
基于iTRAQ 技术的脂多糖刺激下奶牛乳腺上皮细胞的蛋白组学分析
2016-11-01 11:28王根林
畜牧兽医学报 2016年9期
李 莲,唐 娟,吴 洁,王根林
(南京农业大学动物科技学院,南京 210095)
基于iTRAQ 技术的脂多糖刺激下奶牛乳腺上皮细胞的蛋白组学分析
李莲,唐娟,吴洁,王根林*
(南京农业大学动物科技学院,南京 210095)
基于iTRAQ多重标记串联质谱技术系统研究脂多糖刺激下乳腺上皮细胞蛋白的差异表达,并对与乳蛋白、乳脂肪相关差异表达基因进行验证。培养纯化后的乳腺上皮细胞,经脂多糖(LPS)处理后提取细胞蛋白,采用 iTRAQ 标记和联合二维液相色谱-串联质谱(2D-LC-MS/MS)检测分析得到各组间细胞差异表达蛋白,并采用蛋白印迹和酶联免疫法(ELISA)对差异蛋白进行验证,然后对与乳蛋白、乳脂肪相关基因的相互作用蛋白进行PPI(Protein protein interaction)预测。乳腺上皮细胞共鉴定出 2 487个蛋白,其中差异蛋白共341个,与对照组相比,163个蛋白表达上调,178个蛋白表达下调。iTRAQ测定结果显示,LPS处理组与对照组相比,αs1-酪蛋白(CSN1S1)、κ-酪蛋白(CSN3)和脂肪酸合酶(FASN)基因蛋白表达水平有统计学意义;经蛋白印迹试验结果表明,LPS诱导组αs1-酪蛋白表达水平同对照组比较,明显降低(P<0.05);应用ELISA方法测定细胞培养液中αs1-酪蛋白、κ-酪蛋白和脂肪酸合酶的浓度结果显示,LPS诱导组细胞培养液中上述蛋白的浓度明显低于对照组(P<0.05)。综上表明,LPS诱导的炎症反应可影响乳腺上皮细胞的乳蛋白和乳脂肪的合成。
牛;乳腺上皮细胞;iTRAQ 技术;蛋白质组学;脂多糖
乳腺炎是奶牛养殖行业中造成经济损失最大的疾病之一。从乳腺结构来看,乳腺上皮细胞是乳腺防御病原菌入侵的第一线[1],在免疫应答、局部抵御、合成及分泌乳汁中发挥重要作用[2-4]。对于奶牛临床乳腺炎来说,其中大肠杆菌是奶牛感染临床型乳腺炎的主要病原菌,乳腺中大肠杆菌数量增加越快,乳腺中存在的脂多糖(LPS)越多,就可能更快导致临床型炎症的发生。LPS是革兰阴性菌细胞壁上一种成份[5],广泛存在于自然界致病原。研究表明,LPS 是一种高效的促炎反应因子[6],能够促进动物机体产生急性期反应蛋白和大量的细胞因子,引起机体炎症反应,致使乳腺泌乳营养代谢模式、内分泌调控模式等随之改变,对乳脂肪、乳蛋白的合成和含量有较大的影响[2-4]。本研究通过iTRAQ (Isobaric tag for relative and absolute quantification)技术从蛋白水平研究脂多糖对奶牛乳腺上皮细胞基因表达的影响,并且进一步筛选分析与乳腺上皮细胞泌乳功能相关基因,为揭示LPS对乳腺分泌功能影响的作用机制提供理论支持。
1 材料与方法
1.1材料与试剂
生长培养基(DMEM) 、胎牛血清、胰蛋白酶、二甲基亚砜(DMSO)均购自美国 Gibco公司。FASN抗体购于Santa Cruz 公司。αs1-酪蛋白、κ-酪蛋白抗体、脂多糖(Escherichiacoli0111:B4)购于Sigma公司。αs1-酪蛋白、κ-酪蛋白和脂肪酸合成酶(FASN)ELISA试剂盒购于上海劲马生物技术公司。电热恒温 CO2培养箱(美国 Thermo) 、倒置显微镜(CKX41,日本 Olympus) 、超低温冰箱 (美国 Thermo) 、冷冻离心机(5415R 型,德国 Eppendorf)等。
1.2方法
