
亚热带马尾松林恢复过程中物种丰富度及生物量变化
2016-04-05 08:28高一飞张静唐旭利王万同尹光彩1中国科学院华南植物园广东广州510650中国科学院大学北京10009河南师范大学旅游学院河南新乡5007广东工业大学环境科学与工程学院广东广州510006
生态环境学报 2016年1期
高一飞,张静,唐旭利,王万同,尹光彩1.中国科学院华南植物园,广东 广州 510650;2.中国科学院大学,北京 10009;.河南师范大学旅游学院,河南 新乡 5007;.广东工业大学环境科学与工程学院,广东 广州 510006
亚热带马尾松林恢复过程中物种丰富度及生物量变化
高一飞1,2,张静1,2,唐旭利1*,王万同3,尹光彩4
1.中国科学院华南植物园,广东 广州 510650;2.中国科学院大学,北京 100049;
3.河南师范大学旅游学院,河南 新乡 453007;4.广东工业大学环境科学与工程学院,广东 广州 510006
摘要:马尾松林恢复过程中物种丰富度及生物量变化有助于了解该过程中物种多样性及碳汇功能的变化。选取亚热带东部湿润地区的马尾松(Pinus massoniana)林、针阔叶混交林及常绿阔叶林样地构建马尾松林自然恢复系列,通过比较不同森林中生物量和物种丰富度的变化探讨马尾松林恢复过程中碳汇功能以及物种多样性的变化。依据中国科学院战略性先导科技专项“应对气候变化碳收支认证及相关问题”之森林课题调查数据库中亚热带东部湿润地区的部分数据进行物种多样性和生物量的分析,分别计算不同胸径(diameter at breast height,DBH)等级(1 cm≤DBH<5 cm、5 cm ≤DBH<10 cm、10 cm≤DBH<20 cm、DBH≥20 cm)以及群落总的物种丰富度和生物量,采用Tukey多重比较以及简单线性回归的方法分析不同森林类型之间生物量以及物种丰富度的差异,并探索物种丰富度与生物量之间的关系。结果表明亚热带东部地区的马尾松林、针阔叶混交林以及常绿阔叶林的总平均生物量分别为(114.74±4.82)、(124.99±4.99)、(220.9±8.62)t·hm-2,总物种丰富度分别为(5.81±0.61)、(17.55±1.17)、(23.71±2.04),在马尾松林转变为针阔叶混交林的过程中群落的物种数量显著增加(P=0.000),但碳贮量增加不明显(P=0.305);在针阔叶混交林转变为常绿阔叶林的过程中碳贮量显著增加(P=0.000)且生物量向大个体(DBH≥20 cm)的方向集中,但群落整体的物种数量增加不明显(P=0.159)。总体而言,亚热带地区所选森林中物种丰富度与生物量之间未表现出良好的线性关系,但针阔叶混交林的大个体(DBH≥5 cm)以及3种森林类型的小个体(DBH<5 cm)的物种丰富度与生物量之间呈现显著的线性相关。研究表明,亚热带东部湿润地区森林中小径级的个体生物量随物种丰富度增加其变化趋势明显,增加小个体的物种丰富度既能够促进群落中资源的利用效率,又能够促进森林生态系统的物质循环;针阔叶混交林大个体的生物量随物种丰富度的增加而显著增加,对针阔叶混交林中大个体乔木进行合理的管理也能够促进群落对资源的有效利用。
关键词:马尾松;常绿阔叶林;物种丰富度;生物量;亚热带
GAO Yifei,ZHANG Jing,TANG Xuli,WANG Wantong,YIN Guangcai.Variation of Biomass and Species Richness in Subtropical Forest based on Pinus massoniana Succession [J].Ecology and Environmental Sciences,2016,25(1):22-29.
