
干旱胁迫下外源甜菜碱对石榴光合作用、渗透调节及保护酶活性的影响
2017-03-21 08:14范春丽罗青
江苏农业科学 2016年11期
范春丽+罗青
摘要:选用2年生泰山红石榴为试验材料,用200 mmol/L浓度甜菜碱叶面喷施,研究干旱胁迫下喷施甜菜碱对石榴光合作用、渗透调节及抗氧化酶响应的影响。结果表明,干旱胁迫下,喷施甜菜碱提高了石榴叶片可溶性糖、游离脯氨酸含量,同时提高苹果幼树渗透调节能力;提高了超氧化物歧化酶(SOD)、过氧化物酶(CAT)、抗坏血酸过氧化物酶(APX)抗氧化酶活性,降低了丙二醛(MDA)、H2O2积累,缓解了干旱胁迫对细胞结构的伤害;同时,提高叶片相对含水量、叶绿素含量,使叶片光合能力增强。甜菜碱提高苹果幼树抗旱性的主要作用机制,是对渗透调节物质代谢的调节及对抗氧化酶等生物大分子物质结构的保护。
关键词:外源甜菜碱;干旱;石榴;光合作用;渗透调节;保护酶
中图分类号: S665.401 文献标志码: A
文章编号:1002-1302(2016)11-0229-04
近年来,河南省降水量呈减少趋势,年降水分布不均匀,干旱对果树生产影响越来越严重[1]。干旱胁迫打破叶片水分平衡,造成水分代谢碳代谢失衡,影响植物一系列代谢活动[2],相关研究表明,干旱胁迫导致气孔导度(Gs)降低,净光合速率(Pn)下降[3]。长时间的干旱胁迫导致保护酶失活, O-2·、丙二醛(MDA)等积累,损伤细胞结构,发生光破坏[4-6]。
长期的进化使植物形成一整套的生理生化机制帮助其抵御不良的外界环境。其中,渗透调节、抗氧化酶是植物抵抗干旱环境的重要手段[7]。相关研究表明,抗旱能力较强的品种拥有较高的渗透调节能力及抗氧化酶活性, O-2· 、H2O2等有害物质积累较少,同时光合能力较高[8-10]。据报道,施用某些外源物质是增强植物抗旱能力的有效手段,如施用钙、壳聚糖、硅等[11-13]。甜菜碱是一种季铵类化合物,是重要的细胞相溶性物质,干旱胁迫下在许多植物细胞内大量积累,其生理功能主要是参与细胞渗透调节,参与稳定生物大分子的结构与功能,影响离子在细胞内的分布和吸收等[14]。施用甜菜碱能够有效提高作物抗旱性,相关研究在小麦、玉米、烟草等上有较多的报道[15-17]。
石榴是果树开发的重要树种,目前关于石榴的研究主要集中在育种、果实组分及果色形成机制等方面,对于抗逆性的研究较少,尤其关于甜菜碱调节石榴抗旱性方面研究更是鲜见报道[18-19]。本研究以2年生石榴幼树为试验材料,研究干旱胁迫下施用外源甜菜碱,石榴幼树光合作用、渗透调节、酶促调节复合响应,以期为提高石榴抗旱能力提供技术支持。
1 材料与方法[LL]
1.1 材料