1.2.1乳腺上皮细胞的获取自南京市附近的屠宰场采集3头屠宰后的健康泌乳期奶牛的乳腺组织,所采集3头奶牛情况:1头泌乳中期(因吞食了铁钉,铁钉刺穿胃壁刺到心包膜),2头泌乳后期(由于产量低而被淘汰)。所采集的乳腺组织,均有乳汁分泌。奶牛屠宰后立即采用无菌术采集奶牛乳腺组织,并将其置于无菌的PBS中,带回实验室备用。将采集的乳腺组织采用酶消化法获取乳腺上皮细胞,根据成纤维细胞与上皮细胞对胰蛋白酶耐受时间的不同及 2 种细胞贴壁速度的差异分离和纯化乳腺上皮细胞。对细胞中角蛋白-18 进行鉴定[10]。纯化的奶牛乳腺上皮细胞接种于含10%胎牛血清的DMEM培养基中,在5% CO2和37 ℃的条件下贴壁生长,每隔24 h更换培养基。用 0.25%胰蛋白酶消化进行传代,倒置显微镜观察细胞生长状况。
1.2.2脂多糖处理乳腺上皮细胞待细胞铺满培养板90%时,更换为无血清培养液,并加入试剂处理。试验分为对照组( Con)和脂多糖处理组(10 μg·mL-1),处理细胞24 h后,收集细胞上清液,刮取细胞,置于-80 ℃冰箱保存。脂多糖处理组浓度参照文献[7]确定。然后将来自于3头奶牛的乳腺上皮细胞混合备用。
1.2.3蛋白质提取消化将3头奶牛的细胞样品用液氮磨成干粉;用 200 μL 的 TEAB 溶解上述干粉,然后加入 4 倍体积的冷丙酮(含终浓度为 10 mmol·L-1DTT)沉淀2 h;13 000 r·min-1离心 20 min,收集沉淀;加入800 μL 的冷丙酮(含终浓度为 10 mmol·L-1DTT)重悬沉淀;13 000 r·min-1离心 20 min,收集沉淀,然后风干沉淀;加入 100 μL TEAB 溶解蛋白。采用选择常规的 Bradford 定量方法定量总蛋白质。混合3头奶牛中对照组和试验组的细胞蛋白,然后对照组和试验组分别作2个技术重复,共4份样品。
1.2.4iTRAQ标记及高效液相色谱法分离肽段提取的总蛋白质进行酶切和除盐后,采用4-plex进行标记(武汉金开瑞生物工程有限公司)。iTRAQ 试剂标记蛋白按操作手册的步骤进行:取出 iTRAQ试剂,使其恢复至室温,每份 iTRAQ 试剂加 70 μL 乙醇,涡旋,离心;在4份样品内分别加入113、114、117和118 iTRAQTM试剂,涡旋,离心,室温孵育1 h;混合 iTRAQ 试剂标记的蛋白,加入同一试管,混匀涡旋,离心。混合标记蛋白前,需确认所消化的蛋白是否已成功标记 iTRAQ 标记物:所标记样品经纯化后用ABI 4700 进行标记验证,如果标记成功,可见到所标记的报告部分( 本研究中选用113-114为对照组报告部分,117-118 为处理组报告部分);若未检测到相应的峰值则需要重新标记)。标记后等量混合,反相柱分离。
1.2.5LC-MS/MS 质谱检测与数据处理将样品通过LC-MS-MS进行蛋白质检测。对质谱数据用ProteinPilotTM软件进行检索和筛选。 对于proteinpilot的鉴定结果,需进一步过滤,对于鉴定到的蛋白,unused score≥1.3(即可信度水平在95%以上),每个蛋白至少包含一个unique肽段的蛋白为可信蛋白,不符合该条件的蛋白不包括在结果中;对于鉴定的肽段,可信度在95%以上认为可信肽段,对于蛋白质定量,为了得到更全面的关于某蛋白的定量信息,Proteinpilot软件使用了conf≥15的所有肽段,不符合该条件的肽段不包含在结果中。
使用Proteinpilot软件对蛋白质组 iTRAQ定量。当差异倍数达到 1.3倍及以上(即上调≥1.3倍和下调≤0.77倍),且经过显著性统计检验其P-value值≤0.05时,视为显著差异蛋白。运用富集分析算法对初步鉴定的蛋白数据进行分组,鉴定的蛋白与数据库进行比对,选择GO(Gene Ontology) 的生物过程、细胞成分和分子功能注释对蛋白进行分类。1.2.6ELISA检测细胞培养液中αs1-酪蛋白、κ-酪蛋白及脂肪酸合成酶(FSAN)浓度检测细胞培养液中αs1-酪蛋白、κ-酪蛋白和脂肪酸合成酶(FSAN)水平,按照试剂盒(上海劲马生物有限公司)说明书进行测定。