
植物细胞固定化条件对海南粗榧细胞生产三尖杉酯碱的影响
2016-07-26 01:20潘亚华蒋旋娴李永成
海南大学学报(自然科学版) 2016年2期
潘亚华,蒋旋娴,李永成
(海南大学 食品学院,海南 海口570228)
植物细胞固定化条件对海南粗榧细胞生产三尖杉酯碱的影响
潘亚华,蒋旋娴,李永成
(海南大学 食品学院,海南 海口570228)
摘要:为了研究海南粗榧细胞的固定化条件及其对细胞的生长和三尖杉酯碱合成的影响,本研究利用海藻酸钠凝胶包埋法对海南粗榧悬浮细胞进行固定化,并改变了其固定化条件,同时测定了生物量、PAL酶活性、凝胶球硬度、蔗糖含量和三尖杉酯碱产量等指标,结果显示:海南粗榧固定化细胞的生长周期为40 d,其中,在0~15 d其细胞生长停滞,在15~30 d其细胞生长缓慢,而30~40 d为缓慢衰亡期;海南粗榧细胞固定化培养的最佳产物提取时间为第15 d;包埋过程中CaCl2的最适质量浓度为0.023 g/mL;鲜重细胞的最适接种量为10%.由此得出结论: 海南粗榧细胞固定化培养抑制了细胞的生长,延长了细胞的生长周期,但缩短了细胞生产三尖杉酯碱的周期.
关键词:海南粗榧; 固定化; 海藻酸钠; 三尖杉酯碱
海南粗榧(CephalataxusmanniiHook.f.),为三尖杉科(Cephalotaxaceae),三尖杉属(Cephalotaxus)植物,主产于海南.海南粗榧的根、茎、叶和果实中含有抗癌功效的生物碱,其中三尖杉酯碱和高三尖杉酯碱已在临床被广泛应用[1].但是,由于受到气候、地理、季节等的影响,海南粗榧的生长较为缓慢,自然资源有限,因此,使用海南粗榧细胞培养技术是实现规模生产三尖杉酯碱和高三尖杉酯碱等产物的有效途径.目前,陈林[2]等建立了海南粗榧悬浮细胞培养体系,获得的产物产量为1.4mg/L;李永成[3-4]、龙晓娟[5]等通过诱导调节海南粗榧悬浮细胞来合成三尖杉酯碱等的研究也取得了一定的进展.然而,在海南粗榧细胞的悬浮培养中也存在一些问题,例如放大培养困难,细胞的生长速度慢以及在培养过程中不稳定等.
就培养方式而言,植物细胞固定化是一种有前途的培养方式[6].它是通过提高单位重量细胞的生产率来提高三尖杉碱类的产量[7].与悬浮培养相比,植物细胞固定化培养具有很多优点,如提高抗剪切能力、保持细胞活力、促进次级代谢产物的合成与分泌[8]、可重复使用并能提高生产效率、易于生产后处理等[9].如今,国内外研究者已建立了包括银杏、长春花、红豆杉等药用植物在内的50余种植物细胞的固定化培养系统.Gillet等[10]通过烟草组织固定化培养来生产东莨菪苷的研究发现,烟草细胞固定化培养比悬浮培养的产量更高.目前,还未见有关通过海南粗榧细胞固定化培养来生产三尖杉酯碱类生物碱的报道.为此,本研究以工业化生产三尖杉酯碱为目标,尝试了将海南粗榧细胞的固定化培养作为生产次生代谢物的新途径.本试验采用植物细胞固定化法来培养海南粗榧细胞,并主要探讨了海南粗榧细胞的固定化方法、条件以及其对细胞的生长与次级代谢产物生产周期的影响,旨在为进一步的扩大培养提供理论与技术基础.
1材料与方法
1.1植物材料将王成韬诱导[11]的、实验室继代培养的海南粗榧愈伤组织均匀分散在改良的MS液体培养基中,并按陈林的方法[2]进行培养来获得海南粗榧悬浮细胞,最终选用继代3次以上的海南粗榧悬浮细胞系来进行试验和研究.
