
烟草抗番茄斑萎病毒基因筛选与生物信息学分析
2016-09-26 06:57杨金广樊韦华杨柳青王耀峰宋玉川王凤龙刘晓璐
中国烟草科学 2016年4期
杨金广,樊韦华,杨柳青,丁 涛,王耀峰,宋玉川,董 昕,王凤龙,刘晓璐*
(1.中国农业科学院烟草研究所,烟草行业烟草病虫害监测与综合治理重点实验室,青岛 266101;2.北京科技大学化学与生物工程学院,北京100083;3.甘肃省烟草公司庆阳市公司,甘肃 西峰745000;4.云南烟草保山香料烟有限责任公司,云南 保山 678000)
烟草抗番茄斑萎病毒基因筛选与生物信息学分析
杨金广1,樊韦华2,杨柳青2,丁 涛2,王耀峰3,宋玉川4,董 昕2,王凤龙1,刘晓璐2*
(1.中国农业科学院烟草研究所,烟草行业烟草病虫害监测与综合治理重点实验室,青岛 266101;2.北京科技大学化学与生物工程学院,北京100083;3.甘肃省烟草公司庆阳市公司,甘肃 西峰745000;4.云南烟草保山香料烟有限责任公司,云南 保山 678000)
通过同源克隆的方法在TSWV高感烟草品种NC89、TSWV耐病烟草品种中烟100中分别克隆出一个疑似抗番茄斑萎病毒(Tomato spotted wilt virus, TSWV)的基因,分别命名为NtSw-5NC89和NtSw-5zy100。序列分析表明,NtSw-5NC89长3819bp,包含1个完整的读码框,编码1273个氨基酸,理论等电点和相对分子质量分别为5.64和147529.5Da;NtSw-5zy100长3165bp,包含1个完整的读码框,编码1054个氨基酸,理论等电点和相对分子质量分别为6.75和122032.9Da;NtSw-5NC89和NtSw-5zy100均不含信号肽,均无明显跨膜区,二者均含有大多数植物抗性蛋白所共有的CC-(NB-ARC)-LRR典型结构。系统进化树分析表明,NtSw-5NC89与马铃薯假定晚疫病抗性蛋白同源R1A-3近源关系最近,NtSw-5zy100与烟草假定晚疫病抗性蛋白同源R1A-3同型X2近源关系最近。该结果可为烟草番茄斑萎病毒抗性研究和烟草番茄斑萎病毒的防治提供理论基础。
烟草;番茄斑萎病毒;抗性基因;生物信息学分析
近年来,随着农业产业结构的调整以及全球农副产品贸易的频繁,造成众多植物病毒及其媒介昆虫的全球传播和蔓延。番茄斑萎病毒(Tomato spotted wilt virus,TSWV)正是在这种情况下,随其虫媒介体蓟马(Thrip)进入我国,在我国烟草种植区猖獗危害[1]。种植抗病品种是防治番茄斑萎病最有效的防治措施。当前对TSWV抗性的研究主要集中在番茄和辣椒上,对烟草TSWV的抗性研究较少[2]。已明确有两个抗病基因Sw-5和Sw-7对TSWV具有明显抗性,通过转基因技术或基因渗透(杂交)获得的含Sw-5的植株表现出较强的抗性,但Sw-5 对TSWV6、Anwa-1、DaWA-1d和TOTAS-1d等4个株系的TSWV不表现抗性[3-5]。Sw-7可帮助番茄有效抵御TSWV6和Anwa-1株系的侵染[6]。TSWV在农作物上的暴发流行,除了病毒本身传播介体繁多外,另一个重要的因素是TSWV可以通过基因突变和重组迅速适应并打破寄主抗性。对TSWV基因组变异特征分析表明TSWV的NSs和NP两个蛋白突变导致寄主Tsw基因抗性丧失[7]。国内外对番茄斑萎病毒的抗性品种进行了大量研究,目前可通过小孢子原生质体融合技术,将野生烟草的抗性基因导入栽培品种使其获得抗性,但迄今尚未培育出对番茄斑萎病毒抗性较强的烤烟品种[8]。由于对TSWV的自然抗病基因非常少,筛选工作困难,目前对烟草番茄斑萎病抗性基因的研究比较缓慢,科学家正致力于基因工程手段,将筛选得到的抗性基因经优化表达后导入易染病的栽培烟草基因组中,培育抗病品种。
本研究根据番茄中已确定为抗 TSWV基因序列,于烟草全基因组数据库中,通过生物信息学的方法,筛选烟草基因组中疑似抗TSWV的基因,通过同源克隆的方法在 TSWV高感品种 NC89和TSWV耐病品种中烟100中克隆疑似靶基因。对所获基因进行测序、蛋白结构和功能域预测、蛋白进化分析,推测该基因编码蛋白在细胞中的存在位置、结构和功能以及与其他蛋白的近源关系,为烟草番茄斑萎病毒抗性研究和烟草番茄斑萎病毒的防治提供理论基础。