1.2.7乳腺上皮细胞αs1-酪蛋白表达提取乳腺上皮细胞蛋白,测定浓度后,将样品在5%~10% SDS-PAG中电泳(浓缩胶80 V;分离胶120 V)2 h后,转印于硝酸纤维素膜上(100 mA 45 min)。硝酸纤维素膜经封闭液(5%脱脂乳)封闭1.5 h后,TBS洗涤3次,每次10 min,于αs1-酪蛋白一抗工作液(稀释比例1∶1 000)中孵育过夜。TBS洗涤3次,用辣根过氧化物酶标记的二抗工作液(稀释比例1∶2 000)孵育1.5 h。TBS洗涤3次,超敏发光液显光,感光X光片后,显影定影。β-tublin蛋白作为内参蛋白。1.2.8相互作用蛋白质组预测蛋白质很少单独发挥作用,本研究选择了与乳腺脂肪和蛋白合成相关的5个目标蛋白质,对检测到的蛋白进行PPI(Protein-protein interaction)预测,绘制出互作关系网络图。
1.3统计分析
使用SPSS 19.0软件对数据进行统计分析。所
有数据用“平均数±标准差(x±s)”表示,组间比较采用t检验。P<0.05 表示差异显著。
2 结 果
2.1差异蛋白及GO 功能分类
乳腺上皮细胞总共鉴定出 2 487个蛋白;蛋白芯片所获得的数据使用绝对值≥1.3检出,其中检出差异蛋白共341个,与对照组相比163个蛋白表达上调,178个蛋白表达下调。其中与炎症和免疫相关的有代表性的上调差异蛋白是增殖细胞核抗原(PCNA)、前列素内环氧化物合成酶2(PTGS2/COX2)、血红素加氧酶1(HO-1,Heme oxygenase 1)和免疫共刺激分子CD276等。与乳脂肪和乳蛋白合成相关的有代表性的下调差异蛋白如αs1-酪蛋白(CSN1S1)、κ-酪蛋白(CSN3)和FASN。其中上调的基因还有与热休克相关的热休克蛋白70(HSPA13)。将341个差异蛋白质按照蛋白质分子功能、生物过程和信号通路分别进行分析。其中P<0.05的生物学过程共177条,筛选其中与炎症以及合成代谢相关的过程 (图1和图2)。与炎症相关的8条生物学过程中,其中炎性反应过程2条,防御反应过程1条,其余5条均与免疫反应相关。与合成和代谢相关的18条生物学过程中,其中14条与代谢过程有关,4条与生物合成有关。

A.炎症反应;B.急性炎症反应;C.防御反应;D.免疫系统过程;E.免疫系统过程调控;F.免疫应答的调控;G.体液免疫应答;H.免疫系统过程正调控A.Inflammatory response;B.Acute inflammatory response;C.Defense response;D.Immune system PROCRSS;E.Regulation of immune system proecss;F.Regulation of immune response;G.Humoral immune response图1 与炎症相关的生物学过程Fig.1 Biological processes associated with inflammation

A.类固醇代谢进程的调控;B.甘油脂代谢过程;C.含硫氨基酸的代谢过程;D.细胞蛋白质代谢过程的正调控;E.多糖代谢过程;F.蛋白质代谢过程的正调控;G.不饱和脂肪酸的代谢过程;H.脂代谢过程;I.核苷磷酸代谢过程;J.磷脂代谢过程;K.有机磷代谢过程;L.氨基聚糖分解代谢过程;M.细胞脂质代谢过程;N.肪酸生物合成过程;O.细胞因子生物合成过程;P.黏多糖的生物合成过程;Q.核苷磷酸生物合成过程;R.