
中国特有濒危植物翅果油树的SSR引物开发及特性
2016-03-24 06:32叶占洋王兆山李云晓饶国栋张建国
西北植物学报 2016年2期
叶占洋,王兆山,3,李云晓,饶国栋,3,张建国,3*
(1 中国林业科学研究院 林业研究所 林木遗传育种国家重点实验室,北京 100091;2 中国林业科学研究院 林业研究所 国家林业局林木培育重点实验室,北京 100091;3 南京林业大学 南方现代林业协同创新中心,南京 210037)
中国特有濒危植物翅果油树的SSR引物开发及特性
叶占洋1,2,王兆山1,2,3,李云晓1,2,饶国栋1,2,3,张建国1,2,3*
(1 中国林业科学研究院 林业研究所 林木遗传育种国家重点实验室,北京 100091;2 中国林业科学研究院 林业研究所 国家林业局林木培育重点实验室,北京 100091;3 南京林业大学 南方现代林业协同创新中心,南京 210037)
摘要:翅果油树是中国特有的濒危植物,利用磁珠富集法开发适用于翅果油树的SSR引物,并在2个翅果油树群体进行多态性验证。结果表明,筛选的20对引物中有19对是多态且选择中性的SSR引物,其中引物CGY19表现单态,仅有一个位点显示Hardy-Weinberg平衡的偏离(P<0.01),在2个群体中均未检测到连锁不平衡现象;每个位点的等位基因数为1~9个,平均为4.225个,观测杂合度和期望杂合度的变化范围分别为0~0.900和0~0.847。利用开发的19对多态性SSR引物对2个翅果油树自然群体的遗传多样性水平分析结果表明,其平均遗传多样性水平为97.37%,说明翅果油树濒危的原因并不是遗传多样性水平丧失;遗传分化系数FST为0.033 1,表明遗传变异主要存在于种群内,这可能是由于种群之间的基因流(Nm=32.081 8)较大的结果,这也是2个翅果油树种群遗传相似度高的原因。这些特异性引物的开发和遗传多样性的研究将对翅果油树的交配系统、遗传结构的研究起到重要的作用,并为翅果油树种质资源的取样策略、保护和利用提供有效的信息。
关键词:翅果油树;微卫星引物;遗传结构;保护和利用
ElaeagnusmollisDiels,an endangered shrub or small deciduous tree,is endemic to Lüliang and Qinling Mountains area of China[1].The seeds ofE.molliscontain abundant nutritional ingredients such as fatty acid,VC,VEand amino acids.Hence,the fact sheds light on the unquestionable fact thatE.mollishas become a very precious yet rare wild woody oil tree species with high value in economy and medicine[2].Furthermore,E.mollisis also a good greening tree species in the Northwest of China due to its outstanding characteristics of drought,cold and barren resistance,rapid growth as well as developed root system,which are helpful for windbreak and sand-fixation[3-4].However,its distribution range is limited to a small area with a small population size and it has been listed as a rare and endangered species in China.To understand the population genetics and evolutionary history of this species is of great importance to take concrete measures to conserve and effectively utilize its germplasm resource.Although 10 pairs of microsatellite markers forE.mollishave been developed,they were all selected from theHippophaerhamnoidesmarkers which were selected fromVitisviniferawithout verification via clone sequencing[5-6].Biased even wrong conclusions would be obtained if using these markers in genetic studies becauseVitisviniferaandE.mollisshare few common features.Based on the above detailed analysis,we reported a set of novel high-polymorphic microsatellite markers forE.mollis,and the markers could be employed to inspect the genetic diversity level of natural populations ofE.mollis.To close,this paper possesses great reference value for studying its population genetics structure,analyzing the reasons of endangerment and providing valuable information for its conservation.