1.2海南粗榧细胞固定化凝胶珠的制备取一定质量(鲜重)的海南粗榧细胞与质量浓度为20g·L-1的无菌海藻酸钠溶液混匀,用滴管吸取海藻酸钠-海南粗榧细胞混合液,并将其一滴一滴地均匀滴入无菌CaCl2溶液中,静置30~60min后,将其制成直径为4~5mm的均匀包埋有海南粗榧细胞的固定化凝胶珠.最后,用无菌新鲜液体培养基冲洗固定化凝胶珠2次,待用[12].
1.3海南粗榧细胞固定化凝胶珠的培养方法在无菌条件下,将洗涤干净的海南粗榧细胞固定化凝胶珠转入改良的MS培养基中,摇瓶振荡,并进行暗培养,培养温度为27 ℃,摇床转速为120r·min-1.
1.4产物提取海南粗榧固定化细胞胞外三尖杉酯碱的提取:将海南粗榧固定化细胞培养液抽滤,用氨水将滤液pH调至8.0,并用与滤液等体积的三氯甲烷萃取3次,接着,将三氯甲烷提取液于45 ℃下真空浓缩,直至完全蒸干,再用1mL色谱级甲醇复溶,在过孔径为0.22μm的滤膜后,待测.
海南粗榧细胞固定化凝胶球及细胞内三尖杉酯类碱的提取:将海南粗榧细胞固定化培养液抽滤,将包埋有海南粗榧细胞的海藻酸钙凝胶球混合石英砂,待研磨至糊状后加入三氯甲烷60mL,在超声30min后浸提12h,然后抽滤,并于45 ℃下将滤液浓缩,直至三氯甲烷完全蒸干,再用1mL色谱级甲醇复溶,在使用孔径为0.22μm的滤膜过滤后,待测.
采用高效液相色谱法分析三尖杉酯类碱量,测定条件与李永成设定的方法[3]一致.采用Agilent1200高效液相色谱仪,条件为:色谱柱为XDB-C18柱(50mm×4.6mm×10μm);流动相配比为V0.02mol·L-1乙醇铵∶V甲醇=55 ∶45;柱温为25 ℃;检测波长为288nm;进样量为10μL;流速为0.8mL·min-1.
1.5指标测定生长曲线的测定按万晓琦等的方法[13]进行.将海南粗榧细胞固定化培养后,每隔5天取大小相同的固定化细胞凝胶球15粒,将凝胶球放入0.1mol·L-1,pH=7.8的磷酸缓冲液中振荡,待凝胶球破碎溶解后过滤,过滤后的残渣即为海南粗榧的鲜重细胞,并依据细胞鲜重的测定结果绘制海南粗榧的生长曲线.
培养基含糖量的测定按李永成所采用的苯酚硫酸法[14]进行.
苯丙氨酸解氨酶(PAL酶)活性的测定[3].将海南粗榧细胞固定化凝胶球与50mmol·L-1pH8.8的Tris-HCl缓冲液(包含15mmol·L-1的β-硫基乙醇)均匀混合,冰浴研磨,将研磨液在10 000r·min-1和4 ℃下离心30min,收集上清液得PAL酶粗提液.将10mmol·L-1的L-苯丙氨酸、0.4mL的蒸馏水、PAL酶粗提液混合,在30 ℃下反应30min[15],然后添加0.1mL6mol·L-1的HCl来终止反应.在290nm波长处测定反应产物吸光度,并以每小时OD值在290nm处变化0.01为一个单位.
培养基游离Ca2+浓度的测定:使用Sigma钙离子检测试剂盒(CalciumColorimetricAssay),并参照说明书中的测定方法进行.分别将10μL样品和10μL蒸馏水(空白调零组)加入到含有96孔板的管槽中,加入蒸馏水至50μL,滴加90μL显色剂和60μL的测定缓冲液到管槽中,混合均匀.在室温避光条件下使其反应5~10min,然后将反应后的混合液在波长575nm处测定吸光度,将吸光度值代入钙离子标准曲线中的到培养基中钙离子浓度.
凝胶球硬度的测定参考王秀娟[16]改进后的方法.使用CT3型质构仪对海藻酸钙凝胶球进行TRA质构分析,测定条件:探头为TA-39(直径为2mm)平底柱型探头;测试速度为0.5mm·s-1;触发点负载为5g;行变量30%.
1.6数据处理本试验数据为3个平行实验的平均数,数据以±s表示,同时利用Origin8.0和SigmaPlot12.5进行绘图,P<0.05视为差异显著.
