, 邹 全, 于洪亮
(1.中国海洋大学水产学院,山东 青岛 266003; 2.威海市文登区海洋与渔业局,山东 威海 264400)
技术报告
黄海中部近岸产卵场日本鳀卵子大小的时空变化❋
李秀梅1, 叶振江1❋❋, 李增光1, 胡海生1, 邹全1, 于洪亮2
(1.中国海洋大学水产学院,山东 青岛 266003; 2.威海市文登区海洋与渔业局,山东 威海 264400)
摘要:2013年4月下旬至7月上旬,在黄海中部近岸产卵场20 m以浅水域进行6个航次的鱼类浮游生物调查,对日本鳀(Engraulis japonicus)卵子形态及大小的时空变化进行研究。结果表明:调查海区日本鳀卵子形态呈椭球形,卵子大小在不同发育阶段无显著性差异,但其时空变化差异显著。从5月上旬至7月上旬,卵子大小整体上呈逐渐减小的趋势,且与水温呈显著负相关。日本鳀卵子在调查海区最早出现于5月上旬,卵子最大,长径(1.57±0.09)mm,短径(0.77±0.04)mm,纵切面积(0.95±0.07)mm2;6月下旬卵子最小,长径(1.20±0.07)mm,短径(0.64±0.03)mm,纵切面积(0.61±0.43)mm2。同历史数据相比,调查海区的日本鳀卵子5月上旬显著大于东海海域,6月下旬显著小于山东半岛南部。
关键词:黄海中部;海州湾;日本鳀;卵子大小;时空变化
LI Xiu-Mei, YE Zhen-Jiang, LI Zeng-Guang, et al. Spatial and temporal changes of the egg size of Japanese anchovy (Engraulis japonicus) inhabiting the near shore spawning ground of central Yellow Sea [J]. Periodical of Ocean University of China, 2016, 46(2): 54-60.
卵子大小与繁殖力是鱼类生活史上的2个重要生殖习性特征,其研究对解析渔业资源数量变动及对环境变动的应答机制具有重要意义[1]。生物广布种的形态、生理、行为和生态特征往往在不同地区存在显著的差异[2],鱼类繁殖力、产卵期等生殖习性特征可因海域与栖息环境而异[3-4]。鱼类卵子体积是体现鱼类遗传属性及环境适应性特征的重要生物学参数,对后代存活率、产卵场环境状况、鱼类群体划分等具有重要指示意义,相关研究有较多报道[5-6]。
日本鳀(Engraulisjaponicus)广泛分布于中国东海的西北部、黄海、日本海、日本九州岛西部近海、濑户内海和日本群岛的太平洋近海[7],作为主要饵料鱼种在海洋生态系统中起重要作用[8],为高资源量的小型中上层鱼类[9]。黄海日本鳀产卵期为5—10月,产卵期长[10]。有关中国近海日本鳀的研究,主要集中在产卵亲鱼的生殖生态学特征、群体结构以及卵子分布与环境的关系等,而关于其卵子大小仅零星报道中有所提及[11],缺乏深入系统的研究。黄海中部近岸海域是黄海主要产卵场之一,也是黄海日本鳀产卵场的重要组成部分,本文以黄海中部近岸水域为研究海域,对春夏季日本鳀卵子大小及时空分布做了初步研究,为该鱼种生殖习性现状及资源变动机制的研究提供依据。