1Material and methods
1.1Biological material and DNA extraction
A total of 96 individuals of 2 separated populations (40,56 individuals from NLXKM,AQ,respectively) were collected from Yicheng,Shanxi Province,China,in 2014,and the leaves were dried with silica gel.Total genomic DNA was extracted from these dried leaves using the CTAB method[7]with some modifications and Plant Genomic DNA Kit (TIANGEN Biotech (Beijing) Co.,Ltd).DNA quantity was detected by agarose gel electrophoresis.
1.2Marker procedure
RsaⅠ andXmnⅠ were applied to digest total DNA (New England Biolabs,Ipswich,MA) in a 20 μL reaction volume overnight at 37 ℃ and then the fragments were ligated to the double-strand Super SNX-24 linker (forward:5′-GTTTAAGGCCTAGCTAGCAGAATC-3′;reverse:5′-pGATTCTGCTAGCTAGGCCTTAAACAAA-3′).The ligated DNA fragments were hybridized with two kinds of single-strand bio-tinylated microsatellite probes [5′-(CA)15-Biotin,5′-(GA)15-Biotin] whose products were captured by streptavidin-coated paramagnetic beads (Dynal Biotech Dynabeads M-280 Streptavidin,Oslo,Norway) and gathered with a magnetic particle concentrator (MPC,Dynal Biotech Dynal MPC-S,Dynal,Oslo,Norway).After that,the enriched DNA was amplified via polymerase chain reaction (PCR) with super SNX-24 linker-forward as a primer,and the purified PCR products were randomly ligated into the pGEM-T easy vector (Promega Corp.,Madison,Wisconsin,USA) and then transformed intoEscherichiacoli(TransGen Biotech,Beijing,China) Top 10 competent cells.
Seventy-two positive clones were selected randomly and sequenced orderly,and 56 (78%) reads were tested containing microsatellite motifs.Of which,40 sequences were selected for primer designing using Primer premier 5.0 software (Premier Biosoft International,Silicon Valley,California,USA).Finally,20 pairs of primers (Table 1) were chosen because their PCR products showed single and clear bands in the agarose gel electrophoresis.
1.3Amplification and sequence
One of the fluorescent dyes (FAM and HEX) was labeled to the forward primer for polymorphism detection,and the following PCR was performed in 20 μL reaction system containing 10-50 ng of DNA template,2 μL of 10×PCR buffer (Mg2+Plus),1.6 μL of 2.5 mmol/L dNTP mixture,0.8 μL of 10 μmol/L each primer,and 0.1 μL of 5 U/μL rTaqDNA polymerase (TaKaRa,Dalian,Liaoning,China).Amplification was carried out in a Master Thermal Cycler (Applied Biosystems,Foster City,California,USA) as follows:4 min at 94 ℃,followed by 35 cycles of 30 s at 94 ℃,30 s at Ta,90 s at 72 ℃ and a final extension at 72 ℃ for 10 min.The PCR products were detected by an ABI 3 130 xl genetic analyzer alongside the GeneScan 500 LIZ size standard (Applied Biosystems,Foster City,California,USA) and the microsatellite profiles were analyzed by GeneMapper version 4.0(Applied Biosystems).

Table 1 Characteristics of 20 polymorphic microsatellite markers in E.mollis
1.4Data analysis
Forty individuals from populations NLXKM and AQ (20 individuals per population) were aided to detected the polymorphism of these markers using Arlequin software Version 3.5.1[8]with the following parameters:the departures from Hardy-Weinberg equilibrium (H-W) and the gametic linkage disequilibrium between pairs of loci (LD).Then the polymorphic SSR markers were employed to analyze the genetic diversity of the two natural populations ofE.mollis.These parameters encompassed:the genetic diversity level (P),the mean number of alleles (Na),the effective number of alleles (Ne),the observed and expected heterozygosity (HOandHE),the Shannons information index (I),the Fixation index (F),the gene flow (Nm),and the significant fragmentation (FST) were analyzed using the computer program GenAlEx 6.5[9].