森林作为陆地生态系统最大的碳汇,在吸收温室气体以及应对气候变化的过程中起着重要作用(Pan et al.,2011)。当前的气候变化日益明显,人们对森林的碳汇功能也愈加重视。速生林或新造林因生长迅速而被认为具有较大的固碳潜力(Intergovernmental Panel on Climate Change,IPCC,2014),因此在水热条件适宜的地方造林已经成为林业固碳增汇的主要经营管理方式。但此类森林在种类组成上往往较为单一,系统应对环境变化的缓冲能力相对较差,能否维持稳定的碳汇功能是值得深入探讨的问题。与新造林或速生林相比,地带性植被类型往往具备较高的物种多样性,系统不仅能够较好地抵御环境变化的影响,还能够维持相对稳定的生产力(Yachi et al.,1999;Hooper et al.,20053-5;Balvanera et al.,2006;Cardinale et al.,2007)。因此,对于陆地生态系统而言,碳汇功能与多样性之间是否存在协同性成为了当前研究的热点问题之一。目前普遍认为由于受补偿效应或取样效应的影响,生态系统中物种数量的增加可以促进植物生物量增长,但多数结论是基于物种添加或去除的控制实验。当对大尺度的自然森林群落进行研究时,较大的空间异质性以及非生物因素可能会影响这种关系(Loreau et al.,2001;Hooper et al.,200519-21)。因此物种多样性与生物量在大尺度下的关系尚需要更多的研究进一步论证(Szwagrzyk et al.,2007555-556;Paquette et al.,2011;Bu et al.,20141)。
马尾松(Pinus massoniana)是中国松属树种中分布最广的一种。由于能够在贫瘠的环境中迅速生长,马尾松通常作为先锋树种被用于造林过程,在中国南方的分布面积达到1.13×107hm-2,占全国总造林面积的20%(罗应华等,2013)。有研究表明中国南方近1/4的马尾松林群落存在不同程度的退化(莫江明等,2004)194;且长期纯林经营还使得马尾松人工林立地衰退,层次结构简单,容易遭受松毛虫侵害,严重威胁着林地的可持续性经营(莫江明等,2004194;张真等2008140-150)。已有研究表明利用演替理论指导进行植被恢复能够改善群落的结构并提高生态系统稳定性(方炜等,199531;王希华等,2001)。由演替初期的马尾松纯林,过渡到以马尾松和阳生阔叶树种构成的针阔叶混交林,再到地带性的常绿阔叶林的群落结构变化被认为是亚热带常绿阔叶林恢复-演替的模式(彭少麟等,1993;方炜等,199531)。随着演替的正向进行,物种多样性因物种组成的改变将发生强烈变化,但森林碳汇在这一过程中有何变化?与群落物种多样性的变化是否一致?这些问题有待深入研究。
针对上述问题,本研究选取位于亚热带东部湿润地区的马尾松林、针阔叶混交林以及常绿阔叶林群落为研究对象,以生物量作为反映森林群落碳汇功能的指标(Fang et al.,2001;Zhao et al.,2005;Xu et al.,2010),以物种丰富度作为反映群落物种多样性的测度指标(马克平,1994162-163),分析生物量以及物种丰富度的变化,探讨二者之间的关系,旨在探讨马尾松林恢复过程中森林碳汇功能以及物种多样性的变化,为马尾松林的合理管理提供参考和建议。
1 材料和方法
1.1研究区概况与样地选取
研究区域包括位于中国亚热带范围内的安徽、福建、广东、广西、贵州、湖北、湖南、江苏、江西、四川、浙江、重庆等12个省份及直辖市,地理坐标为21.85°~33.00°N,106.22°~121.99°E。区内年平均气温介于13~20 ℃,最冷月平均气温在0~15 ℃,年降雨量一般高于1000 mm,最高可达3000 mm以上;植被类型丰富,既有地带性的植被类型常绿阔叶林,又有大量的速生人工林,以及由结构较为单一的人工林向地带性森林转化的过渡性植被类型。马尾松作为中国南方常见的造林树种在中国亚热带东部湿润地区的10省市均有广泛的分布。
本研究选择了中国科学院战略性先导科技专项“应对气候变化碳收支认证及相关问题”(以下简称碳专项)中森林课题调查数据库中亚热带东部湿润地区的部分数据进行分析。碳专项对森林生态系统的调查参照《IPCC优良做法指南》(IPCC,2003)采用网格调查的方法,在全国设置了7200个典型森林生态系统的代表性样地(周国逸等,2015)102-104。根据研究目的,本研究选取了亚热带东部湿润地区12省(直辖市)的马尾松林(109个样地)、包含马尾松的针阔叶混交林(123个样地)、以及地带性植被类型常绿阔叶林(84个样地)为研究对象。尽管部分地区符合条件的调查样地数量较少,但就大尺度的研究而言,调查样地的数量并不存在确定的限制(Zhou et al.,2014),而且碳专项的调查是在全国森林资源分布的基础上进行3%~5%的随机抽样布设的样地,样地具有充分的代表性(周国逸等,2015)102-104。所选取的样地分布如图1所示。