试验材料为泰山红石榴(Punica granatum L. ‘Taishanhong),长势一致,2年生,取自河南省河阴石榴基地。2014年3月中旬栽植于盆口直径40 cm、高35 cm的塑料花盆中,挖20 cm土穴,将盆埋入,防止高温对根系的影响。基质配比为园土 ∶有机肥 ∶细沙=3 ∶1 ∶1,每盆基质15 kg,田间持水量21.5%,有机质含量14.3 g/kg,全氮含量2.2 g/kg,速效磷含量 39.8 mg/kg,速效钾含量118.4 mg/kg。置于避雨棚内,每株树留取相同枝量(5根枝条),自然条件下生长。
1.2 试验设计
2015年5月下旬进行试验处理,选取20株于傍晚浇透水后停止浇水,进行干旱处理,第2天(土壤相对含水量85%~75%)进行第1次采样,用烘干称质量法测土壤相对含水量,并进行光合测定,此后每隔1 d用烘干称质量法测量土壤相对含水量,在土壤相对含水量达到60%~50%、45%~35%、30%~20%时进行取样并进行光合测定。试验设2个处理:(1)处理组,选取待干旱处理的10株树,干旱处理前叶面喷施200 mmol/L甜菜碱(本研究采用甘氨酸甜菜碱,简称GB经预试验200 mmol/L效果最好),连续喷施5 d,至开始干旱处理结束,08:00、20:00定时2遍喷施;(2)对照(CK),选取待干旱处理的10株树喷施去离子水,喷施方法同处理组。取样时选取枝条中部功能叶片采样。
1.3 测定项目与方法
1.3.1 光合作用测定 取营养枝,以生长点为起始点选取第6~8节位功能叶,作为待测叶片,重复15次。利用TPS-2光合仪(Hansatech,英国),晴天09:00—11:00测定光合参数,包括光合速率、气孔导度、细胞间隙二氧化碳浓度(Ci)等。
1.3.2 叶片相对含水量(RWC)、叶绿素含量(Chl)测定 叶片相对含水量采用称质量法[20]测定;叶绿素含量测定采用95%乙醇浸泡法[21]。
1.3.3 保护酶活性测定 超氧化物歧化酶(SOD)活性采用NBT法[22]测定;过氧化氢酶(CAT)活性测定采用紫外吸收法[22];抗坏血酸过氧化物酶(APX)活性采用赵世杰等方法[22]测定。
1.3.4 丙二醛(MDA)、H2O2含量测定 MDA采用硫代巴比妥酸法[22]测定;H2O2含量测定采用可见光法试剂盒(南京建成生物工程研究所)。
1.3.5 渗透调节物质含量测定 可溶性糖含量采用蒽酮比色法[22]测定;游离脯氨酸含量采用茚三酮比色法[22]测定。
1.4 数据处理
采用SPSS 17.0软件对数据进行统计分析,采用单因素方差分析(One-Way ANOVA)和最小显著差异法(LSD)比较不同数据间的差异。利用Excel 2003软件作图。
2 结果与分析
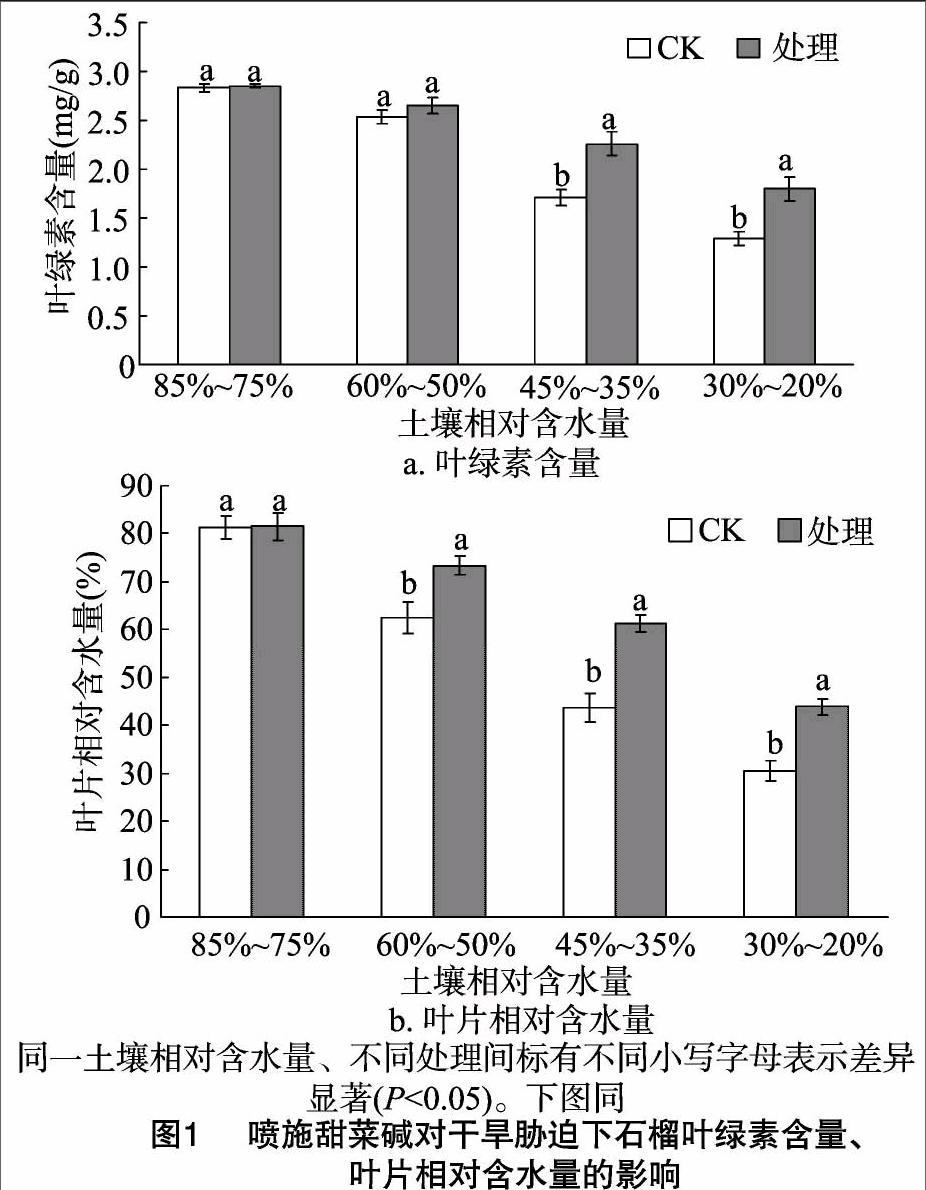
2.1 外源甜菜碱对干旱条件下石榴叶片相对含水量及叶绿素含量的影响
