
桂西北喀斯特峰丛洼地典型灌丛植物种间关系及环境解释
2016-12-17 09:06郑生猛盖爽爽谢强苏以荣陈香碧胡亚军何寻阳
农业现代化研究 2016年6期
郑生猛,盖爽爽,谢强,苏以荣,陈香碧,胡亚军,何寻阳*
(1.广西师范大学生命科学学院,广西 桂林 541004;2.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南 长沙 410125; 3. 中国科学院环江喀斯特生态系统观测研究站, 广西 环江 547100)
桂西北喀斯特峰丛洼地典型灌丛植物种间关系及环境解释
郑生猛1,2,3,盖爽爽1,2,3,谢强1,苏以荣2,3,陈香碧2,3,胡亚军2,3,何寻阳2,3*
(1.广西师范大学生命科学学院,广西 桂林 541004;2.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南 长沙 410125; 3. 中国科学院环江喀斯特生态系统观测研究站, 广西 环江 547100)
郑生猛, 盖爽爽, 谢强, 苏以荣, 陈香碧, 胡亚军, 何寻阳. 桂西北喀斯特峰丛洼地典型灌丛植物种间关系及环境解释[J].农业现代化研究, 2016, 37(6): 1198-1205.
Zheng S M, Gai S S, Xie Q, Su Y R, Chen X B, Hu Y J, He X Y. The interspecific associations between dominant species and their environmental interpretation under a typical shrub community in karst peak-cluster depression regions of Northwest Guangxi[J]. Research of Agricultural Modernization, 2016, 37(6): 1198-1205.

喀斯特峰丛洼地典型灌丛群落物种种间关系分析对于深入认识群落结构特征及群落动态,探讨喀斯特灌丛物种共存机制具有重要的意义。本研究在详细的野外调查基础上,基于2×2列联表,采用方差比率法、χ2检验、Pearson相关系数检验和Spearman秩相关系数检验等方法,对桂西北典型喀斯特灌丛群落的30个木本植物优势种进行了种间关联和相关性分析;结合冗余分析(RDA)进行生态种组划分。结果表明,用来检测VR值偏离1的显著性W=195.31 >χ20.05(100),说明群落总体间呈显著正关联;χ2检验中种对间正负关联比为1.38,反映群落种间联结性较强。种间相关分析中,Pearson相关系数检验与Spearman秩相关系数检验种间相关显著率分别为42.99%,51.03%,说明Spearman秩相关系数检验的灵敏度最高,更能反映群落种间相关性。根据30个优势种的环境适应性和主导生态因素,结合RDA排序,可将它们划分为3个生态种组,分别为黄荆生态种组,米念芭、三脉叶荚蒾生态种组,小芸木生态种组。在喀斯特植被恢复和植树造林的实践中,应充分考虑物种的生态习性和喀斯特生境的独特性,利用种间关系合理选择同一生态种组物种进行搭配种植。
喀斯特;灌丛群落;优势种;种间关系;生态种组
植物群落由时间上和空间上彼此相互关联的共存物种组成[1-2],物种间的相互关系使群落处于相对稳定的状态[3]。种间关系是植物群落重要的数量和结构特征之一[4-6],是对各个物种在不同生境中相互影响相互作用所形成的有机联系的反映,对群落结构的形成和动态演化起着重要作用[7-8]。研究种间关系对正确认识群落的结构、功能和分类有着重要的指导意义[9],并能为植被的经营管理、自然植被恢复和生物多样性保护提供理论依据[10-12]。种间联结和种间相关是分析种间关系的主要方法[13]。种间联结是以物种是否存在的二元数据为基础,属于定性测定;而种间关联是对物种的数量数据进行定量分析[14]。两者结合能够客观地测定物种间的关联程度,并能有效地反映物种间的相互作用和群落中物种的配置情况[15]。国内外学者结合群落结构[16]、种间竞争[17-19]、生态种组[20-21]、群落演替[22]等对不同植被类型进行了大量有关种间关系的研究,但有关喀斯特地区植物种间关系的研究较少[23],而针对喀斯特灌丛木本优势种的种间关系研究更鲜见报道。
灌丛是喀斯特地区的主要植被类型之一,也是植被演替的重要过渡阶段[30]。在国家对石漠化地区大力进行生态恢复的背景下,对干扰后处于封育阶段的典型灌丛进行优势种种间联结性和关联分析,有助于深入了解自然恢复过程中喀斯特灌丛群落结构及动态,探索灌丛群落优势物种对喀斯特异质性生境的响应及环境对种间关系的影响,根据物种的生态习性进行生态种组划分,进一步为喀斯特地区的植树造林及植被保护提供理论参考。