2Results and analysis
The mean number of alleles ranged from 1 to 9,with an average of 4.225.The observed and expected heterozygosity per locus ranged respectively from 0 to 0.900 and from 0 to 0.847 with averages of 0.462 and 0.516 (Table 2).The locus (CGY19) was monomorphic in both of the two populations.For the author’s part,it should be avoided while studying the indigenous populations in future.
Among 37 tests of H-W expectations,1 locus (CGY3) in AQ population and 2 loci (CGY3,CGY4) in NLXKM population showed significant departures from H-W (P<0.01),but only 1 locus (CGY3) showed significant deviation from H-W (P<0.01) in both of the two populations,which was probably caused by the influence of selection.Thus,this locus should be avoided in future studies.
Tests of linkage disequilibrium for 380 pairs of loci manifested that 1 pair of loci in AQ population and 4 pairs of loci in NLXKM population exhibited linkage disequilibrium,but none of them existed in the two populations simultaneously.Therefore,the conclusion can be drawn that they are probably not a consequence of physical linkage.
Nineteen pairs of polymorphic SSR markers obtained from our research were utilized to analyze the genetic diversity level of the two natural populations ofE.mollis.The result indicated that all the four populations ofE.mollishad an average high level of genetic diversity based on the percentage of Polymorphic Loci (P(mean) = 97.37%),and a low coefficient of gene differentiation (FST=0.033 1) between populations,which revealed that the genetic variation mainly existed within the populations and primarily was due to the frequent gene flow between populations (Nm=32.081 8).In this vein,it accounted for the reason why the Shannon’s information index (I=genetic similarity) was very high in each population.The mean fixation indexes (F) of the two populations was 0.061 9 (SE=0.036),which showed that the populations of this species slightly deviated from balance and had surplus homozygotes at the population level (Table 3).
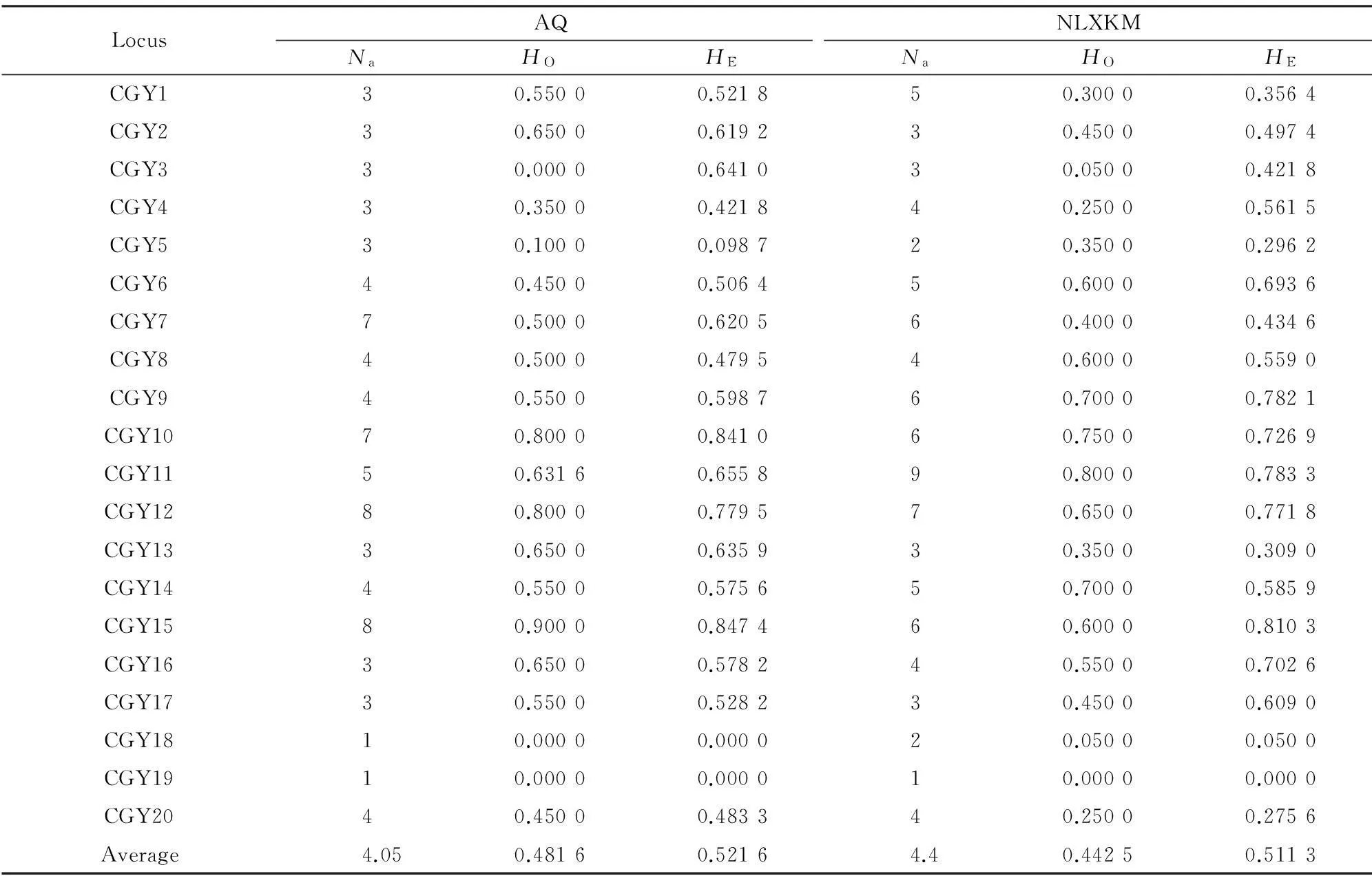
Table 2 Results of initial primer screening in E.mollis(N=20)
Notes:Na.Observed number of alleles;HO.Observed heterozygosity;HE.Expected heterozygosity.