糖胺聚糖的生物合成过程A.Regulation of sterid metabolic process;B.Glycerolipid metabolic process;C.Sulfur amino acid metbolic process;D.Positive regulation of celluar protein metabolic process;E.Polysaccharide metabolic process;F.Positive regulation of protein metabolic process;G.Unsaturated fatty acid metabolic process;H.Lipid metabolic process;I.Nucleosiede monophosphate metabolic process;J.Phospholipid metabolic process;K.Organophosphate metabolic process;L.Aminoglcan catabolic process;M.Cellular lipid metabolic process;N.Fatty acid biosynthetic process;O.Cytokine biosynthetic process;P.Glycosaminoglycan biosynthetic;Q.Nucleside monophosphate process;R.Aminoglycan biosynthetic process图2 与合成和代谢相关的生物学过程Fig.2 Biological processes related to synthesis and metabolism
2.2乳腺上皮细胞中αs1-酪蛋白的表达
LPS处理组乳腺上皮细胞αs1-酪蛋白表达水平较对照组降低,两组比较差异显著(P<0.05),与 MS鉴定试验中αs1-酪蛋白下调的结果一致。
2.3乳腺上皮细胞培养上清中αs1-酪蛋白、κ-酪蛋白和脂肪酸合成酶(FASN)的浓度
LPS处理组乳腺上皮细胞培养上清中αs1-酪蛋白、κ-酪蛋白的含量较对照组明显降低,两组比较差异极显著(P<0.01,图4A和B),与质谱鉴定试验中αs1-酪蛋白和κ-酪蛋白下调的结果一致。LPS处理组乳腺上皮细胞脂肪酸合成酶的活性显著低于对照组(P<0.05,图4c),且与质谱鉴定试验结果一致。
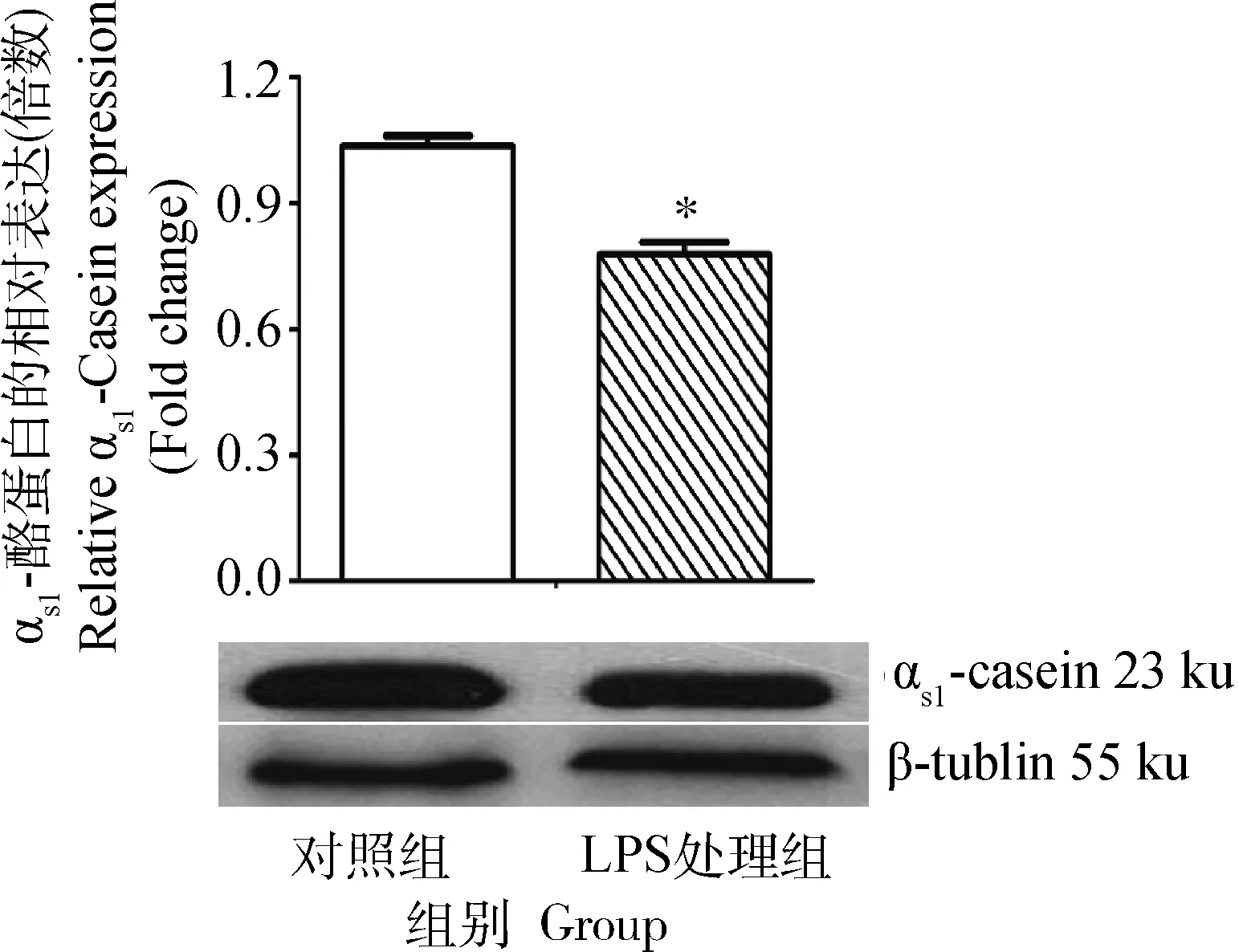
图3 乳腺上皮细胞αs1-酪蛋白的表达Fig.3 The expression of αs1-casein in BMEC

图4 培养上清中αs1-酪蛋白、κ-酪蛋白和脂肪酸合成酶的浓度Fig.4 The concentration of αs1-casein,κ-casein and fatty acid synthase in culture medium
2.4与乳蛋白和乳脂肪合成相关蛋白的互作网络
蛋白质很少单独发生作用,在所有生物功能中,通常以蛋白质复合物的功能占主导地位。为了进一步了解蛋白质的功能,以与乳蛋白(CSN、LTA、EIF4E)和乳脂肪合成(FASN、CD36)相关的5个蛋白为目标蛋白质(图中蓝色边框V点代表目标蛋白质)进行了PPI预测。如图5为其中3个蛋白的质谱图。另如图6所示,图中共有271个点,形成的两两互作关系有966个。其中下调基因用绿色表示,上调基因用红色表示。在此网络图的5个目标蛋白中连接度(Degree)和压力向心性最高的基因是“EIF4E”,随后为CD36、LTF、FASN和CSN3。由于连接度值越大,反映此节点在网络中的重要性越大,而压力向心性值越大,说明此节点越可能成为网络压力响应中的关键点,所以EIF4E在5个目标蛋白质中重要性最大。另外与脂肪相关的CD36和