本文通过对桂西北典型喀斯特峰丛洼地灌丛主要木本植物进行种间关联分析,了解喀斯特灌丛优势种之间的种间联结性和关联性现状,探索优势种在喀斯特生境异质性对灌丛群落种间关系的影响,深入认识喀斯特灌丛群落结构特征及群落动态,探讨喀斯特灌丛群落种间关系的维持机制。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于桂西北典型喀斯特峰丛洼地——广西壮族自治区环江毛南族自治县(107°51′-108°43′E,24°44′-25°33′N)。原生植被为亚热带常绿阔叶林,成土母岩以砂页岩和石灰岩为主。该地区属亚热带季风气候,雨热同季,年平均气温16.5-19.9℃,1月平均气温10.1℃,7月平均气温28℃,无霜期长,年平均降雨量为1 389-1 750 mm,集中于4-9月,占全年降雨量的70%,平均蒸发量为1 571.1 mm,相对湿度平均为70%[25]。喀斯特峰丛洼地集中分布在该县的西南部,土壤分布不连续,土层浅薄,岩石裸露率高,坡度大,水土流失严重。灌丛是常见的植被类型,其主要优势种有黄荆(Vitex negundo)、火棘(Pyracantha fortuneana)、盐肤木 (Rhus chinensis)和红背山麻杆(Alchornea trewioides)等。
1.2 研究方法1.2.1 样地设置及数据采集 在充分勘察的基础上,选择位于广西环江毛南族自治县中国科学院环江喀斯特生态系统观测站内一个受干扰退化,经封育后自然恢复良好的典型喀斯特峰丛洼地坡面灌丛,设置一块面积为1 hm2(100 m×100 m)的永久监测样地。用Topcon全站仪将样地划分为100个10 m×10 m的相邻网格样方。以样地的西南角为原点,东西方向为横轴(x),南北方向为纵轴(y),以确定水平坐标。将每个10 m×10 m的样方进一步划分为4个5 m×5m的小样方,依次对每个小样方进行群落学调查。调查样方内胸径(DBH)≥1 cm的木本植物(包括藤本),挂牌并记录其种名、个体相对坐标、胸径、树高、冠幅、生长状况等指标;对于胸径DBH<1 cm的乔木和灌木幼苗以及草本层植物,简要记录其种类组成、株数、高度、盖度等特征。
以每个10 m×10 m的样方为单位,测量海拔、坡度、岩石裸露率等指标。按“S”型路线采用多点(10个点)混合取样法采集表层土壤(0-15 cm)样品,分析土壤的理化性质。土壤样品室内分析指标包括pH、有机质(SOM)、全氮(TN)、全磷(TP)、全钾(TK)、碱解氮(AN)、速效磷(AP)、速效钾(AK)等。以上指标的测定采用鲍士旦《土壤农化分析》的方法进行[26]。
1.2.2 数据处理 1) 重要值计算。由于植物种类较多,为了计算方便,对重要值≥1的30个优势物种(表1)进行种间联结性和关联性的计算和分析。重要值=(相对多度+相对频度+相对盖度)/3。
2)总体关联性。根据物种是否在样方内出现整理出样方二元数据矩阵作为原始数据,采用方差比率法,通过计算物种间联结指数(VR)[27]来测定所研究的整个群落的总体关联性,用W检验关联的显著性[3,28-29]。计算公式为 :

表1 优势种的编号及重要值Table 1 Number and important value of dominant species
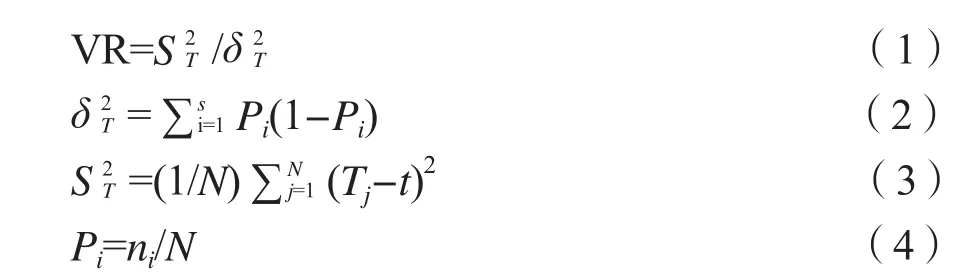
式中:S为物种总数;Pi为物种i出现的频度;N为样方总数;Tj为样方j内出现的研究物种总数,ni为物种i出现的样方数,t为所有样方中种的平均数,t=(T1+T2+…+Tn) /N;ST为所有样方物种数的方差,δT为所有物种出现频度的方差[6]。在独立性假设条件下,VR期望值为1,当VR=1时,接受种间总体无关联的零假设;VR>1时,否定零假设,表明物种间总体呈现正关联;VR<1时,表明物种间存在负关联。采用统计量W检验VR值偏离1的显著程度,W=N×(VR)。W符合χ2分布(df=N-1)。若χ2
0.95(N)<W<χ20.05(N),则物种总体间关联不显著(P>0.05)。若W<χ20.95(N)或W>χ2
0.05(N),则物种总体关联显著(P<0.05)。
3)种间联结性。种间联结一般采用χ2检验进行定性研究,根据2×2列联表的χ2统计量测定成对种间的联结性[30],将30个优势物种是否在100个标准样方中出现转化为100×30的0、1二元数据矩阵,0表示物种在样方中未出现,1表示出现。依据上述原始数据矩阵,构造30个优势种群435个种对的定性数据列入2×2 联列表,计算出a、b、c、d的值[31]。
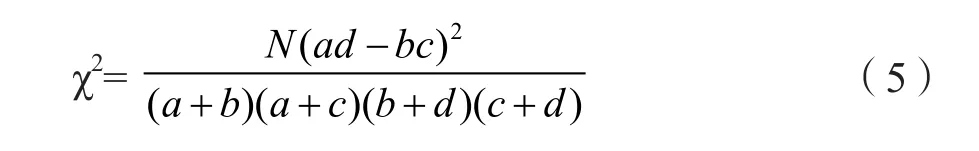
式中:N为总样方数;a为2个物种同时出现的样方数,b、c分别为仅有1个物种出现的样方数,d为2个物种均未出现的样方数。通常P>0.05,即χ2<3.841时,认为两个种彼此独立分布,即中性联结;0.01<P<0.05,即3.841<χ2<6.635时,认为种间联结显著;P<0.01即χ2>6.635时,认为种间联结极显著。当ad>bc时为正联结,ad<bc则为负联结。
当某一物种的频度为100%时,b,d值均为0,这样就无法做χ2检验。所以,应给该种一个加权值,令式中的b,d值均为1,就可以较客观地反映2个种的联结情况[31]。
4)种间相关性测定。应用定量数据(重要值)对种间关系进行Pearson相关系数和Spearman秩相关系数分析[6]。