从图1可见,干旱胁迫下,CK与处理石榴叶片相对含水量、叶绿素含量随胁迫加重呈降低趋势。但是土壤相对含水量低于45%以后甜菜碱处理组的叶片叶绿素含量显著高于CK,而土壤相对含水量60%~50%时甜菜碱处理组叶片相对含水量就显著高于CK。土壤相对含水量为45%~35%、30%~20% 时,处理组叶片相对含水量和叶绿素含量比CK分别提高40.60%、44.24%,32.32%、39.84%。表明干旱条件下施用甜菜碱能有效提高石榴叶片相對含水量、叶绿素含量。
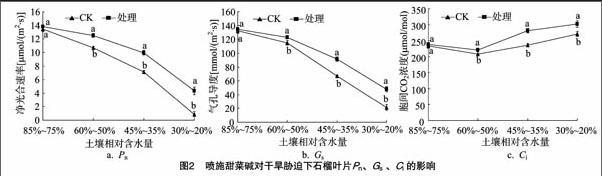
2.2 外源甜菜碱对干旱条件下石榴叶片光合能力、光合参数的影响
从图2可以看出,干旱对石榴叶片光合能力影响较大,轻度的干旱即可引起叶片光合功能大幅度下降。经甜菜碱处理,叶片Pn下降速度减缓,在3种干旱程度下与CK相比较差异显著,Pn分别提高17.19%、40.38%、399.40%。喷施甜菜碱能够有效地提高干旱胁迫下石榴叶片的光合能力。Gs变化趋势与Pn一致,呈下降趋势,Ci先降低后升高,光合能力下降的影响因子由气孔因素过渡到非气孔因素,在3种胁迫程度下,处理与CK差异显著。
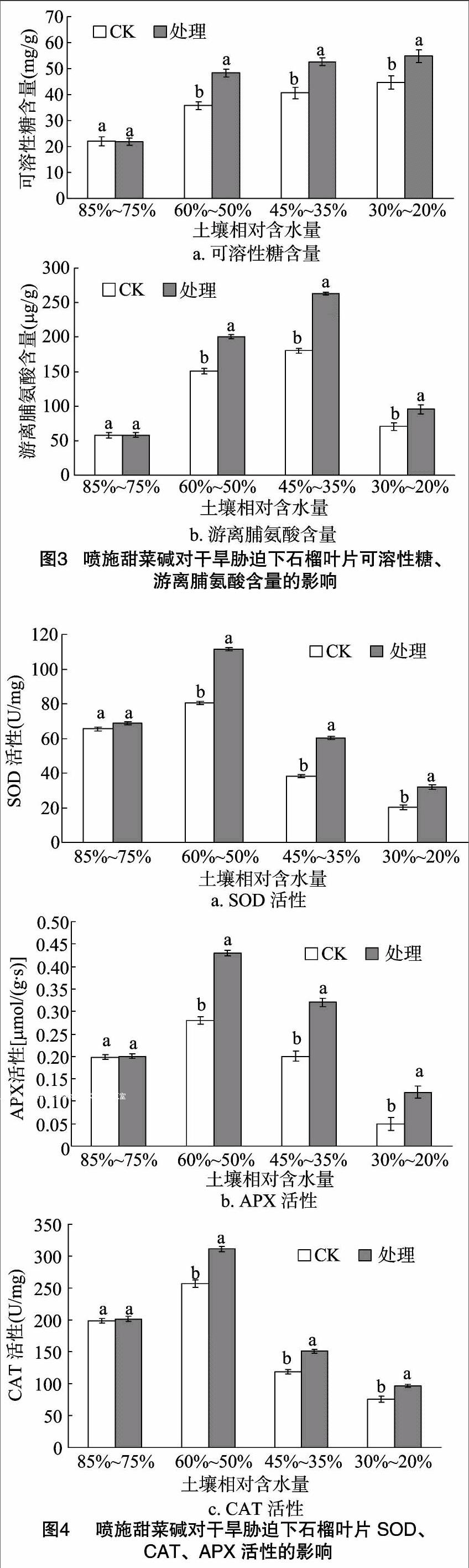
2.3 外源甜菜碱对干旱条件下石榴叶片渗透调节的影响
从图3可以看出,随着干旱程度加重,可溶性糖含量呈升高趋势,游离脯氨酸含量呈先升高后下降的趋势。表明干旱胁迫诱导石榴启动渗透调节,可溶性糖含量等渗透调节物质增加,以抵御干旱环境。在3种干旱程度下,甜菜碱处理组可溶性糖含量、游离脯氨酸含量显著高于CK。在土壤相对含水量分别为60%~50%、45%~35%、30%~20%时,处理叶片可溶性糖含量分别提高34.92%、29.56%、22.87%,游离脯氨酸含量分别提高33.27%、46.11%、35.42%。表明干旱胁迫下喷施甜菜碱能够调节石榴叶片渗透调节物质代谢,促进可溶性糖、游离脯氨酸积累,提高了石榴的抗旱能力。
2.4 外源甜菜碱对干旱条件下石榴叶片保护酶活性的影响
从图4可以看出,3种保护酶(SOD、CAT、APX)活性变化规律一致,随干旱程度加重,呈先升高后下降的趋势。石榴接收环境干旱信号,在土壤相对含水量60%~50%时,3种抗氧化酶活性达到最大,随后干旱加重,3种酶活性降低。甜菜碱处理组3种抗氧化酶活性表现出同CK相同的趋势,但是在相同干旱程度下,甜菜碱处理组3种酶活性显著高于CK,说明干旱胁迫下外源甜菜碱能够维持抗氧化酶较高的活性,提高石榴抗旱能力。
2.5 外源甜菜碱对干旱条件下石榴叶片丙二醛及过氧化氢活性的影响