A.用于鉴定CD36的肽段EVVLEEGTIAFK的MS/MS图谱;B.用于鉴定FASN的肽段ACVDTALENMTSLK的MS/MS图谱;C.用于鉴定EIF4E的肽段WLITLNK的MS/MS图谱。对照组细胞用iTRAQ 试剂113和114标记,LPS处理组细胞用iTRAQ reagent 117和118标记A.Mass spectra for identification the EVVLEEGTIAFK peptide fragment of CD36;B.Mass spectra for identification the ACVDTALENMTSLK peptide fragment of FASN;C.Mass spectra for identification the WLITLNK peptide fragment of EIF4E .The control groups were labeled by iTRAQ reagent 113 and 114,LPS-treated groups were labeled by iTRAQ reagent 117 and 118.图5 用于鉴定和定量CD36、FASN和EIF4E的MS/MS质谱图Fig.5 Mass spectra for identification and quantification of CD36,FASN和EIF4E

图中共有271个点(基因),966个互作关系,点的颜色随差异基因变化倍数的增大由绿色(下调)-白色-红色(上调)渐近变化,即颜色越深代表变化倍数越大;中心节点(degree最大)的点最大;基因之间的互作关系可信度越高,节点间的连线越粗;蓝色边框V点代表目标蛋白质There were 271 points(gene)and 966 interactions between these genes in figure 6. The color of points was asymptotic change from green(down-regulated)-white-red(up-regulated) according to the fold change of different genes,darker represents a higher fold change; the central node(maximum degree) was the maximum point; the higher credibility of interaction between genes was, the thicker lines between the nodes was; the blue border point V represents the target protein图6 互作关系图Fig.6 Protein-protein interaction network
FASN之间具有一定的互作关系(可信度得分404),而CSN3与其他两个候选蛋白LTF和EIF4E则无互作关系。
3 讨 论
近年来,蛋白组学提供了一个有效的方法来发现乳腺炎中不同蛋白的表达以及为发现生物标记物和药物靶点的基础理论[8]。运用iTRAQ技术并结合液相质谱分析健康和乳房炎乳腺组织在炎症反应后期的蛋白表达情况,通过该研究揭示与宿主组织损伤的有关机制[9]。使用大肠杆菌内毒素(LPS)处理乳腺组织导致乳腺快速炎症来作为模型研究奶牛乳房的炎症反应[6]。但从乳腺的结构来看,乳腺上皮细胞是乳腺防御病原菌入侵的第一道防线,所以本研究是通过LPS处理乳腺上皮细胞后观察其蛋白质表达谱的变化,从而从细胞水平揭示大肠杆菌引发乳腺炎后期对乳蛋白及乳脂肪合成的影响机制。