1 材料与方法
1.1材料
选用烟草品种NC89、中烟100,经田间试验检测NC89为TSWV高感品种,中烟100为TSWV耐病品种。
1.2生物信息学分析及引物设计
于烟草基因组数据库 http://www.pngg.org/ tgi/press_release.html中,筛选烟草基因组中与番茄Sw-5基因同源性高的基因,经由序列比对,设计8对引物(表1),引物由上海生物工程技术公司合成。

表1 8对引物名称及序列Table 1 Names and sequences of the eight pair of primers
1.3种植烟草提取RNA,反转录为cDNA
将烟草NC89、中烟100种植于温室中,保持水分以确保种子发芽率。烟草生长繁盛时期,取叶片液氮速冻,用TIANGEN RNAprep pure Plant Kit提取RNA,invitrogenSuperScriptⅢ First-Strand反转录为cDNA。
1.4PCR扩增
以NC89、中烟100的cDNA为模板,表1中的引物,用phusionmix和KOD进行扩增。PCR扩增体系:取 cDNA l µL,10×buffer 5µL,10 mmoL/LdNTPs 4 µL,20 µmol/L上下游引物各l µL,TaqDNA聚合酶0.5 uL,加ddH2O至50 µL后进行扩增。PCR循环:94℃预变性5 min;94 ℃ 30 s,57 ℃ 30 s,72 ℃ 3min 30 s,35次循环;最后72 ℃延伸l0 min。PCR产物用0.8%琼脂糖凝胶电泳检测。用Promega Wizard SV Gel and PCR Cleanup System进行胶回收纯化。
1.5克隆与测序
PCR产物纯化后克隆到载体pEASY上,并转化大肠杆菌Trans-T1感受态细胞。培养后,进行涂板(培养基含氨苄青霉素)检测,每对引物对应的培养物各挑8个单克隆用rTaq进行菌落PCR阳性检测。将阳性结果送上海生物工程技术公司进行双向测序。
1.6蛋白功能域预测
利用 Expasy-ProtParam进行理化性质分析,ExPASy-ProtScale进行疏水性结构分析,用SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP/)预测蛋白信号肽,用在线 TMHMM Serverv.2.0(http: //www.cbs.dtu.dk/services/TMHMM/)预测蛋白序列跨膜区,用 NCBI-Blast进行结构域预测分析,用Predictprotein进行作用位点分析。
1.7蛋白进化分析
应用NCBI-Blast和MEGA6等软件对所得序列进行分析。
2 结 果
2.1RNA提取
紫外分光光度计测定提取的叶片总 RNA,OD260/OD280在1.9~2.0,电泳检测条带清晰无拖带,所得RNA质量很好,基本无降解。
2.2利用8对全长引物获得的烟草抗TSWV基因的克隆
表1中第2和第7对引物对NC89 cDNA各扩增出一段4000 bp左右的片段,第3和第4对引物对中烟100 cDNA各扩增出一段4000 bp左右的片段。但菌落PCR阳性检测中只有第2和第3对引物对应的单克隆培养物检测出4000 bp左右的条带。
2.3筛选所得核酸序列和推导出的氨基酸序列
获得了烟草品种NC89的目的片段,测序结果表明,该片段全长3819 bp,包含1个完整的读码框,编码1273个氨基酸(图1)。经NCBI序列比对,该片段与Sw5-a、Sw5-b、Sw5-c、Sw5-d、Sw5-e的相似度分别为78%、78%、77%、77%、77%,相似度较高,命名此段序列为NtSw-5NC89。
获得了烟草品种中烟100的目的片段,测序结果表明,该片段全长3165 bp,包含1个完整的读码框,编码1054个氨基酸(图2)。经NCBI序列比对,该片段与Sw5-a、Sw5-b、Sw5-c、Sw5-d、Sw5-e的相似度分别为79%、79%、78%、79%、77%,相似度较高,命名此段序列为NtSw-5zy100。
2.4筛选所得基因编码蛋白质的结构分析