1材料和方法
1.1 调查海域、站位与航次
以黄海中部海州湾近岸20m以浅水域(34.5°N~35.5°N,119.0°E~120.5°E)为调查海域,分别于2013年4月23—24日、5月10—12日、5月23—25日、6月6—8日、6月21—23日和7月6—8日(下简称4月下旬、5月上旬、5月下旬、6月上旬、6月下旬和7月上旬)开展了6个航次的调查,每半月大潮调查1次,共设置了75个调查站位(见图1),租用50~180马力调查渔船,采用浅水Ⅰ型网进行鱼类浮游生物斜拖采样(网长145cm,网口直径50cm,网口面积0.2m2),网口设置流量计以校正每网滤水量,每站位拖网时间为10min,拖速2kn左右。各站位以XR-420型CTD进行温度、盐度、水深等环境要素的测定。

图1 调查海域及站位分布
1.2 样品处理与图像采集
样品以5%海水甲醛溶液固定保存,样品的室内分析处理均按照《海洋调查规范》[12]中规定方法进行。实验室内以形态学方法,参考《中国近海鱼卵与仔鱼》[13]和《台湾海域鱼卵图鉴》[14],利用JSZ5型体视显微镜(20X)对鱼类浮游生物样品进行分类鉴定,依据Thompson &Riley鱼卵ⅴ期发育标准[15]对样品进行发育分期鉴定。利用浮游生物计数框和医用计数器,对各站位样品进行取样计数。已有研究表明日本鳀鱼卵和秘鲁鳀鱼卵在甲醛溶液中不会收缩,形状不会改变[16-17],因此,本文卵子大小量度指标的测量系直接对甲醛固定样品进行。
6个航次调查共采集到日本鳀的鱼卵747411粒。根据鱼卵密度分布特征,各航次分别在调查区域内选取主要鱼卵密集区,并兼顾其空间分布,确认4~6个图像采集站位(见图2),在各站位随机选取50粒卵子,在10×放大倍数下,使用SMZ800体视显微镜及图像采集系统进行卵子图像的采集,个别站位不足50粒卵的全取。6个航次中除4月下旬外,其余5个航次皆有鳀鱼鱼卵出现。实验共采集24个站位、1123粒的日本鳀卵子图像(见表1,图2)。
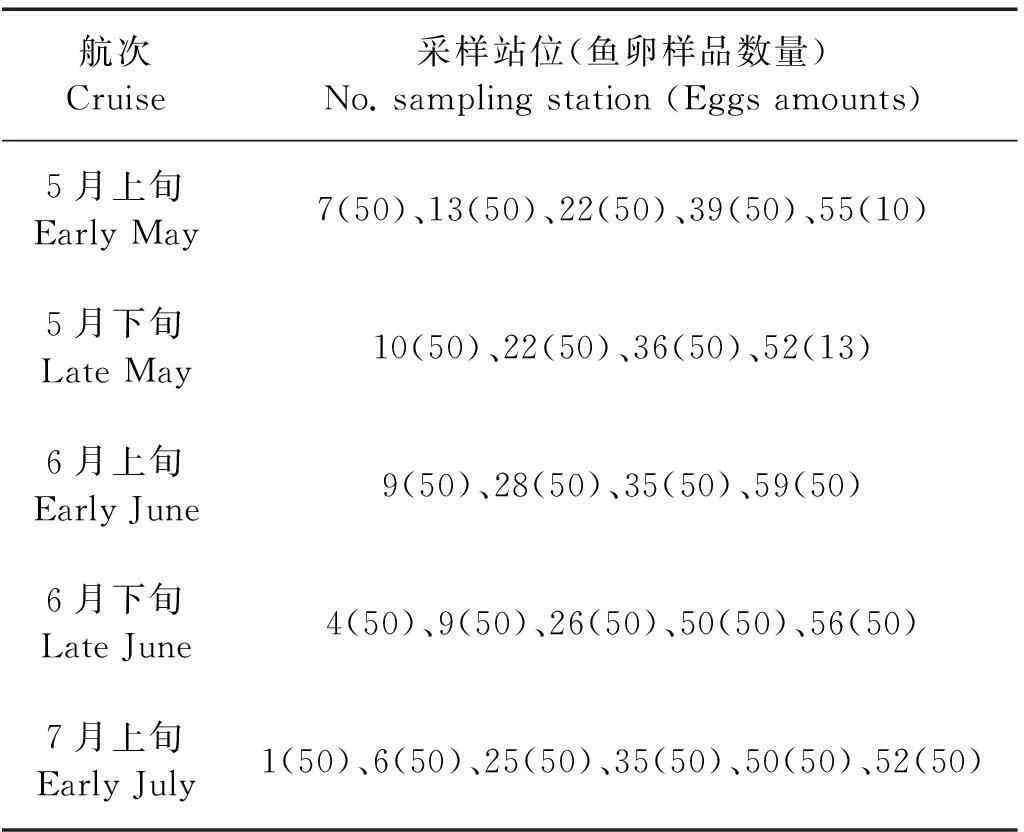
表1 各航次测量站位及鱼卵样品数量

图2 各航次采样站位位置及卵子总数
1.3 形态测量指标筛选及数据分析
日本鳀卵子呈椭球形。选取卵子长径、短径、长径/短径比及最大纵切面积(下简称面积)做为卵子大小量度指标,以Image-Pro Plus软件对卵子图像进行量度指标的测量。
通过单因素方差分析one-way ANOVA(满足方差齐性)或非参数检验Kruskal-Wallis test(不满足方差齐性),利用SPSS 20.0件对各航次日本鳀大小量度指标进行分析[4,11,18-20]。
2结果
2.1 日本鳀不同发育时期卵子大小变化