1.2研究方法
1.2.1森林类型以及径级划分
马尾松在马尾松林样地以及针阔叶混交林中的平均相对优势度分别约为91.47%和39.69%,生物量占样地总生物量的平均比例约为92.26%和41.46%。同时选取的地带性顶级演替群落为常绿阔叶林,所选森林类型可以反映由马尾松林自然恢复到针阔叶混交林再到常绿阔叶林的演替过程。
依据个体胸径分布特点将3种森林的个体划分为4个不同胸径等级:径级I(1 cm≤DBH<5 cm)、径级II(5 cm≤DBH<10 cm)、径级III(10 cm≤DBH<20 cm)、径级IV(DBH≥20 cm)。马尾松林样地中为径级III的个体所占比例最高(42.76%±1.99%),径级I的个体所占比例最低(22.48%±5.18%);针阔叶混交林以及常绿阔叶林中径级II的个体所占比例最高(40.58%±1.68%,38.47%±1.95%),而个体所占比例最低的径级分别为IV和I(13.16%±1.03%,22.45%±5.26%,图2)。总体而言,3种森林的个体集中分布在径级II和径级III中,分布在径级I和径级IV中的个体数量较少。
1.2.2生物量计算
本研究采用拟合异速生长方程的方式估算森林生物量,分别估算个体叶、枝、干、根各器官生物量,各器官生物量之和为全株生物量,所选方程形式统一如式1所示。径级的单位面积生物量为相应径级所有个体生物量之和与调查样地面积之比,样地单位面积生物量为所有个体总生物量与调查样地面积之比,径级内以及样地单位面积生物量的单位均换算为t·hm-2。计算生物量所使用的异速生长方程基本参数以及R²见表1。

图1 调查样地在亚热带地区的分布Fig.1 Distribution of investigated samples in subtropical area

图2 3种森林类型中的径级结构图Fig.2 Size-class distribution of individuals in three forest types
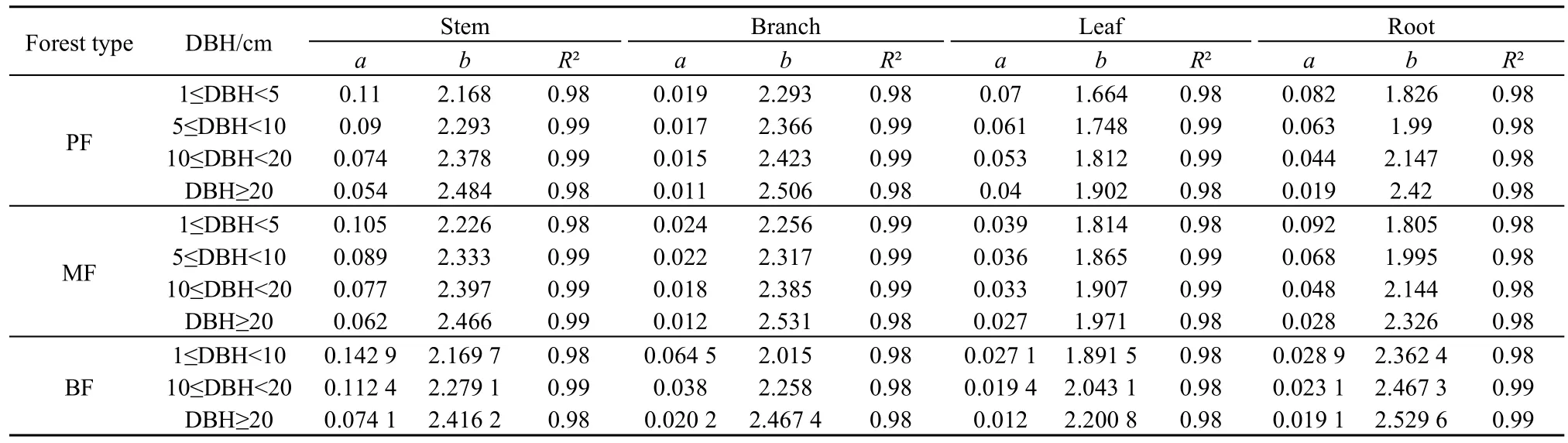
表1 计算调查样地生物量的基本参数以及R²Table1 Parameters and R-square used to calculate biomass in samples

其中,W为个体分器官的生物量(kg);l、b、s、r分别代表个体的叶、枝、干、粗根;a、b为拟合的固定参数,在不同森林类型以及径级中取值不同;DBH为个体胸径。
1.2.3物种丰富度的计算
由于本研究调查样地面积大小不同,采用Chao et al.(2009)提出的公式(式2、式3)估算每个调查样地以及各径级的理论物种数作为物种丰富度指数。
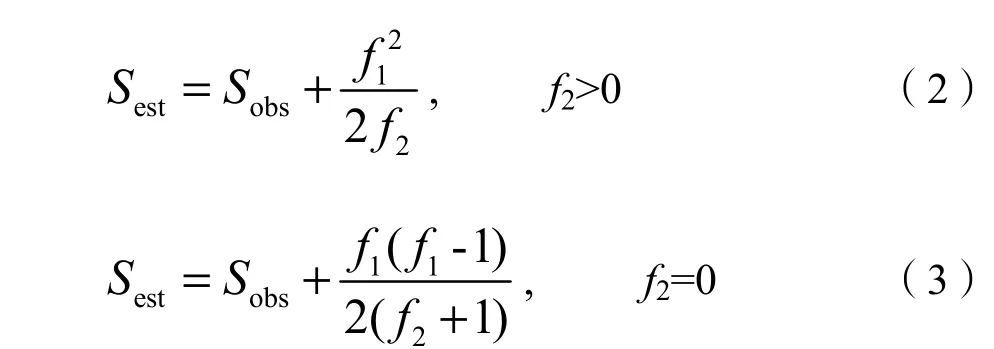
Sest为样地或径级内物种丰富度,Sobs为实际测量的样地或径级内物种数,f1为在样地或径级内个体数为1的物种个数,f2为样地或径级内个体数为2的物种个数。
研究表明物种丰富度与取样面积有关(马克平,1994)163,一些面积较小的样地其物种数可能不具有足够代表性;采用spearman秩相关分析,结果显示本研究样地以及各径级中物种数以及理论物种数与样地面积大小之间相关性未达到显著水平,表明样地面积不会影响群落物种丰富度,亦表明本研究选取样地面积大小已具有足够代表性。
2 结果与分析
2.1不同森林类型的生物量
整体而言,马尾松林(114.74±4.82)t·hm-2以及针阔叶混交林(124.99±4.99)t·hm-2的生物量均显著低于常绿阔叶林(220.9±8.62)t·hm-2,但前两种森林类型的生物量之间差异不显著(图3),表明由马尾松林逐渐恢复到常绿阔叶林的过程中,生物量的显著增加体现在针阔叶混交林向常绿阔叶林演替的阶段(图3)。

图3 3种森林样地总生物量以及物种丰富度的分布Fig.3 The distribution of total biomass and species richness in three forest types

图4 各径级生物量及分配Fig.4 Biomass and biomass allocation in each DBH class
比较各径级的生物量发现(图4),马尾松林样地中径级I~IV的生物量占样地总生物量的比例分别为(3.86%±1.06%)、(10.47%±1.29%)、(45.13%±2.35%)、(46.08%±2.90%),可见马尾松林的生物量集中在DBH≥10 cm的个体中;针阔叶混交林样地中径级I~IV的生物量占样地总生物量的比例分别为(4.53%±0.84%)、(14.51%±1.22%)、(39.35%±1.66%)、(47.55%±2.20%),与马尾松林生物量径级分布特征基本一致;常绿阔叶林样地中径级I~IV的生物量占样地总生物量的比例分别为(0.93%±0.2%)、(5.5%±0.73%)、(19.83%±1.79%)、(74.51%±2.39%),表明其生物量集中在少量大径级(DBH≥20 cm)的个体中。由此可见,随着马尾松林向常绿阔叶林的正向演替,生物量逐渐向大个体集中。