理化性质分析结果显示,NtSw-5NC89的理论等电点和相对分子质量分别为5.64和147529.5Da,总平均疏水指数为-0.201。NtSw-5zy100的理论等电点和相对分子质量分别为6.75和122032.9 Da,总平均疏水指数为-0.141。疏水性分析结果(图3A、B)显示,NtSw-5NC89和NtSw-5zy100的疏水性峰值高于0的氨基酸占总氨基酸数目的比例均低于 50%,且NtSw-5NC89和NtSw-5zy100的总平均疏水指数均小于0,可以断定二者均为亲水性蛋白。蛋白跨膜区域分析结果(图3C、D)显示,二者均无明显跨膜区。NtSw-5NC89和NtSw-5zy100信号肽预测结果(图3E、F)显示,二者均无信号肽。Predictprotein二级结构分析,预测NtSw-5NC89的α-螺旋、β折叠、卷曲结构所占比例分别为56.09%、6.36%、37.55%;预测NtSw-5zy100的α-螺旋、β折叠、卷曲结构所占比例分别为、β折叠、卷曲结构所占比例分别为51.71%、7.87%、37.55%。蛋白质结合位点预测结果(图4A、C)显示,NtSw-5NC89和NtSw-5zy100均含有一个DNA结合位点(DNA-binding-region)和多个可能的蛋白结合位点(protein binding region)。NCBI-Blast结构域预测(图4B、D)显示NtSw-5NC89和NtSw-5zy100均含有抗性蛋白卷曲螺旋(RX-CC_like)和P-loop核苷三磷酸水解酶(P-loop_NT)结构域以及核苷酸结合-臂重复蛋白(NB-ARC)结构域、亮氨酸重复(LRR)结构域,即CC-(NB-ARC)-LRR结构,NtSw-5zy100因LRR结构分布较为分散未能在图中集中体现。

图1 NtSw-5NC89编码区核酸序列及其推导出的氨基酸序列(阴影部分为功能结构域)Fig. 1 Nueleotide and predicted amino acid sequence of NtSw-5NC89(the shaded part for function structure domain)
2.5系统进化树分析通过NCBI-Blastp在线同源性比对,应用MEGA6软件进行系统进化树分析,结果显示(图5)NtSw-5NC89和 NtSw-5zy100分别位于一个大分支的两个不同小分支上,其中NtSw-5NC89与马铃薯假定晚疫病抗性蛋白同源R1A-3(NCBI蛋白序列号XP_ 015168988.1)亲源关系最近(100%氨基酸相似度)。NtSw-5zy100与烟草假定晚疫病抗性蛋白同源R1A-3同型X2(NCBI蛋白序列号XP_ 009777376.1)近源关系最近(84%氨基酸相似度)。所获得的两个基因编码蛋白同番茄斑萎病抗性蛋白家族(抗性蛋白A、抗性蛋白B、抗性蛋白C、抗性蛋白D、抗性蛋白E,分别由Sw5-a、Sw5-b、Sw5-c、Sw5-d、Sw5-e编码,NCBI蛋白序列号分别为AAG31013.1、AAG31014.1、 AAG31015.1、 AAG31016.1、AAG31017.1)不在一个大分支上,同源性相对较低(NtSw-5NC89与抗性蛋白A、抗性蛋白B、抗性蛋白C、抗性蛋白D、抗性蛋白E氨基酸相似度分别为58%、58%、61%、61%、61%;NtSw-5zy100与抗性蛋白A、抗性蛋白B、抗性蛋白C、抗性蛋白D、抗性蛋白E氨基酸相似度分别为64%、65%、63%、60%、61%)。

图2 NtSw-5zy100编码区核酸序列及其推导出的氨基酸序列(阴影部分为功能结构域)Fig. 2 Nueleotide and predicted amino acid sequence of NtSw-5zy100(the shaded part for function structure domain)
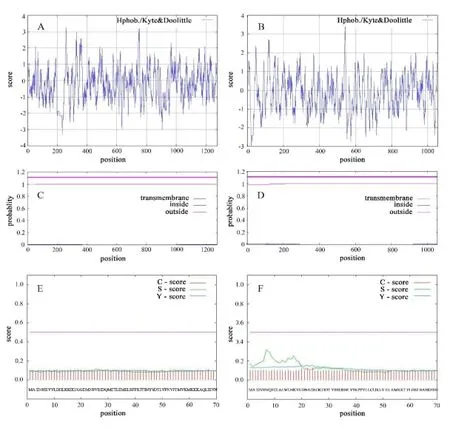
图3 NtSw-5NC89和NtSw-5zy100的疏水性分析、跨膜区域预测和信号肽预测Fig. 3 Hydrophobicity analysis, transmembrane region prediction, and signal peptide prediction of NtSw-5NC89and NtSw-5zy100