为比较不同发育阶段的卵子大小差异,对同一航次同一站位的样品进行分析。5月上旬站位13处于Ⅰ-Ⅴ期的卵子总共4011粒,随机选取270粒,分析不同分期的卵子大小。方差分析表明,处于不同发育期的卵子大小量度指标无显著性差异(面积,P>0.05,F4,265=1.780;长径,P>0.05,F4,265=0.534;短径,P>0.05,F4,265=0.129;长径/短径,P>0.05,F4,265=0.265),因此处于不同发育期的卵子可以合并分析(见表2)。

表2 不同发育期日本鳀卵子长径、短径、面积和长径/短径
Note: ①Developmental stage; ②Longer axis; ③Shorter axis; ④Area; ⑤Ratio of longer axis to shorter axis
2.2 日本鳀卵子大小的空间分布
对各航次的取样站位日本鳀卵子的面积、长径/短径进行分析,结果表明各航次的取样站位间,日本鳀卵子的面积、长径/短径均存在显著性差异(见表3)。
2.3 日本鳀卵子大小随时间的变化
从5月上旬至7月上旬,卵子平均长径为(1.32±0.16)mm,平均短径为(0.68±0.06)mm,平均面积为(0.71±0.14)mm2,长径/短径平均值为1.94±1.15。各航次卵子平均长径范围为1.57~1.22mm,平均短径范围为0.77~0.65mm,平均面积范围为0.95~0.62mm2,长径/短径平均值范围为2.03~1.89,(见表4)。5月上旬至7月上旬,日本鳀卵子长径、短径、面积及长径/短径总体上呈逐渐减小的趋势(见图3)。方差分析结果表明长径(H=764.16,P<0.05)、短径(H=682.61,P<0.05)、面积(H=810.55,P<0.05)及长径/短径(H=196.23,P<0.05)各航次间差异显著,变化趋势见图3。
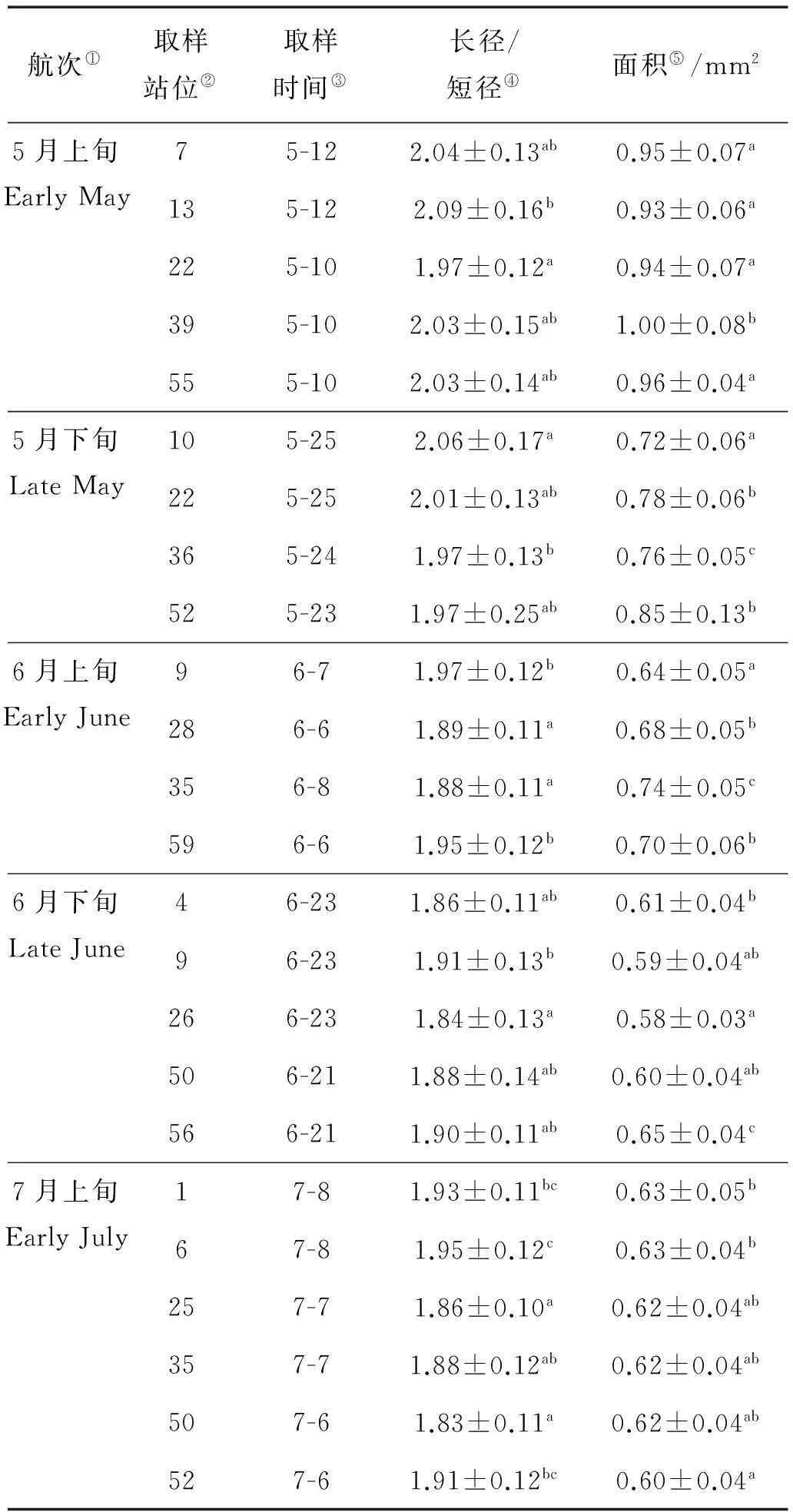
表3 日本鳀卵子面积和长径/短径
注:各航次的同一列中标有不同字母上标的数值相互之间差异显著(P<0.05)。
Note: Values sharing the same letters in the same column of each cruise indicate insignificant difference among groups (P<0.05).
①Cruise;②Sampling station;③Date of survey;④Ratio of longer axis to shorter axis;⑤Area

表4 水温与日本鳀卵子长径、短径、面积和长径/短径

图3 各航次卵子长径、短径、长径/短径及面积
山东半岛南部产卵场日本鳀胚胎发育的阈温度为10.26℃[21],本研究海区各航次取样站位的水温均高于10.26℃,卵子可进行胚胎发育。通过对各航次取样站位的水温分析发现,5月上旬至7月上旬的水温逐渐上升,各航次卵子面积与其对应水温呈显著负相关,其线性回归方程为:
y=1.37-0.035x,(r2=0.90,P<0.01)。
3讨论