2结果与分析
2.1粗榧细胞固定化培养过程中的细胞生长曲线图1和图2分别为海南粗榧悬浮细胞固定化培养过程中的生物量变化及其培养基蔗糖的消耗变化,其接种量为10%.从图1可见,海南粗榧细胞固定化培养周期约为40d:初期其细胞生长停滞;15d后固定化细胞进入生长对数期,生物量增加,细胞缓慢生长;至30d左右生物量不再增加,细胞褐化严重.从图2可见,在海南粗榧细胞固定化培养的初期,培养基糖耗很慢,说明海南粗榧固定化细胞刚进入调整期,并逐渐适应固定化环境,糖代谢缓慢,细胞生长停滞;在培养的第25~30 天,糖耗速度加快,表明此时固定化细胞缓慢生长;固定化培养的末期,由于培养基中蔗糖和其他营养物质的减少而导致细胞开始减少,糖耗速度变缓.但是,从培养40d的海南粗榧固定化细胞中提取培养物后并没有发现三尖杉酯碱.由此推测,海南粗榧固定化细胞的生长周期与次级代谢产物的生产周期不一致,因此,有必要研究海南粗榧固定化细胞合成三尖杉酯碱的代谢周期.

2.2海南粗榧细胞固定化培养的最佳生产周期及其PAL酶活性的变化图3为固定化海南粗榧细胞合成三尖杉酯碱周期的研究结果.由图3可见,随着固定化海南粗榧细胞的培养时间增加所获得的三尖杉酯碱的量先增加后减少,在培养的第15d获得最大产量(2.018mg/L).因此,粗榧细胞固定化培养的产物最佳收获时间是第15d.图4为海南粗榧细胞固定化培养过程中的苯丙氨酸解氨酶(PAL酶)的活性变化.从图4可见,海南粗榧细胞固定化培养前期的PAL酶活性相对较高,但随着细胞的继续培养,其PAL酶活性下降,且在培养后期其活性处于该水平.

2.3包埋过程中CaCl2浓度对海南粗榧固定化细胞生产三尖杉酯碱的影响图5和图6分别为固定化包埋过程中不同浓度的CaCl2溶液对培养基中游离Ca2+浓度以及凝胶球硬度的影响.由图5可见,在海藻酸钠-海南粗榧细胞混合液中滴入不同浓度的CaCl2溶液后,所形成的凝胶球被充分洗涤2次后,其对培养基中游离钙离子质量浓度的影响不显著;当CaCl2质量浓度>0.031g·mL-1时,海南粗榧固定化细胞培养5d后全部褐化死亡;当CaCl2质量浓度在0.015~0.031g·mL-1时,如图6所示,凝胶球硬度随CaCl2质量浓度的升高会不断增强.取不同质量浓度的CaCl2溶液进行固定化包埋后,所制成的海南粗榧固定化细胞凝胶球经培养后生产三尖杉酯碱的情况如图7所示,当CaCl2质量浓度为0.023g·mL-1时,海南粗榧固定化细胞所生产的三尖杉酯碱的产量最高.