Pearson相关系数计算公式如下:
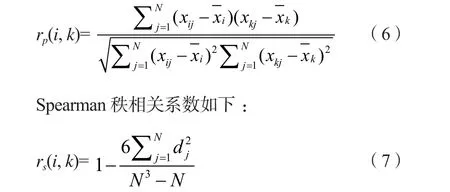
式中:rp(i, k) 为Pearson相关系数,rs(i, k)为Spearman秩相关系数,N为总样方数,dj=(xij-xkj),xij和xkj分别为种i和种k在样方j中的秩。
5)冗余分析(RDA)。环境因子对种间关系的影响采用RDA分析。RDA可以结合多个环境因子进行多元梯度分析,能够较好地反映物种与环境的关系。本文选用30个优势物种的重要值建立30×100重要值数据矩阵和13×100的环境因子(含地形和土壤养分因子)矩阵进行RDA分析。RDA分析采用生态学软件Canoco 4.5进行。
6)生态种组的划分。群落中生态习性相近的物种可以归类为一生态种组。群落内的种间关系揭示了不同种类因生境差异而表现出的相互关系。同一生态种组的物种,在环境适应、资源利用及群落功能等方面表现出一致性,组内种间关系常为正相关。某一生态种组的缺失将会对群落的结构和功能产生重要的影响[11]。为了真实的反映不同种群对生境条件的适应性,以种对间负相关为界限,同一组内尽量多的正相关种对为原则,结合2×2列联表的χ2统计检验、Pearson相关系数和Spearman秩相关系数分析,及物种的自身生物学特性,利用RDA排序结果,对30个优势种进行生态种组划分。
2 结果与分析
2.1 主要物种总体间关联
根据30个主要物种的存在与否的二元数据矩阵,计算出样地典型喀斯特灌丛群落的总体关联性(表2)。VR>1,说明30个物种之间表现出净的正联结。用统计量W检测VR值偏离1的显著性,结果显示W>χ20.05(100)[32],说明30个主要物种之间在总体上表现出显著正关联。反映出喀斯特典型灌丛群落主要物种间关联度高,群落整体关系紧密,种间关系逐步趋于正相关。

表2 主要物种间的总体关联性Table 2 Macro-association among the population of 30 dominant shrub species
2.2 种对间关联性分析
对30个主要木本植物采用 χ2检验进行研究发现(图1),435个种对间有249对表现正关联,占57.70%,其中极显著正关联物种对有77对,显著正关联对数有23对,占总对数的22.99%;179对表现负关联,占41.15%;其中极显著负关联种对数有63对,显著负关联对数有19对,占总对数的18.85%。总体间正负关联比为1.38,有5对表现出无关联,种间关联显著率达41.84%。存在极显著正关联的有米念芭与三脉叶荚蒾、三脉叶荚蒾与火棘等77个物种对。米念芭与南岭榨木、刺壳花椒与八角枫等23个物种对呈显著正关联。正关联达到显著水平的物种多具有相似的生境需求,它们在样地中常伴随出现。存在极显著负关联的有米念芭与小蜡、刺壳花椒与三脉叶荚蒾等63个物种对。米念芭与石山柿、刺壳花椒与竹叶花椒等19个物种对呈显著负联结。显著或极显著负关联关系反映出物种间的生境需求差异性或排斥性。

图1 优势种种间关联性的 χ2检验半矩阵图Fig. 1 Semi-matrix graph of interspecific association χ2-test of dominant woody populations注:物种序号同表1,下同。
2.3 种间相关性分析
Pearson相关系数检验表明(图2),30个主要木本植物435个种对间,正相关种对数190对,占43.68%,其中极显著正相关的有66对,显著正相关的有21对,占总对数的20.00%;有245对表现负关联种,占56.32%,其中极显著负相关的有50对,显著负相关的有50对,占22.99%。正负相关比为0.78,种间关联显著率为42.99%。黄荆与刺壳花椒、米念芭与三脉叶荚蒾等66个物种对呈极显著正相关。呈显著正相关的有刺壳花椒与灰毛浆果楝、小蜡与灰毛浆果楝等21个物种对。黄荆与米念芭、米念芭与刺壳花椒等50个物种对之间呈极显著负相关。黄荆与小果蔷薇、米念芭与毛果巴豆等50个物种对表现出显著负相关。黄荆与毛果巴豆、黄荆与火棘等103个物种对为不显著正相关。表现为不显著负相关的有黄荆与小芸木、黄荆与南岭榨木等145个物种对。
Spearman秩相关系数检验中(图3),有212个物种对表现正关联,占48.74%,呈极显著正相关的物种对有73对,显著正相关对数有26对,占总对数的28.28%;221对表现负关联,占50.80%,极显著负相关种对数有88对,显著负相关对数有35对,占总对数的28.28%。有2个物种对表现不相关,正负相关比为0.96,种间关联显著率达51.03%。黄荆与刺壳花椒、米念芭与三脉叶荚蒾等组成的73个物种对表现出极显著的正相关。黄荆与抱茎菝葜、米念芭与南岭榨木等26个物种对呈显著正相关。黄荆与米念芭、米念芭与刺壳花椒等88个物种对呈极显著负相关。呈显著负相关的物种对有黄荆与红背山麻杆、刺壳花椒与瘤皮孔酸藤子等35个物种对。黄荆与刺壳花椒、黄荆与火棘等113个物种对表现出不显著的正相关。呈不显著负相关的有黄荆与毛果巴豆、黄荆与南岭榨木等98个物种对。三脉叶荚蒾与石岩枫、小果蔷薇与石岩枫呈不相关关系。