本文研究结果表明,从5月上旬至7月上旬,随着水温的升高,日本鳀卵子的所有量度指标均呈逐渐减小的趋势,各航次间卵子各量度指标均存在显著差异。这与其他鱼类的相关研究结果,如加拿大东南部沿海大西洋鳕鱼(Gadusmorhua)[5,22]和智利中北部沿海秘鲁鳀(Engraulisringens)[20,23]等类似。
海洋鱼类的卵径变化受很多因素制约,包括水温、年龄结构、种群栖息地、纬度、产卵期时间、产卵批次和不同批次间的影响等[24]。随着温度升高,鱼类卵子体积(卵径)与温度往往呈负相关关系[19,22],造成上述现象的原因可能与亲鱼繁殖期的推移,储备能量减少有关[25]。黄海区日本鳀亲鱼脂肪储备从5—7月持续下降[26],这可能是造成日本鳀卵径逐渐减小的一个原因。关于卵径随着产卵期推进而减小的这种季节趋势的另一个原因可能是产卵群体年龄结构的变化[27]。在山东半岛南部产卵场,产卵前期以2、3龄日本鳀剩余产卵群体为主,产卵中期则以1龄早生产卵群体和2龄产卵群体为主,而产卵后期及末期主要为1龄晚生补充产卵群体[28]。研究表明,对银大麻哈鱼(Oncorhynchuskisutch)[4]和大麻哈鱼(Oncorhynchusketa)[29]的同批次卵子而言,产卵亲鱼个体体长值越大,卵子卵径就越大;在产卵期的不同阶段,进入山东半岛南部海域产卵场中心产卵的鳀鱼生殖群体全长优势组成存在明显的结构差异,日本鳀生殖个体在产卵前期、中期及后期的平均全长逐渐减小,产卵前期的生殖个体平均全长12.51cm,产卵中期平均全长为11.93cm,产卵后期平均全长为10.84cm[28],因而对日本鳀而言,繁殖季节中日本鳀生殖个体的全长变化亦可能是卵子大小变动的潜在原因。
日本鳀作为广布种,不同海区、不同产卵群体和不同批次的卵子卵径可能存在差异。采用单样本t检验对本次研究结果与以往的研究结果[11]比较,5月上旬日本鳀样品卵子面积显著大于2007年5月7—14日中国东海沿海的日本鳀卵子面积(t=62.73,df=209,检测值testvalue=0.633,P<0.01),也显著大于闽南-台湾浅滩渔场1973年4月—1977年3月采集的日本鳀卵子面积(t=61.75,df=209,检测值testvalue=0.638,P<0.01),略小于1958年日本近海采集的日本鳀卵子面积;5月下旬样品卵子面积显著大于1963年5月14日—6月3日黄海胶州湾的日本鳀卵子面积(t=9.551,df=162,检测值testvalue=0.705,P<0.01);6月下旬样品卵子面积显著小于2003年6月16—29日山东半岛南部产卵场中的日本鳀卵子面积(t=-13.51,df=249,检测值testvalue=0.643,P<0.01)(见表5)。这与其它鱼种的类似研究中,卵子大小随空间环境变化的一般趋势相符,笔者认为其反映了鱼种对高强度捕捞压力及不同产卵场环境的生物学适应[33]。

表5 其他海域日本鳀卵子的大小比较