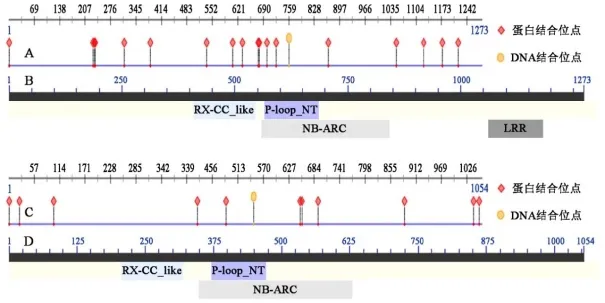
图4 NtSw-5NC89和NtSw-5zy100结合位点和结构域分析Fig. 4 Binding site and structure analysis of NtSw-5NC89and NtSw-5zy100

图5 基于NtSw-5NC89和NtSw-5zy100的系统进化树分析Fig. 5 Phylogenetic tree analysis of NtSw-5NC89and NtSw-5zy100
3 讨 论
本文通过同源克隆方法在TSWV高感品种NC89和TSWV耐病品种中烟100中分别筛选出疑似抗TSWV的基因NtSw-5NC89和NtSw-5zy100。运用生物信息学的方法分析了该基因的理化性质,并对其功能进行了预测。分析表明,NtSw-5NC89和NtSw-5zy100编码蛋白均不含信号肽、均无明显跨膜区,这与Sw5-a和Sw5-b编码蛋白结构相似[9],推测此二者编码的蛋白不是分泌性蛋白,该类蛋白不可能定位于膜上。系统进化树分析表明,所得两个基因编码蛋白与假定晚疫病抗性蛋白同源型近源关系相近,而与番茄斑萎病抗性蛋白家族近源关系相对较远,可能与基因所在物种的差异性有关,不同物种对同一植物病毒的抗性蛋白可能有各自的特异性。
对 NtSw-5NC89和 NtSw-5zy100结构域分析可看出,二者均含有RX-CC_like、P-loop_NT、NB-ARC、LRR等结构,这与Sw5-a、Sw5-b编码的抗性蛋白A、抗性蛋白B极为相似(抗性蛋白A、抗性蛋白B中均含有CC-(NB-ARC)-LRR结构)[9]。RX-CC结构是马铃薯病毒X抗性蛋白(RX)或其相似蛋白的N端卷曲螺旋结构,此结构能够与抗病性反应所必须的RanGAP2辅助因子相作用,进而产生抗病效应。P-loop结构结构含有NTP水解酶结构域,包括 GTP酶激酶(KG)类和额外链催化酶(Additional Strand Catalytic Enzyme, ASCE)类两个主要的结构类,参与多种细胞内反应,在番茄斑萎病抗性蛋白A、抗性蛋白B中均能找到类似结构的激酶结构域[9],该结构域可能在病毒抗性反应中具有重要作用。LRR结构易于形成亮氨酸拉链,参与蛋白质的多聚化和与其他蛋白的结合,番茄斑萎病抗性蛋白A只有一个亮氨酸拉链,而番茄斑萎病抗性蛋白B有两个,通过烟草转化试验发现,Sw5-b对TSWV有抗性,而Sw5-a无,这可能与其结构中的亮氨酸拉链个数有关[9]。NtSw-5NC89的LRR结构连续分布,而NtSw-5zy100分布却较为分散,因此在形成亮氨酸拉链时产生差异,这有可能是 NC89 为TSWV高感品种而中烟100为耐病品种的原因。此外,NB-ARC结构域是植物疾病抗性基因一个显著的信号结构[10],ARC是信号传导途径下游非常重要的蛋白质因子,参与众多信号途径。大多植物病毒抗性基因含有NBS和LRR结构域,以便与受体结合[11],在番茄斑萎病抗性蛋白A、抗性蛋白B中也均发现该结构。CC-(NB-ARC)-LRR结构是植物抗性蛋白所包含的典型结构[12],番茄斑萎病抗性蛋白A、抗性蛋白B以及番茄线虫抗性基因产物Mi 和 Prf均含有此结构[9],这些细胞内的抗性蛋白识别病原体效应蛋白进而促发一个可能使局部细胞死亡的反应。在番茄中,当TSWV入侵植株时,Sw-5显性个体会引发一个过敏反应,导致TSWV入侵位点或入侵位点周围的细胞快速死亡,使病毒感染不至于达到植株损伤程度,同时也避免了病毒入侵点邻近细胞的感染,防止病毒感染扩散,植株因此表现出对TSWV的抗性[13-14]。推测NtSw-5zy100可能在烟草个体中引发一个相类似的过敏反应,从而使中烟100对TSWV产生耐受性,只出现轻微的花叶或坏死斑点,表现为不被TSWV大面积侵染,而NtSw-5NC89未能引发类似的过敏反应或过敏反应未能引起入侵点周围细胞快速死亡,从而使NC89烟草植株受TSWV侵染后易出现坏死症状。