图2 主要木本植物种对间Pearson相关系数半矩阵图Fig. 2 Semi-matrix of Pearson’s correlation coefficients of dominant woody populations

图3 主要木本植物间Spearman秩相关系数半矩阵图Fig. 3 Semi-matrix of Spearman’s rank correlation coefficients of dominant woody populations
由Spearman种间相关系数星座图(图4)可以看出,黄荆、小蜡、灰毛浆果楝3个物种彼此呈极显著正相关。米念芭、三脉叶荚蒾、火棘、南岭榨木、红背山麻杆、蚬壳花椒、圆果化香树、盐肤木、小果蔷薇、瘤皮孔酸藤子、竹叶花椒、全缘火棘、豆梨等13个物种间联结性非常紧密,彼此间以显著(含极显著)正相关为主。刺壳花椒、小蜡、八角枫、抱茎菝葜、南蛇藤、龙须藤、球序鹅掌柴等7个物种间彼此主要呈显著或极显著正相关。毛果巴豆与矮小天仙果呈极显著正相关,与小芸木、斜叶榕呈显著正相关。石山柿与圆果化香树呈极显著正相关,与蚬壳花椒、矮小天仙果呈显著正相关。石山柿、小芸木、粗糠柴、石岩枫等物种相对独立,与其他物种形成的正相关关系中只有少数达到显著水平。

图4 优势种Spearman秩相关系数星座图Fig. 4 The constellation diagram of dominant species based on Spearman’s rank correlation coefficients注:物种序号同表1。
2.4 生态种组的划分
根据物种对环境的适应特点,利用RDA排序(图5),结合以上种间关联和相关分析结果,将喀斯特灌丛30个优势种分成3个生态种组。
I黄荆生态种组:包括黄荆(1)、刺壳花椒(3)、小蜡(5)、灰毛浆果楝(11)、八角枫(17)、抱茎菝葜(21)。黄荆是群落中的主要优势种,广泛分布于研究样地中,在群落结构中占据重要作用。该组物种为深根植物,对土深要求较高,组内种间关系主要呈显著或极显著正关联。
II米念芭、三脉叶荚蒾生态种组:包括米念芭(2)、三脉叶荚蒾(6)、火棘(8)、南岭榨木(10)、红背山麻杆(12)、蚬壳花椒(13)、圆果化香树(14)、盐肤木(15)、小果蔷薇(16)、瘤皮孔酸藤子(18)、竹叶花椒(19)。该组物种最多,为喜阳耐旱植物,在样地中主要分布在中上坡位。多数树种与优势种黄荆呈显著负相关。
III毛果巴豆、小芸木生态种组:包括小芸木(9)、粗糠柴(20)、紫弹树(23)、石岩枫(24)、斜叶榕(26)、矮小天仙果(27)、南蛇藤(28)、九龙藤(29)、球序鹅掌柴(30)。该组物种石生性较强,能够在裸岩率较高的生境生存。

图5 主要木本植物种与环境因子的RDA排序图Fig. 5 RDA ordination diagram of dominant shrub species and environmental variables注:物种序号见表1。Ele:海拔;Pos:坡位; Depth:土深;Rock:裸岩率;TK:全钾;TN:全氮;图中只显示达到显著性(P<0.05)的环境因子。
3 讨论
3.1 喀斯特灌丛种间联结及其成因
种间联结是植物群落重要的数量和结构特征之一,能够反映群落的稳定性[13,29],通常是由于群落生境的差异影响了物种的分布而引起的[32]。本研究中,喀斯特灌丛群落总体上呈现显著正联结,种间关联的显著率高达43.45%,表现出喀斯特地区灌丛植物种间联结性较强。这主要与群落所处的演替阶段[33]、物种间的生态位重叠与分化[34]、物种的生态习性和生境需求[13]有关。本研究对象是典型喀斯特峰丛洼地人为干扰后经封育后自然恢复的次生植被,木本植物主要以灌木为主,群落正处于次生演替中期阶段,种间关系较为复杂,有较多的物种间表现出显著的正关联和负关联[9]。随着演替进行,负关联种对将减少,群落物种关系将趋于正相关,以求得更多物种稳定共存[17,26]。
喀斯特峰丛洼地小生境存在极大的异质性[25],具体表现在小生境类型多、土层分布不连续、岩石裸露率、土壤养分含量分布上存在很大差异。不同树种对生境条件有不同偏好,生境需求一致的物种容易在同一种生境类型中伴随出现,因而种间正联结紧密;而生境需求相异的物种分布在不同小生境,负联结也变得显著。米念芭、三脉叶荚蒾、红背山麻杆等物种喜阳、耐干旱,能够在土层浅薄的生境中生长,多分布在中上坡位。而刺壳花椒、小蜡、八角枫等物种却适应于土层较厚,相对潮湿的环境,主要分布在下坡位。又如石岩枫、斜叶榕、矮小天仙果、南蛇藤、九龙藤、球序鹅掌柴等物种石生性较强,主要分布在裸岩率较高的生境中。
在喀斯特地区,岩石裸露率大,土层浅薄,部分植物夏季面临着高温干旱等胁迫。恶劣环境下,物种间的关系多向彼此有利的方向发展,以维持物种共存。有的正联结的种对可能是在群落中占据不同高度的空间,形成垂直空间上的互补性差异[15],可以充分利用不同层次的空间资源,即出现生态位的分化,避免了因资源利用造成的激烈竞争,所以能在群落中共存。如石山柿、圆果化香树、豆梨等亚乔木树种对火棘、蚬壳花椒、抱茎菝葜等灌木起到保育作用,它们之间呈现出正联结。