Note: ①Coastal waters of the East China Sea; ②Fishing ground of the south Fujian-Taiwan Bank; ③Jiaozhou Bay in Yellow Sea; ④Japanese coastal waters; ⑤Spawning grounds off south Shandong Peninsula
随着纬度的升高,成鱼必须要抵抗更短的生长季和更长的冬季,与低纬度相比,体型可能偏小[34]。生长率和体长的差异与纬度的相关性在生活史第一年尤其明显,仔鱼体长的增加通常会提高其第一年冬季的成活率,最终影响到种群的补充情况[35]。研究表明,鳀科鱼类卵子卵径的变化原因包括亲鱼生殖能量的变化、生殖群体年龄组成的变化以及雌性卵细胞的形成过程等[24,36]。卵子大小在本研究海域相对较小的空间尺度范围内亦呈现出一定的时空异质性特征,其具体成因,以及不同海区、环境和季节下日本鳀卵子大小的变动规律、其对环境的响应机制和对种群资源动态的影响等尚需以后进一步深入研究。
参考文献:
[1]Wu Peiqiu, Wu Hezhou, Yang Jiming. Productive ecoogy of fishes in jiaozhou bay waters [J]. Chinese Journal of Oceanology and Limnology, 1989, 7(3): 184-195.
[2]牛翠娟, 娄安如, 孙儒泳, 等. 基础生态学[M]. 第2版. 北京: 高等教育出版社, 2007: 95-108.
Niu C J, Lou A R, Sun R Y, et al. Fundamentals of Ecology[M]. Beijing: Higher Education Press, 2007: 95-108.
[3]Conover D. The relation between capacity for growth and length of the growing season: evidence for and implications of counter gradient variation [J]. Transactions of the American Fisheries Society, 1990, 119(3): 416-430.
[4]Fleming I A, Gross M. Latitudinal clines: a trade-off between egg number and size in pacific salmon [J]. Ecology, 1990, 71(1): 1-11.
[5]Kjesbu O S, Solemdal P, Bratland P, et al. Variation in annual egg production in individual captive Atlantic cod (Gadusmorhua)[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53: 610-620.