The endangered mechanism of plant is mainly derived from the two factors both inside and outside of species.The internal causes include the plant inheritance,physiology,development,reproduction and so on,whereas the external factors can be attributed to human disturbance,ecological environment change,etc[10].At the species level,the result showed that the genetic diversity indexes ofE.mollis(Na=5.105 3,HO=0.515 9 andHE=0.552 1) were not much different from the other two species in the same family (Russian olive:Na=4.5,HO=0.465 5,HE=0.548;Hippophae rhamnoides:Na=6.667,HO=0.232 2,HE=0.389 9)[11-12].This result showed that the genetic diversity level ofE.molliswas not low.However,what causes this result is not evident.Two reasons may be illustrated to accounted for this phenomenon:(1)the genetic diversity of existing populations ofE.mollismay inherit from their ancestral population with a rich genetic diversity because of the influence of the Quaternary glaciation and anthropogenic activities;(2)outcrossing which is an important mechanism to maintain high levels of genetic variation within populations may be also a vital reason[13].Form a scientific view,it could be verified by estimating the mating system ofE.mollis.

Table 3 Genetic diversity parameters of two natural populations
Notes:N.No.of samples;P.Genetic diversity/%;Na.Number of alleles;I.Shannons information index;HO.Observed heterozygosity;HE.Expected heterozygosity;F.Fixation index;FST.Coefficient of gene differentiation;Nm.Gene flow.
As has shown from the previous analysis,the level of its genetic diversity may not be the main endangering factor.As Wang and Qin said,the loss of genetic diversity ofE.molliswas more likely to be the end result of its endangerment rather than the cause of the endangerment[14].Moreover,the successful cultivation ofE.mollisin many other areas implied that ecological environment change was also not the main endangering factor[15].During two times field surveys,we got the insight that the germination (no more than 30%) and seed setting rate were extremely low,which may result from inbreeding depression i.e.,the increased homozygosities of recessive genes caused by inbreeding will result in high mortality of the embryos.A low coefficient of gene differentiation (FST=0.033 1,SE=0.007) was detected between the two populations and it showed that genetic variation mainly exist within the populations.These findings will facilitate and promote the following studies in sampling strategies,conservation and utilization of germplasm genetic resources,especially for endangered species.