因此,在进行自然植被恢复过程中,可以考虑进行适当的人为调控,进行人为诱导植被恢复。例如,利用乔木树种对耐阴灌木树种的保育作用,在原乔木树种下补种耐阴灌木,以达到维持物种多样性及促进植被恢复速率的效果。同时,对于部分承受较大竞争压力的乔、灌木树种,可以适当控制周围对其起到竞争抑制作用的物种(比如,藤本植物)的密度,以促进灌丛群落向乔木群落演替的速率。
3.2 喀斯特灌丛木本植物生态组分类及指示意义
群落中生态种组由生态习性相近的物种组成。在同一生态组的物种对,具有较大的关联性和相关性,它们在适应环境、资源利用及对群落所起的功能作用常表现出相似性[15]。本研究将灌丛群落优势种划分为三个生态种组,同一生态种组的物种之间对环境的需求差异较小[35],组内物种正相关性非常显著,种间联结性强,生态种组内的物种彼此依赖,具有一定的群落依存关系[36]。不同的生态种组的物种具有不同的生活习性和生态要求[37],代表了不同的群落功能类型,如第一生态种组的黄荆、刺壳花椒、八角枫等物种,耐旱、耐贫瘠,对环境需求较低,繁殖力强,能够在群落中快速定居、扩散。第二生态种组的物种最多,彼此间联结性紧密,主要分布在中上坡位,能够适应较高pH。而第三生态种组则是一些石生性较强的物种,如石岩枫、斜叶榕等甚至能够在裸岩上生长。不同生态种组间物种联结松散,多以负相关为主,反映出它们生境需求上存在明显差异。如第一生态种组与第二生态种组的物种间主要表现为显著负相关,物种间表现出排斥性。因此,在植树造林时,可以结合生境特点和物种生态习性[13]考虑选择同一生态种组的物种进行合理搭配种植。例如,在相对干旱的中上坡位,可将米念芭、三脉叶荚蒾、火棘等树种搭配种植,在促进植被恢复的同时,又能维持较高水平的植物物种多样性。而在岩石裸露率大、土壤分布较少的生境中,可以考虑种植石生性较强的物种,以促进石漠化治理和植被恢复速率。而对于出现在特定生境下却不适宜该生境生存的物种幼小个体(终将被淘汰),可以考虑移走或换掉,以给适宜生长的物种腾出空间,促进群落稳定性。
3.3 三种检验方法的比较
本研究利用χ2检验、Pearson相关系数和Spearman秩相关系数检验等方法,对桂西北典型喀斯特灌丛群落的主要木本植物进行了种间关联和相关性检验。三种检验方法得到的结果存在一定差异:χ2检验的正负关联比为1.38,关联显著率为41.84%;Pearson相关系数检验的正负关联比为0.78,显著率为42.99%;Spearman秩相关系数检验正负关联比为0.96,关联显著率为51.03%。其原因在于,χ2检验只能根据两个物种同时存在与否的二元数据做出定性判断,而Pearson相关系数检验和Spearman秩相关系数检验是基于重要值等定量数据的检验方法,能更好地反映种间关系的本质[18]。此外,相对于Pearson相关系数检验,Spearman秩相关系数属于非参数检验,在相关性检验的使用上更灵活,具有更高的灵敏度[31]。因此在进行群落种间关联和相关性研究时,综合利用χ2检验和Pearson相关系数、Spearman秩相关系数检验,结果可信度更高,更能准确反映群落间的种间联结性[18]。
4 结论
本研究的典型喀斯特峰丛洼地灌丛主要木本植物优势种间总体正关联性显著,种间关联和相关性均有较高的关联显著率,说明群落中物种种间关系联结性较强,与群落处于演替中期有关。特殊的喀斯特生境异质性影响树种分布,进而影响群落种间关联性。根据物种对环境的适应特征,将30个喀斯特灌丛优势种划分为3个生态种组,分别为对环境要求较低、繁殖力强的黄荆生态种组;喜阳、耐干旱的米念芭、三脉叶荚蒾组;石生性较强的小芸木生态种组。在喀斯特植被恢复和植树造林的实践中,应充分考虑物种的生态习性和喀斯特生境的独特性,利用物种间的种间关系合理考虑选择同一生态种组物种进行搭配种植。
[1] Armas C, Pugnaire F I. Plant interactions govern population dynamics in a semi-arid plant community[J]. Journal of Ecology, 2005, 93(5): 978-989.
[2] 王文进, 张明, 刘福德, 等. 海南岛吊罗山热带山地雨林两个演替阶段的种间联结性[J]. 生物多样性, 2007, 15(3): 257- 263.
Wang W C, Zhang M, Liu F D, et al. Species association in tropical montane rain forest at two successional stages in Diaoluo Mountain of Hainan Island[J]. Biodiversity Science, 2007, 15(3): 257-263.
[3] 张桂萍, 张峰, 茹文明. 山西绵山植被木本植物优势种群种间关联[J]. 生态学杂志, 2006, 25(3): 295-298.