[6]Craig P C. Identification of sockeye salmon (Oncorhynchusnerka) stocks in the Stikine River based on egg size measurements[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42: 1696-1701.
[7]Whitehead P J P, Nelson G J, Wongratana T. FAO species catalogue. Clupeoid fishes of the world (SuborderClupeoidei). An annotate and illustrated catalogue of the herring, sardine, pilchards, sprats, shads, anchovies and wolf-herrings. Part 2. Engraulidae[M]. Rome: FAO Fisheries Synopsis, 1988.
[8]唐启升, 苏纪兰. 中国海洋生态系统动力学研究I[M]. 北京: 科学出版社, 2000: 1-252.
Tang Q S, Su J L. Study on China ocean ecosystem dynamics I [M]. Beijing: Science Press, 2000: 1-252.
[9]朱德山, Iversen A. 黄、东海鳀鱼及其他经济鱼类资源声学评估的调查研究[J]. 海洋水产研究, 1990, 11: 1-141.
Zhu D S , Iversen S A. Research on acoustic assessment of anchovyEngraulisjaponicusand other commercial fish in Yellow Sea and East China Sea[J]. Marine Fisheries Research, 1990, 11: 1-141.
[10]李富国. 黄海中南部鳀鱼生殖习性的研究[J]. 海洋水产研究, 1987(8): 41-50.
Li F G . Research on reproductive characteristics of anchovyEngraulisjaponicusin the central and southern Yellow Sea[J]. Marine Fisheries Research, 1987(8): 41-50.
[11]Wan R J, Bian X D . Size variability and natural mortality dynamics of anchovyEngraulisjaponicuseggs under high fishing pressure [J]. Marine Ecology Progress Series, 2012, 465: 243-251.
[12]国家质量技术监督局. GB 12763. 6-91 海洋调查规范[M]. 北京: 中国标准出版社, 1991.
General Administration of Quality Supervision, Inspection and Quarantine of China. GB 12763. 6-91. The specification for oceanographic survey[M]. Beijing: Standards Press of China, 1991.
[13]赵传絪, 张仁斋.中国近海鱼卵与仔鱼[M]. 上海: 上海科学技术出版社, 1985: 182.
Zhao C Y, Zhang R Z. China offshore fish eggs and larvae[M]. Shanghai: Shanghai Science and Technology Press, 1985: 182.
[14]Shao K T. Identification guide of fish eggs in Taiwan Waters[M]. Taipei: Institute of Zoology Academic Sinica, 2001.
[15]Thompson B M, Riley J D. Egg and larval development studies in the North Sea cod (GadusmorhuaL. ) [J]. Rapports et Procès-verbaux des Réunions Conseil international pour l’Exploration de la Mer, 1981, 178: 553-559.
[16]Imai Ch, Tanaka S. Effect of sea water temperature on egg size of Japanese anchovy [J]. Nippon Suisan Gakkaishi, 1987, 53(12): 2169-2178.
[17]Fisher W Huevos. Criasy primeras prelarvas de la anchovetaEngraulisringensJenyns[J]. Rev Biol Mar, 1958, 8(1-2-3): 111-124.
[18]Landaeta M F, Castro L R. Seasonal and annual variation in Chilean hakeMerlucciusgayispawning locations and egg size off central Chile [J]. Progress in Oceanography, 2012, 92-95: 166-177.
[19]Kokita T. Potential latitudinal variation in egg size and number of a geographically widespread reef fish, revealed by common-environment experiments [J]. Marine Biology, 2003, 143: 593-601.
[20]Llanos-Rivera A, Castro L R. Latitudinal and seasonal egg-size variation of the anchoveta (Engraulisringens) off the Chilean coast [J]. Fishery Bulletin, 2004, 102: 207-212.
[21]万瑞景, 赵宪勇. 山东半岛南部产卵场鳀鱼的产卵形态Ⅱ. 鳀鱼的产卵习性和胚胎发育特性[J]. 动物学报, 2008, 54(6): 988-997.
Wan R J, Zhao X Y. Spawning ecology of the anchovyEngraulisjaponicusin the spawning ground of the Southern Shandong Peninsula Ⅱ. Spawning habits and embryonic developmental characteristics of the anchovy[J]. Acta Zoologica Sinica, 2008, 54(6): 988-997.