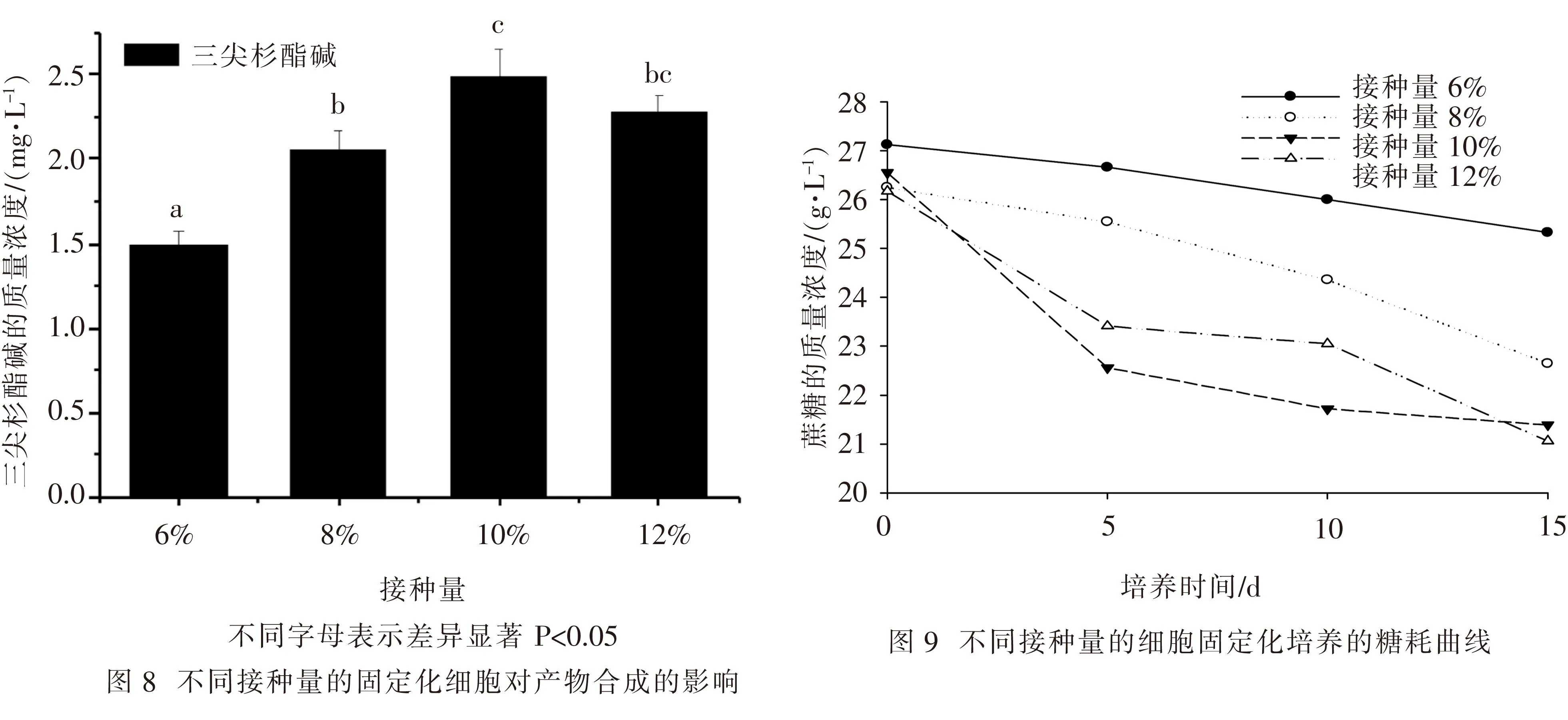
2.4细胞接种量对海南粗榧固定化细胞生长和其生产三尖杉酯碱的影响 由图8表明,在海南粗榧细胞固定化培养的过程中,随着接种量的增加三尖杉酯碱的产量先增加后减少,接种量为10%时可获得最大产物产量;接种量为12%时比接种量为8%时所获得的产物产量要高.因此,利用海南粗榧固定化细胞生产三尖杉酯碱的最佳接种量为10%.图9为不同接种量下培养基蔗糖含量的测定结果,图9表明,随着接种量的增加,细胞耗糖速率也随之增加,但是,当接种量超过10%时糖耗速度反而减缓.