Zhang G P, Zhang F, Ru W M. Interspecific correlations among dominant populations of ligneous species in Mianshan Mountain of Shanxi[J]. Chinese Journal of Ecology, 2006, 25(3): 295-298.
[4] Connell J H. On the prevalence and relative importance of interspecific competition: Evidence from field experiments[J]. The American Naturalist, 1983,122: 661-696.
[5] Bruno J F, Stachowicz J J, Bertness M D. Inclusion of facilitation into ecological theory[J]. Trends in Ecology & Evolution, 2003, 18(3): 119-125.
[6] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004.
Zhang J T. Quantitative Ecology[M]. Beijing: Science Press, 2004.
[7] 张先平, 王孟本, 张伟锋, 等. 庞泉沟国家自然保护区森林群落木本植物种间关系的分析[J]. 植物研究, 2007, 27(3): 350-355.
Zhang X P, Wang M B, Zhang W F, et al. Inter specific relationships among woody plants of forest communities in Pangquangou National Nature Reserve at Mt. Guandi, Shanxi, China[J]. Bulletin of Botanical Research, 2007, 27(3): 350-355.
[8] Brooker R W, Maestre F T, Callaway R M, et al. Facilitation in plant communities: The past, the present, and the future[J]. Journal of Ecology, 2008, 96(1): 18-34.
[9] Suzuki R O, Numata S, Okuda T, et al. Species associations among dipterocarp species co-occurring in a Malaysian tropical rain forest[J]. Journal of Tropical Ecology, 2012, 28(3): 281-289.
[10] Rosenthal G. Selecting target species to evaluate the success of wet grassland restoration[J]. Agriculture, ecosystems & environment, 2003, 98(1): 227-246.
[11] 张金屯, 焦蓉. 关帝山神尾沟森林群落木本植物种间联结性与相关性研究[J]. 植物研究, 2003, 23(4): 458-463.
Zhang J T, Jiao R. Interspecific association between woody plants in Shenweigou of Guandi mountains, Shanxi Province[J]. Bulletin of Botanical Research, 2003, 23(4): 458-463.
[12] Arroyo A I, Pueyo Y, Saiz H, et al. Plant-plant interactions as a mechanism structuring plant diversity in a Mediterranean semiarid ecosystem[J]. Ecology and Evolution, 2015, 5(22): 5305-5317.
[13] 周刘丽, 张晴晴, 赵延涛, 等. 浙江天童枫香树群落不同垂直层次物种间的联结性与相关性[J]. 植物生态学报, 2015, 39(12): 1136-1145.
Zhou L L, Zhang Q Q, Zhao Y T, et al. Species association and correlation between vertical layers in the Liquidambar formosana community in Tiantong region, Zhejiang Province[J]. Chinese Journal of Plant Ecology, 2015, 39(12): 1136-1145.