LPS处理乳腺上皮细胞后,引起了细胞的炎症反应。从GO分析中可以看出,LPS激活了与炎症相关的8条生物学过程,且其中炎性反应过程2条,防御反应过程1条,其余5条均与免疫反应相关。所以本研究中10 μg·mL-1的脂多糖处理组可引发乳腺上皮细胞的炎症反应[7]。在奶牛乳腺组织分泌的乳汁中,常见的酪蛋白4 种分别为αs、β、κ 和 γ,其中αs酪蛋白占总酪蛋白的40%以上,β-酪蛋白占总酪蛋白的25%,κ-酪蛋白占总酪蛋白的9%,γ-酪蛋白占总酪蛋白的1%[10]。本研究质谱结果表明,其中αs1-酪蛋白(0.74倍)、κ-酪蛋白(0.47倍)和β-酪蛋白(0.8倍)表达水平降低,而αs2-酪蛋白(1.0倍)表达水平无显著变化,通过蛋白印迹和ELISA也对结果加以验证。S.Schmitz等对奶牛乳腺组织LPS处理后9 h后,发现αs1-酪蛋白、αs2-酪蛋白和β-酪蛋白基因 mRNA表达水平下降,但差异不显著,而κ-酪蛋白基因mRNA表达显著下降(P<0.05)[6]。本研究结果与上述研究结果不完全一致,这可能是由于LPS直接处理乳腺上皮细胞对上皮细胞作用更直接的缘故导致。
LPS 可通过调控乳腺组织中乳脂肪合成的关键酶,如脂肪酸合成酶和乙酰辅酶羧化酶等[11],下调脂蛋白脂肪酶的活性[12]。诱导反刍动物发生瘤胃酸中毒,发现释放的LPS改变了乳脂肪的生产和能量的利用效率[13-14]。本研究发现经LPS处理后,乳腺上皮细胞脂肪酸合成关键酶脂肪酸合成酶(FASN)蛋白表达水平下降。另外在脂肪酸的摄取转运过程中,其中主要是脂肪酸转位酶(CD36)和质膜蛋白脂肪酸结合蛋白(FABP)来调控[15]。本研究质谱的结果表明脂肪酸转位酶(CD36)和质膜蛋白脂肪酸结合蛋白5(FABP5)表达水平均下降,分别为0.74倍和0.58倍。综上表明,LPS不仅影响脂肪酸的合成,而且对脂肪酸的摄取和转运过程产生影响。这与GO分析得出的差异基因参与合成和代谢的生物学过程结果相一致。
本研究在筛选了与乳蛋白和乳脂肪相关的基因后,以目标基因为中心进一步对相互作用蛋白质组进行PPI(Protein-Protein Interaction)预测,从而找出这些基因之间的互作关系网络。从本研究的互作关系图来看,所选的5个与乳脂肪和乳蛋白相关的目标蛋白质中,只有两个CD36和FASN有直接的互作关系,而其他3个蛋白间无直接的互作关系(CSN3、LTF和EIF4E),但EIF4E的连接度(Degree)和压力向心性最高。由于EIF4E是mTOR信号通路中重要的翻译起始因子,其磷酸化水平决定该通路介导的蛋白合成过程,故可以对该基因做进一步研究[16]。
4 结 论
本研究结果表明,LPS诱导的炎症反应可影响乳腺上皮细胞的乳蛋白和乳脂肪的合成。与脂肪转运和合成相关的CD36和FASN蛋白之间有直接的互作关系,而且EIF4E是mTOR信号通路介导的蛋白合成过程中重要的翻译起始因子,其与多种蛋白具有互作关系。本研究为乳腺炎影响乳腺分泌的机制提供了新的证据。
[1]GENINI S,BADAOUI B,SCLEP G,et al.Strengthening insights into host responses to mastitis infection in ruminants by combining heterogeneous microarray data sources[J].BMCGenomics,2011,12(1):225.
[2]GRIESBECK-ZILCH B,OSMAN M,KÜHN C,et al.Analysis of key molecules of the innate immune system in mammary epithelial cells isolated from marker-assisted and conventionally selected cattle[J].JDairySci,2009,92(9):4621-4633.