3讨论
植物细胞固定化培养使细胞生长期延长,它使细胞在较长的时间内保持了较高的活力[17].当接种量为6%时进行粗榧细胞的悬浮培养,此时可发现,悬浮细胞生长的0~10d为生长滞后期,培养的第10~25 天为对数生长期,第25~30d为稳定期[2].与之相比,本试验中所采用的固定化培养法使海南粗榧细胞的生长期延长了.细胞被固定化后,由于其周围的生长环境较悬浮培养环境有了较大的不同,故细胞的生长周期以及代谢周期都发生了一定变化[18].采用植物细胞固定化法时,在包埋及培养过程中虽然细胞直接与固定剂接触会对细胞产生一定的毒性作用,不利于细胞的生长,但是,这对于次生代谢物的合成是有利的[19].PAL酶存在于所有绿色植物中,它是次级代谢途径的起始酶,也是初生代谢和次生代谢的纽带.诱导植物细胞的防御反应虽能够促进细胞次级代谢产物的合成,但同时也会伴随细胞的防御反应,这同样会引起PAL酶活性的变化[3].本研究的结果表明,在海南粗榧细胞固定化培养的过程中细胞的生长周期(40d)与次级代谢产物的生产周期不一致,其合成三尖杉酯碱的总量在固定化法培养15d左右达到最高(2.018mg·L-1),与之相比,陈林[2]等要在海南粗榧细胞悬浮培养的第25 天才能获得产物的最大产量(1.4mg·L-1);细胞固定化培养的第15 天以后,三尖杉酯碱量不断减少,至海南粗榧固定化细胞生长的末期已检测不到三尖杉酯碱.可见,海南粗榧细胞固定化培养不仅提高了产物的产量,而且还缩短了生产周期,提高了生产效率.在海南粗榧细胞固定化培养的过程中,其PAL酶的活性由高降低,这可能是由于在海南粗榧细胞固定化培养的初期,海南粗榧细胞受到固定化条件的胁迫作用而引起细胞的防御反应,从而致使细胞生长缓慢并导致了其PAL酶的活性较高,但同时也促进了细胞合成三尖杉酯碱.随后,由于细胞逐渐适应了固定化培养环境而进入对数生长期,此时由于PAL酶的活性有所降低,因而细胞合成三尖杉酯碱的量逐渐减少.但是,在海南粗榧固定化细胞生长的末期,为何细胞内外三尖杉酯碱的含量会不断减少,目前对这一现象的代谢机制尚不清楚.
在植物细胞的代谢过程中,钙离子作为高等植物细胞内普遍存在的一种信使分子,它参与并调节了植物细胞的多种生理反应[20].但是,过多的Ca2+也会对植物细胞产生毒性刺激,从而对细胞的生长代谢产生影响;此外,在固定化过程中由于氯化钙质量浓度的不同也会导致海藻酸钙凝胶球硬度的不同[7],从而造成细胞的机械胁迫情况不同.本试验研究了海南粗榧细胞固定化包埋过程中不同CaCl2质量浓度对固定化细胞代谢的影响,结果表明,海南粗榧细胞固定化所形成的凝胶球被充分洗涤后不会影响培养基中的游离Ca2+浓度,从而排除了培养基中的游离Ca2+质量浓度的变化对细胞代谢的诱导作用.因此,用不同质量浓度的CaCl2溶液作固定剂所造成的海藻酸钙凝胶球硬度的不同是影响固定化细胞代谢的主要因素,由结果可见,当凝胶球达到一定的硬度时会促进细胞合成三尖杉酯碱,但当凝胶球硬度过高时细胞合成三尖杉酯碱的量则会降低.
由于在植物细胞固定化培养中可通过提高单位重量细胞的生产率来提高产量[9],因此,在海南粗榧细胞固定化培养的工艺条件中也可通过提高海南粗榧细胞的接种量来提高三尖杉酯碱的产量,这也是一种较直接的途径,但是由于营养物质以及氧气的限制,过高的接种量并不会再提高产物的产量,相反,由于细胞生长代谢受到抑制后反而会降低产物产量.本试验的结果表明,当接种量达到10%时,海南粗榧固定化细胞消耗基质中蔗糖的速率以及合成三尖杉酯碱的总量均已达到最大.
参考文献:
[1] 符文英,杜道林,邢诒旺.海南粗榧保护和开发利用的研究[J].分子植物育种,2003,1(5/6):795-799.
[2] 陈林,李永成.海南粗榧细胞悬浮培养体系的建立[J].广东农业科学,2014(24):54-62.
[3]LiYC.Effectsandactionmechanismsofsodiumfluoride(NaF)onthegrowthandcephalotaxineproductionofCephalotaxusmanniisuspensioncells[J].EnzymeandMicrobialTechnology,2014,67:77-81.
[4]LiYC.EnhancedcephalotaxineproductioninCephalotaxusmanniisuspensionculturesbycombiningglycometabolicregulationandelicitation[J].ProcessBiochemistry,2014,49:2279-2284.
[5] 龙晓娟,李永成.诱导子对海南粗榧悬浮细胞生长及三尖杉酯类碱合成的影响[J].热带作物学报,2015,36(6):1125-1130.
[6]BrodeliusP,DeusB,MosbachK,etal.Immobilizedplantcellsfortheproductionandtransformationofnaturalproducts[J].FEBSLett,1979,103(1):93-97.
[7] 余龙江,李为,刘幸福,等.中国红豆杉悬浮细胞固定化培养生产紫杉醇[J].华中理工大学学报,1998,26(5):1-4.