2.2不同森林类型的物种丰富度
马尾松林、针阔叶混交林以及常绿阔叶林的总物种丰富度分别为(5.81±0.61)、(17.55±1.17)、(23.71±2.04)(图4),整体而言针阔叶混交林与常绿阔叶林的物种丰富度水平相当,均显著高于马尾松林(图4)。
马尾松林中各径级的物种丰富度显著低于针阔叶混交林(图5);针阔叶混交林与常绿阔叶林的差异只在径级IV中达到显著水平,在其余径级中针阔叶混交林的物种丰富度略低于常绿阔叶林,但差异均未达到显著水平(图5)。因此,马尾松林在转变为常绿阔叶林的过程中物种多样性将显著提高,物种数量整体增长较快的过程可能出现在马尾松林向针阔叶混交林转变的阶段,在针阔叶混交林转变为常绿阔叶林的阶段,生长状况良好的大个体物种数量将有所上升,但群落整体的物种数量并未出现显著增加。

图5 各径级物种丰富度Fig.5 Species richness in each DBH class
2.3生物量以及物种丰富度之间的关系
总体而言,各森林类型中生物量均不随物种丰富度增加存在明显的变化趋势(图6),但生物量与物种丰富度的关系在不同的径级中表现不同,马尾松林、针阔叶混交林以及常绿阔叶林的小个体中(DBH<5 cm),生物量随物种丰富度的增加而显著增加;此外,针阔叶混交林中大个体(DBH≥5 cm)的生物量也随物种丰富度增加而显著增加。但是马尾松林以及常绿阔叶林大个体的生物量未随物种丰富度增加表现出显著的变化趋势。

图6 森林样地中物种丰富度与生物量的关系Fig.6 The relationship between species richness and biomass in forest samples
3 讨论
3.1不同森林碳汇功能以及物种多样性的变化
本研究结果表明从马尾松林演替至常绿阔叶林的过程中,生物量以及碳贮量显著提高。已有的研究认为森林植被层的生物量或碳贮量随演替进行逐渐增加,在达到演替顶级前趋于稳定(Law etal.,2003;Hudiburg et al.,2009)。大量研究已经证明常绿阔叶林作为亚热带地区的演替顶级群落而具有高生物量(林益明等,1996;温达志等,1997;杨同辉等,2010),这可能是常绿阔叶林对亚热带地区气候适应的结果(Kenzo et al.,2010;Ma et al.,2015358-360);马尾松林与针阔叶混交林的群落生物量相对较低,且二者之间的差异不显著(图3),表明两种森林的碳贮量相当,这与前人的研究结果一致(Zhang et al.,2007;Ma et al.,2015358-360),原因可能由于演替初期的马尾松作为速生树种很快地实现了固碳过程,使碳贮量达到较高水平(Chen et al.,2015),但在转变为针阔叶混交林的过程中,生长速率逐渐稳定,且林分密度有所下降(Peterson et al.,2014;Ma et al.,2015358-360),碳贮量未有显著上升。
本研究结果表明,马尾松林以及针阔叶混交林的生物量集中在DBH≥10 cm的个体,但常绿阔叶林的生物量主要集中在DBH≥20 cm的个体(图4)。已有的研究表明个体死亡率随径级的增大明显降低,小个体死亡率易受环境因素的影响(Wang et al.,2012)5,导致固定的碳因树木死亡而被归还至大气中的可能性较大。因此当马尾松林以及针阔叶混交林转变为常绿阔叶林时,生物量向大个体集中,碳库结构的稳定性以及森林应对气候变化的能力将有所提升。
本研究结果表明随着森林恢复演替的不断进行,物种多样性显著增加(图4)。物种数量的增加将提升生态系统的稳定性,因为功能相似的不同物种对环境的响应以及响应时间的不同可缓冲环境对生态系统的影响(Lehman et al.,2000;Chapin et al.,2002265-277;Loreau et al.,2013;Morin et al.,2014)。马尾松林的物种丰富度显著低于针阔叶混交林,可见其群落的稳定性远不及针阔叶混交林,这在已有的研究中已得到证实(张真等,2008)142-144;而针阔叶混交林与常绿阔叶林的物种丰富度差异只在径级IV(DBH≥20 cm)中达到显著水平(图5),且二者的总物种丰富度差异不显著(图4),表明针阔叶混交林的群落稳定性已经接近常绿阔叶林的水平。
3.2物种多样性与生态系统功能的关系