[22]Chambers R C, Waiwood K. Maternal and seasonal differences in egg sizes and spawning characteristics of captive Atlantic cod,Gadusmorhua[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53: 1986-2003.
[23]Castro L R, Claramunt G, Krautz M C, et al. Egg trait variations in anchovetaEngraulisringens: A maternal effect to changing environmental conditions in contrasting spawning habitats[J]. Marine Ecology Progress Series, 2009, 381: 237-248.
[24]Chambers R C. Environmental influences on egg and propagule sizes in marine fishes[M]//Early Life History and Recruitment in Fish Populations. Berlin: Springer Netherlands, 1997: 63-102.
[25]Rass T S. Biogeographic rule of inverse relation between egg size and environmental temperature in poikilothermous animals [J]. Trudy IOAN, 1986, 116: 152-168.
[26]赵宪勇. 黄海鳀鱼种群动力学特征及其资源可持续利用[D]. 青岛: 中国海洋大学, 2006.
Zhao X Y . Population Dynamics Characteristics and Sustainable Utilization of the Anchovy Stock in the Yellow Sea[D]. Qingdao: Ocean University of China, 2006.
[27]Simpson A C. The spawning of the plaice (Pleuronectesplatessa) in the North Sea[M]// Fishery Investgation. London: HM Stationery Office, 1959, 12: 1-111.
[28]李显森, 赵宪勇, 李凡, 等. 山东半岛南部产卵场鳀鱼生殖群体结构及其变化[J]. 海洋水产研究, 2006, 27(1): 46-53.
Li X S, Zhao X Y, Li F, et al. Structure and its variation of the anchovy (Engraulisjaponicus) spawning stock in the Southern waters to Shandong Peninsula[J]. Marine Fisheries Research, 2006, 27(1): 46-53.
[29]Beacham, T D, Murray C B. Effect of female size, egg size, and water temperature on developmental biology of chum salmon (Oocorhynchusketa) from the Nitinat River, British Columbia[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42: 1755-1765.
[30]江素菲, 郑小衍. 闽南-台湾浅滩渔场鳀鱼卵和仔鱼的形态特征及其产卵场[J]. 台湾海峡, 1984, 3(2): 224-231.
Jiang S F, Zheng X Y. The morphological characters of eggs and larvae ofEngraulisjaponicusTemminck et Schlegel and its spawning area in the fishing ground of the south Fujian and Taiwan Bank[J]. Taiwan Strait, 1984, 3(2): 224-231.
[31]Ruan H C. Studies on the eggs and larvae ofEngraulisjaponicun[J]. Stud Mar Sin, 1984, 22: 29-56.
[32]Uchida K, Imai S, Mito S, et al. Studies on the eggs, larvae and juvenile of Japanese fishes[M]// Series 1. Fukuoka in Japanese: Second Laboratory of Fisheries Biology, Fisheries Department, Faculty of Agriculture. Fukuoka: Kyushu University, 1958: 17-18.
[33]Thorson G. Reproductive and larval ecology of marine invertebrates [J]. Biological Review, 1950, 25: 1-45.
[34]Mousseau T A. Ectoderms follow the converse to Bergmann’s Rule [J]. Evolution, 1997, 51: 630-632.
[35]Garvey J E, Wright R A, Stein R A. Overwinter growth and survival of age -0 largemouth bass: Revisiting the role of body size [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55: 2414-2424.
[36]Blaxter J H S, Hunter J R. The biology of clupeoid fishes [J]. Adv Mar Biol, 1982, 20: 3-194.