干旱胁迫诱导细胞活性氧代谢,H2O2作为抗氧化酶清除活性氧自由基的產物逐渐积累,活性氧自由基对细胞结构产生伤害,其中MDA是衡量膜损伤的指标。从图5可以看出,随着干旱程度加重,叶片中MDA、H2O2含量随着干旱程度加重逐渐升高,表明细胞结构受损愈加严重。与CK比较,土壤相对含水量在60%~20%条件下,甜菜碱处理组MDA、H2O2含量显著降低,在干旱程度最严重时,甜菜碱处理组MDA、H2O2含量分别比CK降低27.94%、20.28%。表明干旱胁迫条件下喷施甜菜碱可有效减少有害物质积累,缓解膜损伤。
3 讨论
干旱胁迫下,植物根系吸收水分能力降低,导致地上部叶片相对含水量下降,叶片失水,维持气孔开张的膨压降低,导致气孔导度下降,植物光合能力下降,此时气孔限制是影响光合能力下降的主要因素[23-24]。随着干旱胁迫加重,ROS积累,产生氧化胁迫,ROS不仅导致光合膜结构损伤并且与叶绿素降解有关[25-26],从而影响光合作用光反应,使植物光合能力下降,此时非气孔限制是引起光合能力下降的主要因素。本研究中根据Pn、Gs、Ci的变化[27],土壤相对含水量50%以下,非气孔限制是光合能力下降的主要因素,干旱导致叶绿素含量下降,MDA、H2O2含量升高,膜结构损伤严重。
据相关报道,甜菜碱对蛋白和色素具有保护作用[28],甜菜碱处理的小麦单位叶面积的色素含量显著高于未施用外源甜菜碱的小麦幼苗[29];侯彩霞等研究表明,外源甜菜碱能够维持D1蛋白等大分子结构稳定[30];赵新西等报道,甜菜碱能够减轻胁迫对类囊体膜的损伤[15];黄义春等报道,外源甜菜碱能显著提高干旱胁迫下玉米SOD等抗氧化酶活性[16]。本研究也发现,甜菜碱处理显著提高了石榴叶片SOD、CAT、APX抗氧化酶活性,MDA含量降低,膜损伤减轻,H2O2积累减少。甜菜碱对酶蛋白的保护作用,使抗氧化酶保持在较高的活性,及时清除干旱产生的ROS,缓解了ROS对细胞结构的氧化损伤,另一方面ROS可能参与Chl 4-吡咯环的破坏[26],ROS的减少,可能是叶绿素含量增加的原因之一。
干旱胁迫下,绝大多数植物启动渗透调节以抵御干旱环境[31-32]。植物的渗透调节是通过在细胞中合成积累具有渗透活性而又无毒的有机物来实现的,这些有机物可以促进细胞内水分维持和从外部吸收水分[33]。本试验结果表明,喷施甜菜碱能够促进渗透调节物质可溶性糖、游离脯氨酸的积累,提高了树体吸水能力,叶片相对含水量显著增加,改善了叶片水分状况,为光化学反应和电子传递提供适当的生理环境,这可能是喷施甜菜碱提高光合效率的原因之一。
综上所述,干旱胁迫下喷施甜菜碱能够减少细胞膜结构的损伤,提高叶片叶绿素含量,缓解干旱对光合电子传递链抑制,从而维持较高的光合能力,提高石榴抗旱性。甜菜碱调节渗透物质代谢,促进渗透调节,改善了树体水分状况对生物大分子的保护作用,可能是干旱胁迫下甜菜碱处理使叶片维持较高光合能力、提高树体抗旱性的主要原因。
参考文献:
[1]韩 玮,韩永红,杨沈斌. 1961—2011年山东气候资源及气候生产力时空变化特征[J]. 地理科学进展,2013,32(3):425-434.
[2]董 蕾,李吉跃. 植物干旱胁迫下水分代谢、碳饥饿与死亡机理[J]. 生态学报,2013,33(18):5477-5483.
[3]温国胜,张国盛,吉川贤. 干旱胁迫对臭柏水分特性的影响[J]. 林业科学,2004,40(5):84-87.
[4]原向阳,郭平毅,张丽光,等.干旱胁迫下草甘膦对抗草甘膦大豆幼苗保护酶活性及脂质过氧化作用的影响[J]. 中国农业科学,2010,43(3):698-705.
[5]许长成,赵世杰,樊继莲,等. 干旱胁迫下大豆与玉米叶片光破坏的防御[J]. 植物生理学报,1998,24(1):17-23.
[6]Zhao C M,Wang G X. Effects of drought stress on the photoprotection in Ammopiptanthus mongolicus leaves[J]. Acta Botanica Sinica,2002,44(11):1309-1313.
[7]Wei L M,Jia L R,Hu X A. Advances in studies on the physiology and biochemistry of maize drought resistance[J]. Agricultural Research in the Arid Areas,1997,15(4):66-71.
[8]李德全,邹 琦,程炳嵩. 土壤干旱下不同抗旱性小麦品种的渗透调节和渗透调节物质[J]. 植物生理学报,1992,18(1):37-44.
[9]Dong L,Li J Y,Wang J H,et al. Effect of drought stress on osmotic regulation substances of five Catalpa bungei clones[J]. Agricultural Science Technology,2013,14(9):1335-1343.
[10]Ramachandra R A,Chaitanya K V,Vivekanandan M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants[J]. Journal of Plant Physiology,2004,161(11):1189-1202.
[11]胡景江,張林森,刘建朝. 干旱及外源低聚壳聚糖对1年生苹果幼树渗透调节能力的影响[J]. 林业科学,2012,48(9):15-21.
[12]李清芳,马成仓,尚启亮. 干旱胁迫下硅对玉米光合作用和保护酶的影响[J]. 应用生态学报,2007,18(3):531-536.