[3]BANNERMAN D D,PAAPE M J,LEE J,et al.EscherichiacoliandStaphylococcusaureuselicit differential innate immune responses following intramammary infection[J].ClinDiagnLabImmun, 2004,11(3):463-472.
[4]RIOLLET C,RAINARD P,POUTREL B.Differential induction of complement fragment c5a and inflammatory cytokines during intramammary infections withEscherichiacoliandStaphylococcusaureus[J].ClinDiagnLabImmun,2000,7(2):161-167.
[5]BEUTLER B,RIETSCHEL E T.Innate immune sensing and its roots:the story of endotoxin[J].NatRevImmunol,2003,3(2):169-176.
[6]SCHMITZ S,PFAFFL M W,MEYER H H D,et al.Short-term changes of mRNA expression of various inflammatory factors and milk proteins in mammary tissue during LPS-induced mastitis[J].DomesAnimEndocrinol,2004,26(2):111-126.
[7]SUN Y,LI L,WU J,et al.Bovine recombinant lipopolysaccharide binding protein (BRLBP) regulated apoptosis and inflammation responsein lipopolysaccharide-challenged bovine mammary epithelial cells (BMEC)[J].MolImmunol,2015,65(2):205-214.
[8]LIPPOLIS J D,BRUNELLE B W,REINHARDT T A,et al.Proteomic analysis reveals protein expression differences inEscherichiacolistrains associated with persistent versus transient mastitis[J].JProteomics,2014,28(108):373-381.
[9]HUANG J,LUO G,ZHANG Z,et al.iTRAQ-proteomics and bioinformatics analyses of mammary tissue from cows with clinical mastitis due to natural infection withStaphylococciaureus[J].BMCGenomics,2014,2(15):839.
[10]PENASA M,DE MARCHI M,CASSANDRO M.Short communication:Effects of pregnancy on milk yield,composition traits,and coagulation properties of Holstein cows[J].JDairySci,2016,99(6):4864-4869.