责任编辑朱宝象
Spatial and Temporal Changes of the Egg Size of Japanese Anchovy (Engraulisjaponicus)Inhabiting the Near Shore Spawning Ground of Central Yellow Sea
LI Xiu-Mei1, YE Zhen-Jiang1, LI Zeng-Guang1, HU Hai-Sheng1, ZOU Quan1, YU Hong-Liang2
(1.College of Fisheries, Ocean University of China, Qingdao 266003, China; 2.Bureau of Oceanic and Fishery of Wendeng District, Weihai 264400, China)
Abstract:The Japanese anchovy Engraulis japonicas is widely distributed along the western North Pacific, such as eastern China Sea, Yellow Sea, Japan Sea, etc. and it has also supported one of the largest fisheries in China over the last three decades with the peak yield of more than 1 million tons in 1990s. However, there are few studies for the early life stages of this species in North Pacific, especially in near shore spawning ground of central Yellow Sea. Egg size of fish is not only easy to quantify, but also often reflects the amount of resources mothers invest in individual offspring. It is treated as one structural element with which to characterize a species’ ontogeny regardless of causal relationships between the species and its habitat. It is therefore that egg size often highly varies both between different species and between stocks of a species, even between clutches of the same individual. As shown by numerous publications dealing with conditions of life and reproduction of fishes, egg size of fish is an important biologi cal parameter characterizing the environment of spawning ground and fish reproductive traits. This study were aimed to as follows: 1) report changes in egg size throughout the Japanese anchovy spring and summer spawning season as well as the spatial characteristic of egg size in near shore waters of central Yellow Sea; 2) evaluate whether egg size varies with respect to space and time, and 3)provide data for studies on its reproductive traits and resources variation mechanism. Six ichthyoplankton cruise surveys were conducted in < 20m depth waters of central Yellow Sea from late April to early July, 2013 with the time interval of half a month. We collected Japanese anchovy eggs from 75 stations along the coastal zone(34.5°N~35.5°N, 119.0°E~120.5°E). This area is one important part of the spawning waters of Japanese anchovy. Eggs were collected with a plankton net(500μm mesh). Upon collection, all eggs were preserved in 5% buffered formalin and then transferred to the lab after the survey. Egg size(length, width, area) was measured by software Image-Pro Plus 6.0 based on the images captured on a dissecting microscope and camera(Nikon SMZ1500). Based on data from the six cruise surveys conducted, the temporal and spatial changes in the morphology and size of Japanese anchovy eggs were investigated. The results indicated that the morphology of elliptical anchovy eggs was similar among anchovy eggs at different developmental stages. However, egg size varied temporally and spatially. Egg size decreased generally from early May to early July, which was negatively correlated with water temperature. The eggs began to occur in the study area in early May with largest longitudinal section area of (0.95±0.07)mm2(longer axis (1.57±0.09)mm and shorter axis (0.77±0.04)mm). Minimum egg size occurred in late June (longitudinal section area (0.61±0.43)mm2, longer axis (1.20±0.07)mm and shorter axis (0.64±0.03)mm). Compared with the documented previously, Japanese anchovy eggs from early May in the currently studied area were significantly larger than those collected in coastal waters of East China Sea, and anchovy eggs from late June were significantly smaller than those collected at the spawning grounds off south Shandong Peninsula. These results is consist with the popular science regularity that egg size varies among different spawning grounds and decreases during the whole spawning season. In the meanwhile, the phenomenon that there were some spatial egg size varies to be founded between positions in so small areas would be paid more attention to it.
Key words:Central Yellow Sea; Haizhou Bay; Japanese anchovy; egg size; temporal and spatial variation
DOI:10.16441/j.cnki.hdxb.20140331
中图法分类号:Q948.3
文献标志码:A
文章编号:1672-5174(2016)02-054-07
作者简介:李秀梅(1989-),女,硕士,主要从事鱼类早期发育研究。E-mail:xiuxiu12345@126.com❋❋通讯作者: E-mail:yechen@ouc.edu.cn
收稿日期:2014-10-16;
修订日期:2015-05-07
基金项目:❋ 海洋公益性行业科研专项经费项目(201305030);高等学校博士学科点专项科研基金项目(20120132130001);中央高校基本科研业务费专项(201262004)资助
引用格式:李秀梅,叶振江,李增光,等. 黄海中部近岸产卵场日本鳀卵子大小的时空变化[J].中国海洋大学学报(自然科学版), 2016, 46(2): 54-60.
Supported by Public Science and Technology Research Funds Projects of Ocean (201305030); Specialized Research Fund for the Doctoral Program of Higher Education (20120132130001); Fundamental Research Funds for the Central Universities (201262004)
猜你喜欢
南水北调与水利科技(2017年1期)2017-02-27
中国人口·资源与环境(2016年7期)2016-10-31
商(2016年29期)2016-10-29
商场现代化(2016年12期)2016-06-06
安徽农学通报(2015年10期)2015-06-15
农业科技与装备(2014年11期)2015-02-02
农业科技与装备(2014年9期)2014-12-26