[14] 周先叶, 王伯荪, 李鸣光, 等. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J]. 植物生态学报, 2000, 24(3): 332-339.
Zhou X Y, Wang B S, Li M G, et al. An analysis of interspecific association in secondary succession forest communities in Heishiding Natural Reserve, Guangdong Province[J]. Chinese Journal of Plant Ecology, 2000, 24(3): 332-339.
[15] 白玉宏, 毕润成, 张钦弟. 山西五鹿山自然保护区辽东栎群落木本植物种间数量关系[J]. 生态学杂志, 2012, 31(8): 1942-1948.
Bai Y H, Bi R C, Zhang Q D. Interspecific relationship of woody plants in Quercus wutaishanica community in Wulu Mountain Nature Reserve, Shanxi Province of China: A quantitative analysis[J]. Chinese Journal of Ecology, 2012, 31(8): 1942-1948.
[16] 王伟, 郭倩, 康海军, 等. 线叶嵩草草地群落构成及种间关联分析[J]. 西北植物学报, 2015, 35(10): 2096-2102.
Wang W, Guo Q, Kang H J, et al. Community composition and interspecific association analysis of Kobresia capillifolia grassland[J]. Acta Botanica Boreali-Occidentalia Sinica, 2015, 35(10): 2096-2102.
[17] Bertness M D, Hacker S D. Physical stress and positive associations among marsh plants[J]. American Naturalist, 1994, 144(3): 363-372.
[18] Cheng D L, Wang G X, Chen B M, et al. Positive interactions: Crucial organizers in a plant community[J]. Journal of Integrative Plant Biology, 2006, 48(2): 128-136.
[19] Esquivias M P, Zunzunegui M, Barradas M C D, et al. Competitive effect of a native-invasive species on a threatened shrub in a Mediterranean dune system[J]. Oecologia, 2015, 177(1): 133-146. [20] Suzuki R O, Numata S, Okuda T, et al. Growth strategies differentiate the spatial patterns of 11 dipterocarp species coexisting in a Malaysian tropical rain forest[J]. Journal of Plant Research, 2009, 122(1): 81-93.
[21] Su S, Liu J, He Z, et al. Ecological species groups and interspecific association of dominant tree species in Daiyun Mountain National Nature Reserve[J]. Journal of Mountain Science, 2015, 12(3): 637-646.
[22] 张岗岗, 王得祥, 张明霞, 等. 秦岭南坡松栎林群落演替过程中种间联结性和相关性研究[J]. 西北植物学报, 2015, 35(8): 1657-1668.
Zhang G G, Wang D X, Zhang M X, et al. Interspecific association and correlation of main species in the succession process of Pine Oak forest community on the south-facing slopes in Qinling mountain[J]. Acta Botanica Boreali-Occidentalia Sinica, 2015, 35(8): 1657-1668.
[23] 张忠华, 胡刚, 倪健. 茂兰喀斯特森林群落的种间分离特征[J].生态学报, 2010, 30 (9): 2235-2245.
Zhang Z H, Hu G, Ni J. Interspecific segregation of old-growth Karst forests in Maolan, Southwest China[J]. Acta Ecologica Sinica, 2010 , 30 (9): 2235-2245.
[24] He X Y, Wang K L, Zhang W, et al. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst[J]. Plant and Soil, 2008, 307(1/2): 123-134.
[25] 韩美荣, 彭晚霞, 曾馥平, 等. 喀斯特峰丛洼地次生林主要土壤矿质成分空间异质性研究[J]. 农业现代化研究, 2011, 32(6): 748-751.
Han M R, Peng W X, Zeng F P, et al. Spatial heterogeneity of soil mineral composition in secondary forest in karst cluster-peakdepression region[J]. Research of Agricultural Modernization, 2011, 32(6): 748-751.
[26] 鲍士旦.土壤农化分析[M]. 北京:中国农业出版社, 2000.
Bao S D, Soil and Agriculture Chemistry Analysis[M]. Beijing: China Agriculture Press, 2000.
[27] Schluter D. A variance test for detecting species associations, with some example applications[J]. Ecology, 1984, 65(3): 998-1005.
[28] 杨晓东, 傅德平, 袁月, 等. 新疆艾比湖湿地自然保护区主要植物的种间关系[J]. 干旱区研究, 2010, 27(2): 249-256.
Yang X D, Fu D P, Yuan Y, et al. Analysis on interspecific relationship of the dominant plant species in the Ebinur Lake Wetland Nature Reserve, Xinjiang[J]. Arid Zone Research, 2010, 27(2): 249-256.
[29] 王乃江, 习世红, 周秦生, 等. 子午岭桥山林区柴松群落主要种群种间联结性研究[J]. 西北植物学报, 2010 (4): 795-805.
Wang N J, Xi S H, Zhou Q S, et al. Interspecific association of dominant populations of Pinus tabulaef ormisf. shekanensis Communities in Qiaoshan forest area[J]. Acta Botanica Boreali-Occidentalia Sinica, 2010 (4): 795-805.
[30] Dice L R. Measures of the amount of ecologic association between species[J]. Ecology, 1945, 26(3): 297-302.