研究区域的森林生物量未随物种丰富度增加呈现显著的变化趋势(图6),表明整体群落的物种多样性与生态系统功能之间无明显的关系,这与一些基于控制实验的研究结果有所不同(Tilman,19991463-1465;Pretzsch,200541-61)。其原因可能有两个:环境因素对于生态系统功能的影响高于物种多样性(Hooper et al.,2005)19-21,且环境的异质性影响甚至掩盖物种多样性与生态系统功能的关系(Fridley,2002;Vila et al.,2005),因此本研究区域内环境异质性加强可能是削弱二者之间关系的一个重要原因;其次,物种添加或去除控制实验中的群落组成与自然森林生态系统均存在较大差异,例如草地中最有利的竞争者往往也具备很高的生产力(Rees et al.,2001),但森林中竞争力强的物种却未必生产力也较高(Szwagrzyk et al.,2007)555-556,在森林的控制实验中物种数量一般较少(Chen et al.,2003;Pretzsch,200541-61),与自然森林生态系统物种组成差异较大,所以结果未必适用于自然的森林生态系统。
本研究发现3种森林的小个体中(1 cm≤DBH<5 cm)的生物量均随物种丰富度的增加而显著增加(图6)。群落中小个体物种数量的增加提升了高生产力物种出现的可能性,从而提高了资源的利用效率,进一步提高了群落的生物量或生产力(Tilman,1999)1463-1464;这一现象也可以认为是由补偿效应引起的,小个体物种数量的增加提高了资源的利用效率,使原本流失的能量转变为生物量或生产力(Hooper et al.,2005)8-10。研究表明存储在小个体中的碳属于活性生物量碳库(Xiao et al.,2014),周转较快。这是因为小个体产生的含有高浓度氮以及低浓度木质素的凋落物能够被快速分解(Chapin et al.,2002151-173);其次由于环境的胁迫、竞争等因素的影响,小个体具备很高的死亡率,易形成粗死木质残体(唐旭利等,2003;Wang et al.,20125-8;Shen et al.,2013);所以小个体生物量的增加能够产生更多优质的凋落物以及粗死木质残体以促进森林养分循环(Yin,1999;Zhou et al.,2007)。本研究中针阔叶混交林的大个体(DBH≥5 cm)的物种丰富度与生物量之间也表现出较明显的线性关系,表明增加针阔叶混交林中大个体的物种数量同样也能够提升大个体的资源利用效率,从而增加大个体的生物量。因此,在演替的进行中对针阔叶混交林大个体进行合理的管理具有重要意义,但在此方面仍然需要有更多深入的研究。
4 结论
亚热带东部湿润地区的马尾松林向常绿阔叶林演替时生物量增加较明显,且演替的过程中群落生物量逐渐向大个体乔木集中,表明该区域位于演替顶级的森林群落拥有最大的碳库组成以及最稳定的碳库结构。马尾松林与针阔叶混交林的物种丰富度差异显著,但针阔叶混交林与常绿阔叶林的物种丰富度差异不显著,表明演替发展至针阔叶混交林时,物种多样性已经接近其演替顶级的状态。3种森林类型中小径级的个体生物量随物种丰富度增加变化趋势明显,因此增加小个体的物种丰富度不仅能够提升群落中资源的利用效率,而且能够促进森林生态系统的物质循环;针阔叶混交林大个体的生物量随物种丰富度的增加也显著增加。因此,对针阔叶混交林中大个体乔木进行合理的管理能够促进群落对于资源的有效利用。
参考文献:
BALVANERA P,PFISTERER A B,BUCHMANN N,et al.2006.Quantifying the evidence for biodiversity effects on ecosystem functioning and services [J].Ecology Letters,9(10):1146-1156.
BU W,ZANG R,DING Y.2014.Field observed relationships between biodiversity and ecosystem functioning during secondary succession in a tropical lowland rainforest [J].Acta Oecologica,55:1-7.
CARDINALE B J,WRIGHT J P,CADOTTE M W,et al.2007.Impacts of plant diversity on biomass production increase through time because of species complementarity [J].Proceedings of National Academy of Sciences,104(46):18123-18128.
CHAO A,COLWELL R K,LIN C W,et al.2009.Sufficient sampling for asymptotic minimum species richness estimators [J].Ecology,90(4):1125-1133.
CHAPIN III F S,MATSON P A,MOONEY H A.2002.Principles of Terrestrial Ecosystem Ecology [M].New York,Springer-Verlag New York.