综上可看出,NtSw-5NC89和NtSw-5zy100均含有抗性蛋白所具有的CC-(NB-ARC)-LRR典型结构,虽然在系统树中与番茄斑萎病抗性蛋白A、抗性蛋白B相对较远,但在结构上却极为相似,这对其在烟草中可能具有的番茄斑萎病抗性奠定了结构基础。番茄中Sw-5包含众多的基因家族成员,各成员编码蛋白除了共有的典型结构域外,也有各自特异之处,不同的同源序列可能对不同的番茄斑萎病毒有抗性[9,14],NtSw-5NC89和 NtSw-5zy100除共有植物抗性蛋白典型结构域外,又互有差别,可能为烟草番茄斑萎病毒抗性基因家族中的一员,在烟草中负责抵抗不同亚型番茄斑萎病毒。
4 结 论
从烟草NC89和中烟100中分别获得疑似抗番茄斑萎病毒基因 NtSw-5NC89和 NtSw-5zy100,NtSw-5NC89长 3819bp,编码1273个氨基酸,NtSw-5zy100长3165bp,编码1054个氨基酸。NtSw-5NC89和NtSw-5zy100均为亲水蛋白,均无明显跨膜区,均无信号肽,均含有植物抗性蛋白所共有的 CC-(NB-ARC)-LRR结构域,二者与假定晚疫病抗性蛋白同源型在系统树中进化关系相近。NtSw-5NC89和 NtSw-5zy100来源于烟草,在烟草番茄斑萎病毒的抗性研究和防治方面有广阔的前景和应用潜力,但对于二者的调控机理和导入烟草植株后的表达状况,还需要进一步的试验验证。
[1] 李丞,李永忠,文国松,等. 烟草番茄斑萎病的研究进展[J]. 中国农学通报,2007,23(2):374-374.
[2] 邱树亮,王孝宣,杜永臣,等. 番茄斑萎病毒TSWV的鉴定及抗病种质的筛选[J]. 园艺学报,2012,39(6):1107-1114.
[3] Ciuffo M, Finetti-Sialer M M, Gallitelli D, et al. First report in Italy of a resistance-breaking strain of Tomato spotted wilt virus infecting tomato cultivars carrying the Sw5 resistance gene[J]. Plant pathology, 2005, 54(4): 564-564.
[4] Gordillo L F, Stevens M R, Millard M A, et al. Screening two Lycopersiconperuvianum collections for resistance to Tomato spotted wilt virus[J]. Plant Disease, 2008, 92(5): 694-704.
[5] Latham L J, Jones R A C. Selection of resistance breaking strains of tomato spotted wilt tospovirus[J]. Annals of Applied Biology, 1998, 133(3): 385-402.
[6] Saidi M, Warade S D. Tomato breeding for resistance to Tomato spotted wilt virus (TSWV): an overview of conventional and molecular approaches[J]. Czech J Genet Plant Breed, 2008, 44: 83-92.
[7] 尼秀媚,陈长法,封立平,等. 番茄斑萎病毒研究进展[J]. 安徽农业科学,2014,42(19):6253-6255.
[8] 谈文,吴元华. 烟草病理学[M]. 北京:中国农业出版社,2003:169-170.
[9] Spassova M I, Prins T W, Folkertsma R T, et al. The tomato gene Sw5 is a member of the coiled coil, nucleotide binding, leucine-rich repeat class of plant resistance genes and confers resistance to TSWV in tobacco[J]. Molecular Breeding, 2001, 7(2): 151-161.