3Conclusions
In this study,we provided 20 nuclear microsatellite markers developed specifically fromE.mollis,most of which showed highly polymorphic,and were selectively neutral.These markers will be useful in our further studies on mating system,genetic structure and gene flow ofE.mollis.If properly used and studied,it may provide valuable information for the recovery and conservation of this endangered species.
The genetic diversity level ofE.mollisobtained in our study was not low.Thus,it falsified the previous wrong perception and proved that this was not the endangered reason of this species.Given the fact that the genetic variation mainly exist within the populations,more individuals should be sampled within populations in the further research on mating system ofE.mollisto understand the reproductive status of this species,which can be used as a ladder to further explore the effective conservation and genetic improvement strategies.
Acknowledgements:We thank MING Wen and ZHANG Chaodong in QHD TIANYA Agriculture and Forest Technique Co.,Ltd for the sampling of biological material.Financial support for this research was provided by corporation project on science and postgraduate education in Beijing and the Collaborative Innovation Plan of Jiangsu Higher Education.
References:
[1]上官铁梁,张峰.我国特有珍稀植物翅果油树濒危原因分析[J].生态学报,2001,21(3):502-505.
SHANGGUAN T L,ZHANG F.The endangered causes ofElaeagnusmollis,an endemic to China[J].ActaEcologicaSinica,2001,21(3):502-505.
[2]冯保英.翅果油树种子化学成分的研究[J].山西林业科技,1989,4(6):6-9.
[3]董志,张飞云,闫桂琴.濒危植物翅果油树的研究进展及其开发前景[J].首都师范大学学报(自然科学版),2005,26(3):65-67.
DONG ZH,ZHANG F Y,YAN G Q.Advances in research on the endemic and endangered plant ofElacagnusmollisand its exploitation prospects[J].JournalofCapitalNormalUniversity(Nat.Sci.Edi.),2005,26(3):65-67.
[4]庞晓慧,张华新,刘涛.我国特有珍稀植物翅果油树开发利用的探讨[J].林业资源管理,2006,3:15.
PANG X H,ZHANG H X,LIU T.Study on development and use ofElaeagnusmollis,an endemic species to China[J].ForestResourcesManagement,2006,3:15.
[5]QIN Y Y,WANG Y L,ZHANG Q D,etal.Analysis on the population genetic diversity of an endangered plant (Elaeagnusmollis) by SSR markers[J].JournalofWuhanBotanicalResearch,2010,28(4):466-472.
[6]孙燕琳,阮成江,金华.沙棘SSR分子标记的开发[J].安徽农业科学,2007,35(1):45-46.
SUN Y L,RUAN C J,JIN H.Exploitation of SSR marker ofHippophaerhamnoides[J].JournalofAnhuiAgriculturalSciences,2007,35(1):45-46.
[7]DOYLE J J.A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J].PhytochemBull,1987,19:11-15.
[8]EXCOFFIER L,LISCHER H E.Arlequin suite ver 3.5:a new series of programs to perform population genetics analyses under Linux and Windows[J].MolecularEcologyResources,2010,10(3):564-567.
[9]PEAKALL R,SMOUSE P E.GenAlEx 6.5:genetic analysis in Excel.Population genetic software for teaching and research-an update[J].Bioinformatics,2012,28(19):2 537-2 539.
[10]洪德元,葛颂,张大明,等.植物濒危机制研究的原理和方法[C]//林业部野生动物和森林植物保护司.生物多样性研究进展.北京:中国科学技术出版社,1995:125-133.
[11]GASKIN J F,HUFBAUER R A,BOGDANOWICZ S M.Microsatellite markers for Russian olive (Elaeagnusangustifolia;Elaeagnaceae)[J].ApplicationsinPlantSciences,2013,1(9):1300013.