CHEN H Y H,KLINKA K,MATHEY A H,et al.2003.Are mixed-species stands more productive than single-species stands:an empirical test of three forest types in British Columbia and Alberta [J].Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere,33(7):1227-1237.
CHEN Y Q,LIU Z F,RAO X Q,et al.2015.Carbon Storage and Allocation Pattern in Plant Biomass among Different Forest Plantation Stands in Guangdong,China [J].Forests,6(3):794-808.
FANG J Y,CHEN A P,PENG C H,et al.2001.Changes in forest biomass carbon storage in China between 1949 and 1998 [J].Science,292(5525):2320-2322.
FRIDLEY J D.2002.Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities [J].Oecologia,132(2):271-277.
HOOPER D U,CHAPIN F S,EWEL J J,et al.2005.Effects of biodiversity on ecosystem functioning:A consensus of current knowledge [J].Ecological Monographs,75(1):3-35.
HUDIBURG T,LAW B,TURNER D P,et al.2009.Carbon dynamics of Oregon and Northern California forests and potential land-based carbon storage [J].Ecological Applications,19(1):163-180.
IPCC.2003.Good Practice Guidance for Land Use,Land-Use Change and Forestry [M].Hayama,Japan:IPCC/IGES.
IPCC.2014.Introductiory Chapter In:Climate Change 2014:Mitigation of Climate Change.Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [M].Cambridge,United Kingdom and New York,NY,USA,Cambridge University Press:122.
KENZO T,ICHIE T,HATTORI D,et al.2010.Changes in above and below ground biomass in early successional tropical secondary forests after shifting cultivation in Sarawak,Malaysia [J].Forest Ecology and Management,260(5):875-882.
LAW B E,SUN O J,CAMPBELL J,et al.2003.Changes in carbon storage and fluxes in a chronosequence of ponderosa pine [J].Global Change Biology,9(4):510-524.
LEHMAN C L,TILMAN D.2000.Biodiversity,stability,and productivity in competitive communities [J].American Naturalist,156(5):534-552.
LOREAU M,DE MAZANCOURT C.2013.Biodiversity and ecosystem stability:a synthesis of underlying mechanisms [J].Ecology Letters,16(S1):106-115.
LOREAU M,NAEEM S,INCHAUSTI P,et al.2001.Ecology biodiversity and ecosystem functioning:Current knowledge and future challenges [J].Science,294(5543):804-808.