[8]ShibasakiKN,IizukaY,YonemotoT.CulturesofNicotianatabacumcellsimmobilizedincalciumalginategelbeadscoatedwithcell-freegelfilm[J].JChemEngJapan,2001,34(11):1431-1438.
[9] 王素芳.植物细胞的固定化培养[J].浙江万里学院学报,2004,17(2):110-114.
[10]GilletF,RoisinC.ImmobilizationofNicotianatabacumplantcellsuspensionswithincalciumalginategelbeadsfortheproductionofenhancedamountsofscopolin[J].EnzymeandMicrobialTechnology,2000,26(2/4):229-234.
[11] 王成韬,钟秋平,李永成.海南粗榧外植体灭菌方法研究[J].热带作物学报,2014,35(1):110-114.
[12] 田中,渥夫.植物细胞固定化的生产技术[J].细胞生物学杂志,1998,10(3):115-121.
[13] 万晓琦,宋萍,封磊,等.雷公藤细胞的固定化研究[J].福建林学院学报,2013,33(3):207-212.
[14] 李永成.东北红豆杉悬浮细胞与内生真菌在紫杉醇合成中相互转化关系的研究[D]. 无锡: 江南大学生物工程学院,2009.
[15]YuanYJ,LiC,HuZD,etal.AdoubleoxidativeburstfortaxolproductioninsuspensionculturesofTaxuschinensisvar.maireiinducedbyoligosaccharidefromFusariumoxysprum[J].EnzymeMicrobiolTechnol,2002,30:774-778.
[16] 王秀娟,张坤生,任云霞,等.海藻酸钠凝胶特性的研究[J].食品工业科技,2008,29(2):259-262.
[17]RamakrishnaSV.ProductionofsolavetivonebyimmobilizedcellsofHyoscyamusmuticus[J].BiotechnologyLetters,1993,15(3):301-306.
[18] 袁丽红.细胞固定化条件对紫草细胞生产紫草色素的影响[J].南京化工大学学报,1993,21(2):44-48.
[19]MithferA,SchulzeB,BolandW.Bioticandheavymetalstressresponseinplants:evidenceforcommonsignals[J].FEBSLett,2004,566:1-5.
[20] 易籽林.钙信号系统与植物激素信号的研究进展[J].中国农学通报,2010,26(15):221-226.
收稿日期:2015-11-26
基金项目:国家自然科学基金(NO.21166007)
作者简介:潘亚华(1990-),男,河北景县人,海南大学食品学院2013级硕士研究生,E-mail:1203737023@qq.com 通信作者: 李永成(1964-),男,苗族,海南海口人,博士,副教授,研究方向:生化工程,E-mail: lyc2360@sina.com
文章编号:1004-1729(2016)02-0155-07
中图分类号:Q 831.1
文献标志码:ADOl:10.15886/j.cnki.hdxbzkb.2016.0024
Effects of the Conditions for Immobilized Cell of Plant on theHarringonineProductionofCephalotaxus manniiImmobilizedCells
Pan Yahua, Jiang Xuanxian, Li Yongcheng
(CollegeofFoodScience,HainanUniversity,Haikou570228,China)
Abstract:In the study, to study the conditions for immobilizing cell of Cephalotaxus mannii and its effects on the growth and harringtonine synthesis of C. manni, the sodium alginate embedding method was performed to immobilize C. mannii, the immobilized conditions were changed, and biomass, the activity of phenylalanine ammonium-lyse, the hardness of gel beads, sucrose content, and the production of harringtonine were determined. The results indicated that the growth cycle of the immobilized cell of C. mannii was 40d, the lag period of growth was during 0-15d, the slow growing time was during 15-30th days, and the stable phase was during 30-40d; the culture conditions of the immobilized cell of C.mannii was optimized as followed: the optimal time point to extract the product of C. mannii was the 15th day; the optimal concentration of CaCl2 was 0.023 g.mL-1; to obtain the maximum harringtonine production, the optimal fresh weight of C. mannii was 10%. The culture conditions for immobilizing cell of C. mannii inhibited the growth and extended the growth time of C. mannii cell, but this method shortened processing periods of the harringtonine.
Keywords:Cephalotaxus mannii; immobilized; sodium alginate; harringtonine