[11]LOPEZ-SORIANO F J,WILLIAMSON D H.Acute effects of endotoxin (lipopolysaccharide) on tissue lipid metabolism in the lactating rat.The role of delivery of intestinal glucose[J].MolCellBiochem,1994,141(2):113-120.
[12]KHOVIDHUNKIT W,KIM M S,MEMON R A,et al.Thematic review series:The pathogenesis of atherosclerosis.Effects of infection and inflammation on lipid and lipoprotein metabolism mechanisms and consequences to the host[J].JLipidRes,2004,45(2):1169-1196.
[13]ANDERSEN P H.Bovine endotoxicosis——some aspects of relevance to production diseases.A review[J].ActaVetScand,2003,44(Suppl 1):S141.
[14]AMETAJ B N,BRADFORD B J,BOBE G,et al.Strong relationships between mediators of the acute phase response and fatty liver in dairy cows[J].CanJAnim, 2005,85(2):165-175.
[15]DOEGE H,STAHL A.Protein-mediated fatty acid uptake:Novel insights frominvivomodels[J].Physiology(Bethesda),2006,21(4):259-268.
[16]QIN X,JIANG B,ZHANG Y.4E-BP1,a multifactor regulated multifunctional protein[J].CellCycle,2016,15(6):781-786.
(编辑程金华)
Analysis of Expression Protein Profiles of Bovine Mammary Epithelial Cell Based on Isobaric Tag for Relative and Absolute Quantification Proteomics and Bioinformatics Technique
LI Lian,TANG Juan,WU Jie,WANG Gen-lin*
(CollegeofAnimalScienceandTechnology,NanjingAgriculturalUniversity,Nanjing210095,China)
The aim of the present study was to screen differential expression proteins associated with inflammatory model of bovine mammary epithelial cells (BMECs) induced by lipopolysaccharide (LPS) using isobaric tags for relative and absolute quantification (iTRAQ) combined with 2-dimensional liquid chromatography-tandem mass spectrometry (2D-LC-MS/MS).Normal and LPS-treated bovine mammary epithelial cells were analyzed using iTRAQ combined with 2D-LC-MS/MS.Bioinformatics analyses of differentially expressed proteins were performed by means of Gene Ontology and PPI (Protein protein interaction)analysis.According to iTRAQ results,a total of 2 487 proteins were identified.Compared with normal BMECs,163 proteins were significantly up-regulated while 178 proteins were significantly down-regulated in response to LPS treatment.The bioinformatics analyses highlighted the effects of LPS treatment on milk proteins and fat synthesis such as αs1-casein,κ-casein and fatty acid synthase.Down-regulation of αs1-casein (CSN1S1) in BMECs was confirmed by Western blotting;αs1-casein,κ-casein (CSN3) and fatty acid synthase (FASN) in culture medium were confirmed by ELISA.The results showed that LPS treatment significantly decreased the expression of αs1-casein in BMECs (P<0.05),and also decreased the concentration of αs1-caseins,κ-casein and fatty acid synthase in culture medium (P<0.05).In conclusion,LPS treatment could affect the synthesis of protein and fat in BMECs.
bovine;mammary epithelial cell;iTRAQ technique;proteomics;LPS
10.11843/j.issn.0366-6964.2016.09.014
2016-04-18
国家自然科学基金(31372290);中央高校基本科研业务费专项资金(KJQN201607)
李莲(1979-),女,内蒙古人,讲师,硕士,主要从事奶牛以及动物生殖生理方面的研究,E-mail:lilian@njau.edu.cn
王根林,教授,主要从事动物生殖生理及调控研究,E-mail:glwang@njau.edu.cn
S823.2
A
0366-6964(2016)09-1853-08
猜你喜欢
今日农业(2022年3期)2022-11-16
中国乳业(2022年4期)2022-05-06
中国饲料(2022年5期)2022-04-26
今日农业(2021年10期)2021-07-28
中国生殖健康(2020年5期)2021-01-18
小天使·一年级语数英综合(2019年12期)2019-01-13
中成药(2018年12期)2018-12-29
中国生殖健康(2018年5期)2018-11-06
中国酿造(2016年12期)2016-03-01
中国洗涤用品工业(2015年8期)2015-02-28