MA J,BU R C,LIU M,et al.2015.Ecosystem carbon storage distribution between plant and soil in different forest types in Northeastern China [J].Ecological Engineering,81:353-362.
MORIN X,FAHSE L,DE MAZANCOURT C,et al.2014.Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics [J].Ecology Letters,17(12):1526-1535.
PAN Y,BIRDSEY R A,FANG J,et al.2011.A large and persistent carbon sink in the world's forests [J].Science,333(6045):988-993.
PAQUETTE A,MESSIER C.2011.The effect of biodiversity on tree productivity:from temperate to boreal forests [J].Global Ecology and Biogeography,20(1):170-180.
PETERSON R L,LIANG J,BARRETT T M.2014.Modeling Population Dynamics and Woody Biomass in Alaska Coastal Forest [J].Forest Science,60(2):391-401.
PRETZSCH H.2005.Diversity and productivity in forests:Evidence from long-term experimental plots [M]//Forest Diversity and Function:Volume 176.Berlin Heidelberg:Springer:41-61.
REES M,CONDIT R,CRAWLEY M,et al.2001.Long-term studies of vegetation dynamics [J].Science,293(5530):650-655.
SHEN Y,SANTIAGO L S,MA L,et al.2013.Forest dynamics of a subtropical monsoon forest in Dinghushan,China:recruitment,mortality and the pace of community change [J].Journal of Tropical Ecology,29(2):131-145.
SZWAGRZYK J,GAZDA A.2007.Above-ground standing biomass and tree species diversity in natural stands of Central Europe [J].Journal of Vegetation Science,18(4):555-562.
TILMAN D.1999.The ecological consequences of changes in biodiversity:A search for general principles [J].Ecology,80(5):1455-1474.
VILA M,INCHAUSTI P,VAYREDA J et al.2005.Confounding factors in the observational productivity-diversity relationship in forests[M].Berlin,Springer-Verlag Berlin:65-86.
WANG X G,COMITA L S,HAO Z Q,et al.2012.Local-Scale Drivers of Tree Survival in a Temperate Forest [J].PloS ONE,7(2):e29469.
XIAO YIN,ZHOU GUOYI,ZHANG QIANMEI,et al.2014.Increasing active biomass carbon may lead to a breakdown of mature forest equilibrium [J].Scientific reports,4:3681.
XU B,GUO Z,PIAO S,et al.2010.Biomass carbon stocks in China’s forests between 2000 and 2050:A prediction based on forest biomass-age relationships [J].Science China Life Sciences,53(7):776-783.
YACHI S,LOREAU M.1999.Biodiversity and ecosystem productivity in a fluctuating environment:The insurance hypothesis [J].Proceedings of the National Academy of Sciences of the United States of America,96(4):1463-1468.
YIN X W.1999.The decay of forest woody debris:numerical modeling and implications based on some 300 data cases from North America [J].Oecologia,121(1):81-98.
ZHANG J,GE Y,CHANG J,et al.2007.Carbon storage by ecological service forests in Zhejiang Province,subtropical China [J].Forest Ecology and Management,245(1-3):64-75.
ZHAO M,ZHOU G S.2005.Estimation of biomass and net primary productivity of major planted forests in China based on forest inventory data [J].Forest Ecology and Management,207(3):295-313.
ZHOU G Y,GUAN L,WEI X,et al.2007.Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong,China [J].Plant Ecology,188(1):77-89.
ZHOU G Y,HOULTON B Z,WANG W,et al.2014.Substantial reorganization of China's tropical and subtropical forests:based on the permanent plots [J].Global Change Biology,20(1):240-250.
方炜,彭少麟.1995.鼎湖山马尾松群落演替过程物种变化之研究[J].热带亚热带植物学报,3(4):30-37.
林益明,林鹏,李振基,等.1996.武夷山甜槠群落的生物量和生产力[J].厦门大学学报:自然科学版,35(2):269-275.
罗应华,孙冬婧,林建勇,等.2013.马尾松人工林近自然化改造对植物自然更新及物种多样性的影响[J].生态学报,33(19):6154-6162.
马克平.1994.生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J].生物多样性,2(3):162-168.
莫江明,彭少麟,BROWN S,等.2004.鼎湖山马尾松林群落生物量生产对人为干扰的响应[J].生态学报,24(2):193-200.
彭少麟,王伯荪.1993.鼎湖山森林群落演替之研究(英文)[J].华南植物学报,试刊:34-42.
唐旭利,周国逸,周霞,等.2003.鼎湖山季风常绿阔叶林粗死木质残体的研究[J].植物生态学报,27(4):484-489.
王希华,宋永昌,王良衍.2001.马尾松林恢复为常绿阔叶林的研究[J].生态学杂志,20(1):30-32.