[13]张会慧,张秀丽,许 楠,等. 外源钙对干旱胁迫下烤烟幼苗光系统Ⅱ功能的影响[J]. 应用生态学报,2011,22(5):1195-1200.
[14]张立新,李生秀. 甜菜碱与植物抗旱/盐性研究进展[J]. 西北植物学报,2004,24(9):1765-1771.
[15]赵新西,马千全,杨兴洪,等. 根施甜菜碱对干旱胁迫下小麦幼苗类囊体膜组分和功能的影响[J]. 植物生理与分子生物学学报,2005,31(2):135-142.
[16]黄义春,李建民,段留生,等. 甜菜碱对玉米幼苗抗旱性的诱导效应[J]. 玉米科学,2011,19(1):95-100.
[17]马新蕾,王玉军,谢胜利,等. 根施甜菜碱对水分胁迫下烟草幼苗光合机构的保护[J]. 植物生理与分子生物学学报,2006,32(4):465-472.
[18]Lansky E P,Newman R A,Punica G. And its potential for prevention and treatment of inflammation and cancer[J]. Journal of Ethnopharmacology,2007,109(3):177-206.
[19]Aviram M,Volkova N,Coleman R,et al. Pomegranate phenolics from the peels,arils,and flowers are antiatherogenic:studies in vivo in atherosclerotic apolipoprotein e-deficient(E0) mice and in vitro in cultured macrophages and lipoproteins[J]. Journal of Agricultural and Food Chemistry,2008,56(3):1148-1157.
[20]Huang B R,Gao H W. Physiological response of diverse tall fescue cultivars to drought stress[J]. Hort Science,1999,34(5):897-901.
[21]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:134-138.[HJ1.75mm]
[22]赵世杰,史国安,董新纯. 植物生理学实验指导[M]. 北京:中国农业科学技术出版社,1998.
[23]Flexas J,Bota J,Galmés J,et al. Keeping a positive carbon balance under adverse conditions:responses of photosynthesis and respiration to water stress[J]. Physiologia Plantarum,2006,127(3):343-352.
[24]Gallé A,Haldimann P,Feller U. Photosynthetic performance and water relations in young pubescent oak(Quercus pubescens) trees during drought stress and recovery[J]. New Phytologist,2007,174(4):799-810.
[25]刘 强,王占武,周晓梅. 两种枸杞对NaCHO3胁迫的抗性生理响应[J]. 南京林业大学学报,2014,38(6):165-169.
[26]Jiangm Y,Yang W Y,Xu J. Active Oxygen damage effect of chlorophyll degradation in rice seedlings under osmotic stress[J]. Acta Botanica Sinica,1994,36:289-295.
[27]李潮海,赵亚丽,杨国航,等. 遮光对不同基因型玉米光合特性的影响[J]. 应用生态学报,2007,18(6) :1259-1264.
[28]Makela P,Karkkainen J,Somersalo S. Effects of glycinebetaine on chloroplast ultra structure,chlorophyll and proteincontent,and RuBPCO activities in tomato grown underdrought or salinity[J]. Biologia Plantarum,2000,43(3):471-475.
[29]郭启芳,马千全,孙 灿,等. 外源甜菜碱提高小麦幼苗抗盐性的研究[J]. 西北植物学报,2004,24(9):1680-1686.
[30]侯彩霞,徐春和,汤章城. 甜菜碱对PSⅡ放氧中心结构的选择性保护[J]. 科学通报,1997,42(17):1857-1859.
[31]熊搖洁,邹晓芬,邹小云,等. 干旱胁迫对不同基因型油菜农艺性状和产量的影响[J]. 江苏农业学报,2015,31(3):494-499.
[32]靳摇容,张爱君,史新敏,等. 干旱胁迫下钾对甘薯幼苗光合特性及根系活力的影响[J]. 江苏农业学报,2014,30(5):992-996.
[33]Friedman R,Altman A. The effect of salt stress on polyamine biosynthesis and content in mung bean plants and in halophytes[J]. Physiologia Plantarum,1989,76(76):295-302.
猜你喜欢
科学(2022年4期)2022-10-25
音乐天地(音乐创作版)(2021年7期)2021-10-13
草原歌声(2021年1期)2021-07-16
少先队活动(2021年1期)2021-03-29
现代农业科技(2017年1期)2017-03-06
现代农业科技(2016年21期)2017-03-06
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