[10] van der Biezen E A, Jones J D G. The NB-ARC domain: a novel signalling motif shared by plant resistance gene products and regulators of cell death in animals[J]. Current Biology, 1998, 8(7): R226-R228.
[11] Folkertsma R T, Spassova M I, Prins M, et al. Construction of a bacterial artificial chromosome (BAC) library of Lycopersiconesculentum cv. Stevens and its application to physically map the Sw-5 locus[J]. Molecular Breeding,1999, 5(2): 197-207.
[12] Pan Q, Wendel J, Fluhr R. Divergent evolution of plant NBS-LRR resistance gene homologues in dicot and cereal genomes[J]. Journal of molecular evolution, 2000, 50(3): 203-213.
[13] Black L L, Hobbs H A, Gatti Jr J M. Tomato spotted wilt virus resistance in Capsicum chinense PI 152225 and 159236[J]. Plant Disease, 1991, 75(8): 863.
[14] Roselló S, Díez M J, Nuez F. Genetics of tomato spotted wilt virus resistance coming from Lycopersicon peruvianum[J]. European Journal of Plant Pathology,1998, 104(5): 499-509.
Screening and the Bioinformatics Analysis of the Nicotiana tabacum L. TSWV-Resistant Genes
YANG Jinguang1, FAN Weihua2,YANG Liuqing2, DING Tao2, WANG Yaofeng3,SONG Yuchuan4, DONG Xin2, WANG Fenglong1, LIU Xiaolu2*
(1. The Tobacco Industry, Tobacco Plant Diseases and Pests Monitoring and Comprehensive Control Key Laboratory, Tobacco Research Institute of CAAS, Qingdao 266101, China; 2. School of Chemistry and Biological Engineering, University of Science and Technology Beijing, Beijing 100083, China; 3. Gansu Tobacco Corporation Qingyang Tobacco Corporation, Xifeng 745000, Gansu,China; 4. Yunnan Tobacco Baoshan Oriental Tobacco Co., Ltd., Baoshan 678000, Yunnan, China)
Two putative TSWV-resistant genes, named NtSw-5NC89and NtSw-5zy100respectively, were isolated from the TSWV-high sensitive Nicotiana tabacum L. NC89 and TSWV- resistant Nicotiana tabacumL.zhongyan 100 using homology-based cloning,. Sequence analysis showed that NtSw-5NC89is 3819 bp long and contains an open reading frame which encodes a polypeptide of 1273 amino acids with the theoretical isoelectric point 5.64 and molecular mass 147529.5 Da. NtSw-5zy100is 3165 bp long and contains an open reading frame which encodes a polypeptide of 1054 amino acids with the theoretical isoelectric point6.75 and molecular mass 122032.9 Da. No signal peptide or transmembrane region was detected in NtSw-5NC89or NtSw-5zy100Both proteins contain a CC-(NBARC)-LRR typical structure which is shared by most plant resistance proteins. Phylogenetic tree analysis showed that NtSw-5NC89had the highest similarity to the Solanum tuberosum putative late blight resistance protein homolog R1A-3 and NtSw-5zy100had the highest similarity to the Nicotiana sylvestrisputative late blight resistance protein homolog R1A-3 isoform X2. Results of this study provide theoretical basis for resistance study, prevention and treatment of Nicotianatabacum L. TSWV.
Nicotiana tabacum L.; TSWV; resistance gene; bioinformatics analysis
S435.72
1007-5119(2016)04-0060-08
10.13496/j.issn.1007-5119.2016.04.011
国家烟草专卖局科技创新平台项目“烟草行业烟草病虫害监测与综合治理重点实验室专项经费”(023201305);云南省烟草公司科技项目“德宏绿色生态优质烟叶生产技术研究及应用”(2013YN37)
杨金广(1979-),博士,副研究员,主要研究方向为分子病毒学与生物防治。E-mail:yangjinguang@caas.cn*通信作者,E-mail:xiaoluliu@ustb.edu.cn
2015-11-12
2016-03-29
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
湖北农业科学(2022年11期)2022-07-18
空间科学学报(2021年1期)2021-05-22
实用肿瘤学杂志(2020年4期)2020-12-08
中国烟草学报(2019年5期)2019-11-14
浙江中西医结合杂志(2017年2期)2017-01-12
中国果菜(2016年9期)2016-03-01
中国蔬菜(2015年9期)2015-12-21
中国老年学杂志(2015年9期)2015-01-31
声屏世界(2014年6期)2014-02-28