温达志,魏平,孔国辉,等.1997.鼎湖山锥栗+黄果厚壳桂+荷木群落生物量及其特征[J].生态学报,17(5):47-54.
杨同辉,宋坤,达良俊,等.2010.中国东部木荷-米槠林的生物量和地上净初级生产力[J].中国科学:生命科学,40(7):610-619.
张真,李典谟.2008.马尾松毛虫暴发机制分析[J].林业科学,44(1):140-150.
周国逸,温达志,唐旭利,等.2015.生态系统固碳观测与调查技术规范[M].北京:科学出版社:102-104.
Variation of Biomass and Species Richness in Subtropical Forest Based on Pinus massoniana Succession
GAO Yifei1,2,ZHANG Jing1,2,TANG Xuli1*,WANG Wantong3,YIN Guangcai4
1.South China Botanical Garden,Chinese Academy of Sciences,Guangzhou 510650,China; 2.University of Chinese Academy of Sciences,Beijing 100049,China; 3.College of Tourism,Henan Normal University,Xinxiang 453007,China; 4.School of Environmental Science and Engineering,Guangdong University of Technology,Guangzhou 510006,China
Abstract:More and more concentration are concentrated on the stable terrestrial carbon sinks with the increasing of atmospheric CO2concentration.Afforestation and reforestation are believed to be effective measures to increase the terrestrial ecosystem carbon sink.However,most afforestation stands are fast growing pure species.Accordingly,such forests are vulnerable to environmental stresses.Compared with afforestation stands,natural forests or climax forests are rich in species composition and therefore have strong resistance for stress.To understand the relationship between biodiversity and carbon density is important to evaluate ecosystem service comprehensively,and also important for forest management.In this study,Forests which represent the natural successional sequence in the eastern subtropical of China were selected in order to investigate the relation of biomass and species richness.We selected 316 forest plots including pine (Pinus massoniana) forests (PF),mixed coniferous and broadleaved forests (MF),and evergreen broadleaved forests (BF) from the database of the strategic pilot research program carried out by the Chinese Academy of Sciences during 2011─2015.Biomass and species richness of each plot was estimated,for a further comparison,trees were classified into 4 size groups according to diameter at breast height (DBH),including 1 cm ≤ DBH < 5 cm,5 cm ≤ DBH < 10 cm,10 cm ≤ DBH < 20 cm,and DBH > 20 cm.Multiple comparison was used to compare the difference of biomass or the difference of species richness among different successional stages,and simple linear regression was used to explore the relationship between biomass and species richness.The results showed that the mean value of total biomass in PF,MF and BF were (114.74±4.82),(124.99±4.99) and (220.9±8.62) t·hm-2,respectively,and the species richness was (5.81±0.61),(17.55±1.17),(23.71±2.04),respectively.Compared with PF,species richness of MF increased significantly (P=0.000),while no great difference existed in biomass.Compared with MF,biomass of BF increased significantly (P=0.000),but species richness showed no difference.Significantly positive relationships were observed between biomass and species richness in DBH ≥ 5 cm arbors in MF (P=0.009) and in DBH ≤ 5 cm arbors in PF (P=0.003),MF (P=0.001) and BF (P=0.000).Our research showed that the increasing of the species richness of small arbors in the eastern subtropical Chinese forests could not only enhance the resource availability of communities,but also promote the material circulation of forest ecosystem.Increasing involvement in large trees management may have significant impactions on the forest carbon sink and species diversity in MF.
Key words:Pinus massoniana Lamb.; evergreen broad leaved forest; species richness; biomass; subtropics
收稿日期:2015-09-18
*通信作者
作者简介:高一飞(1991年生),男,硕士研究生,研究方向为生态系统生态学。E-mail:gaoyifei2013@sina.com
基金项目:中国科学院战略性先导科技专项项目(XDA05050000);中国森林生态系统固碳现状、速率、机制和潜力(XDA05050200)
中图分类号:Q948
文献标志码:A
文章编号:1674-5906(2016)01-0022-08
DOI:10.16258/j.cnki.1674-5906.2016.01.004
引用格式:高一飞,张静,唐旭利,王万同,尹光彩.亚热带马尾松林恢复过程中物种丰富度及生物量变化[J].生态环境学报,2016,25(1):22-29.
猜你喜欢
今日农业(2020年19期)2020-12-14
绿色科技(2019年5期)2019-11-29
亚热带农业研究(2019年2期)2019-09-05
星星·诗歌原创(2017年5期)2017-08-13
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
中国科技信息(2015年2期)2015-11-16
中国果菜(2015年2期)2015-03-11
亚热带资源与环境学报(2015年4期)2015-01-22