[31] 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究——Ⅰ, 种间联结测式的探讨与修正[J]. 植物生态学报, 1985, 9(4): 274-285.
Wang B S, Peng S L. Interspecific association of lower-subtropical evergreen-broadleaved forestsⅠ. The exploration and the revision on the measuring formulas of interspecific association[J]. Chinese Journal of Plant Ecology, 1985, 9(4): 274-285.
[32] 杜荣赛. 生物统计学[M]. 北京: 高等教育出版社, 1999.
Du R S. Biostatistics[M]. Beijing: Higher Education Press, 1999.
[33] Kershaw K A, Looney J H H. Quantitative and Dynamic Plant Ecology[M]. London: Edward Arnold Limited, 1985.
[34] Su S, Liu J, He Z, et al. Ecological species groups and interspecific association of dominant tree species in Daiyun Mountain National Nature Reserve[J]. Journal of Mountain Science, 2015, 12(3): 637-646.
[35] 张震云, 周伶, 上官铁梁. 半干旱区锦鸡儿群落优势种种间关系[J]. 生态学杂志, 2011, 30(9): 1868-1874.
Zhang Z Y, Zhou L, Shangguan T L. Interspecific relationships of dominant species in Caragana communities in semi-arid regions of China[J]. Chinese Journal of Ecology, 2011, 30(9): 1868-1874.
[36] 钟宇, 张健, 杨万勤, 等. 巨桉人工林草本层优势种的种间关系及生态种组的划分[J]. 草业学报, 2010, 19(3): 56-62.
Zhong Y, Zhang J, Yang W Q, et al. Interspecific relationships and ecological species groups of dominant herbage species in Eucaly ptus grandis plantation[J]. Acta Prataculturae Sinica, 2010, 19(3): 56-62.
(责任编辑:王育花)
The interspecific associations between dominant species and their environmental interpretation under a typical shrub community in karst peak-cluster depression regions of Northwest Guangxi
ZHENG Sheng-meng1,2,3, GAI Shuang-shuang1,2,3, XIE Qiang1, SU Yi-rong2,3, CHEN Xiang-bi2,3, HU Ya-jun2,3, HE Xun-yang2,3*
(1. College of Life Science, Guangxi Normal University, Guilin, Guangxi 541004, China; 2. Key Laboratory of Agro-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha, Hunan 410125, China; 3. Huanjiang Observation and Research Station for Karst Ecosystems, Chinese Academy of Sciences, Huanjiang, Guangxi 547100, China)
Study on the community structure characteristics and dynamics of typical shrub community is important to understand the coexistence mechanism of shrub species in the karsts area. In this study, the interspecific associations among 435 species pairs of the 30 dominant woody species of shrub community were analyzed in typical karst peakcluster depression in Northwest of Guangxi Province. Variance analysis showed that the interspecific correlations of the shrub community was significantly positive. The χ2test showed that 249 pairs were positive and 179 pairs were negative correlationship. The correlation ratio was 1.38 among the 435 species pairs, indicating that the associations among species were relatively strong. The Spearman’s rank correlation coefficient tests had higher sensitivity than Pearson’s correlation coefficient test in describing the species associations, suggesting that results of the Spearman’s rank correlation coefficient tests had more species pairs to reach the significant level among the 435 species pairs than did Pearson’s correlation coefficient test. According to the species adaptability to the environment, the leading ecological factors and redundancy analysis (RDA) sort, the 30 dominant species were divided into three ecological species groups: the first ecological species group was represented by Vitex negundo, the second ecological species group by Tirpitzia ovoidea and Viburnum triplinerve, and the third ecological species group by Micromelum integerrimum. In the practice of the vegetation restoration and afforestation in karst region, it is necessary to fully consider the characteristic ofecological species habitat and the uniqueness of the karst environment, and to choose the same ecological species group for proper plantation on the basis of interspecific relationship.
Karst; shrub community; dominant species; interspecific association; ecological species group
HE Xun-yang, E-mail: hbhpjhn@isa.ac.cn.
10.13872/j.1000-0275.2016.0126
S181.1
A
1000-0275(2016)06-1198-08
中国科学院科技服务网络计划(KFJ-EW-STS-092);国家自然科学基金项目(31270551)。
郑生猛(1989-),男,湖南宁远人,硕士研究生,主要从事植物与环境生态方面的研究,E-mail: 297336605@qq.com;通讯作者:
何寻阳(1972-),男,湖南平江人,博士,副研究员,主要从事土壤微生物生态方面的研究,E-mail: hbhpjhn@isa.ac.cn。
2016-04-22,接受日期:2016-10-08
Foundation item: Science and Technology Service Network Initiative (KFJ-EW-STS-092); National Natural Science Foundation of China (31270551).
Received 22 April 2016;Accepted 8 October, 2016
猜你喜欢
水土保持学报(2022年5期)2022-10-10
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
乡村地理(2019年2期)2019-11-16
草业科学(2019年5期)2019-06-04
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
蔬菜(2018年5期)2018-05-17
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
杂草学报(2015年2期)2016-01-04
文化月刊·下旬刊(2014年6期)2014-08-28