[12]WANG A,ZAHNG Q,WAN D S,etal.Nine microsatellite DNA primers forHippophaerhamnoidesssp.sinensis(Elaeagnaceae)[J].ConservationGenetics,2008,9(4):969-971.
[13]万开元,陈防,李作洲,等.珍稀植物濒危机制及保育策略中的营养条件[J].生态环境,2004,13(2):261-267.
WAN K Y,CHEN F,LI Z Z,etal.The nutrition condition of endangered mechanism and conservation strategr of rare plants[J].EcologyandInvironmentalSciences,2004,13(2):261-267.
[14]WANG Y L,QIN Y Y,DU Z,etal.Genetic diversity and differentiation of the endangered treeElaeagnusmollisDiels (ElaeagnusL.) as revealed by simple sequence repeat (SSR) markers)[J].BiochemicalSystematicsandEcology,2012,40:25-33.
[15]ZHANG F,HAN S Q,SHANGGUAN T L.Analysis on relationship between geographic distribution ofElaeagnusnollisand eco-environment factors in China[J].JournalofShanxiUniversity(Nat.Sci.Edi.),2001,24(1):86-88.
(编辑:宋亚珍)
Development and Characterization of Microsatellite Markers in theElaeagnusmollisDiels
YE Zhanyang1,2,WANG Zhaoshan1,2,3,LI Yunxiao1,2,RAO Guodong1,2,3,ZHANG Jianguo1,2,3*
(1 State Key Laboratory of Tree Genetics and Breeding,Research Institute of Forestry,Chinese Academy of Forestry,Beijing 100091,China;2 Key Laboratory of Tree Breeding and Cultivation,State Forestry Administration,Research Institute of Forestry,Chinese Academy of Forestry,Beijing 100091,China;3 Collaborative Innovation Center of Sustainable Forestry in Southern China,Nanjing Forestry University,Nanjing 210037,China)
Abstract:Elaeagnus mollis Diels is an endemic and endangered tree in China.Twenty primer pairs were selected by the combined biotin capture and identified in two populations of E.mollis.Nineteen pairs showed highly polymorphic,and were selectively neutral and the locus CGY19 was monomorphic.One locus manifested significant deviation from Hardy-Weinberg Equilibrium (P<0.01),which was probably caused by the influence of selection.No significant linkage disequilibrium was detected among pairs of loci in each pair.With a mean of 4.225,the number of alleles per locus ranged from 1 to 9.The observed and expected heterozygosity (HOand HE) per locus ranged from 0 to 0.900 and from 0 to 0.847,respectively.The genetic diversity levels of 2 natural populations of E.mollis were estimated by these 19 pairs of SSR markers obtained in this study.It showed that the mean genetic diversity was 97.37%,indicating that the loss of genetic diversity was not the cause of the endangerment of E.mollis.The significant fragmentation (FST=0.0331) was tested between the two populations and it provided the evidence that the genetic variation mainly existed in the populations.Furthermore,due to the frequent gene flow between populations (Nm=32.081 8),it also verified the Shannon’s information index (I=genetic similarity) was very high in each population.On a long view,the new set of loci and the information of genetic diversity will be applied in the mating system and population structure of this species and provide valuable information for its sampling strategy,conservation and utilization.
Key words:Elaeagnus mollis Diels;microsatellite markers;population structure;conservation and utilization.
中图分类号:Q346+.5;Q789
文献标志码:A
作者简介:叶占洋(1990-),男,硕士,主要研究方向为分子生态学。E-mail:1073021320@qq.com*通信作者:张建国,研究员,主要从事人工林定向培育研究工作。E-mail:zhangjg@caf.ac.cn
基金项目:中央级公益性科研院所基本科研业务费专项资金(CAFYBB2016ZX003-1)
收稿日期:2015-10-13;修改稿收到日期:2015-12-02
文章编号:1000-4025(2015)02-0274-06
doi:10.7606/j.issn.1000-4025.2016.02.0274
