
Evidence of cortical reorganization of language networks after stroke with subacute Broca’s aphasia: a blood oxygenation level dependent-functional magnetic resonance imaging study
2017-03-30 04:44WeihongQiuHuixiangWuQingluYangZhuangKangZhaocongChenKuiLiGuorongQiuChunqingXieGuifangWanShaoqiongChen
中国神经再生研究(英文版) 2017年1期
Wei-hong Qiu, Hui-xiang Wu Qing-lu Yang Zhuang Kang, Zhao-cong Chen Kui Li, Guo-rong Qiu Chun-qing Xie Gui-fang Wan Shao-qiong Chen
1 Department of Rehabilitation Medicine, theird A ffiliated Hospital of Sun Yat‐sen University, Guangzhou, Guangdong Province, China
2 Department of Radiology, theird A ffiliated Hospital of Sun Yat‐sen University, Guangzhou, Guangdong Province, China
Evidence of cortical reorganization of language networks after stroke with subacute Broca’s aphasia: a blood oxygenation level dependent-functional magnetic resonance imaging study
Wei-hong Qiu1,#,*, Hui-xiang Wu1,#, Qing-lu Yang1,#, Zhuang Kang2, Zhao-cong Chen1, Kui Li1,*, Guo-rong Qiu1, Chun-qing Xie1, Gui-fang Wan1, Shao-qiong Chen2
1 Department of Rehabilitation Medicine, theird A ffiliated Hospital of Sun Yat‐sen University, Guangzhou, Guangdong Province, China
2 Department of Radiology, theird A ffiliated Hospital of Sun Yat‐sen University, Guangzhou, Guangdong Province, China
How to cite this article:Qiu WH, Wu HX, Yang QL, Kang Z, Chen ZC, Li K, Qiu GR, Xie CQ, Wan GF, Chen SQ (2017) Evidence of cortical reorganization of language networks aer stroke with subacute Broca’s aphasia: a blood oxygenation level dependent-functional magnetic resonance imaging study. Neural Regen Res 12(1):109-117.
Open access statement:is is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
Graphical Abstract

Bilateral hemispheres’ cortex activation of post-stroke Broca’s aphasic patients
#
orcid: 0000-0002-0020-8659 (Wei-hong Qiu) 0000-0002-2642-5213 (Kui Li)
Aphasia is an acquired language disorder that is a common consequence of stroke.e pathogenesis of the disease is not fully understood, and as a result, current treatment options are not satisfactory. Here, we used blood oxygenation level‐dependent functional magnetic reso‐nance imaging to evaluate the activation of bilateral cortices in patients with Broca’s aphasia 1 to 3 months aer stroke. Our results showed that language expression was associated with multiple brain regions in which the right hemisphere participated in the generation of lan‐guage.e activation areas in the lehemisphere of aphasia patients were signi fi cantly smaller compared with those in healthy adults.e activation frequency, volumes, and intensity in the regions related to language, such as the leinferior frontal gyrus (Broca’s area), the lesuperior temporal gyrus, and the right inferior frontal gyrus (the mirror region of Broca’s area), were lower in patients compared with healthy adults. In contrast, activation in the right superior temporal gyrus, the bilateral superior parietal lobule, and the leinferior temporal gyrus was stronger in patients compared with healthy controls.ese results suggest that the right inferior frontal gyrus plays a role in the recovery of language function in the subacute stage of stroke‐related aphasia by increasing the engagement of related brain areas.
nerve regeneration; functional magnetic resonance imaging; cortical functional connectivity; language regions; neuroplasticity; Perisylvian language regions; brain activation; right hemisphere; picture-naming task; neural regeneration
Introduction
Language, which is considered the most distinct function of high‐level cognition in human beings, is controlled by an intricate system involving many brain regions across both hemispheres. In the 95% of humans, the lehemisphere is the main domain implicated in language function (Sza fl arski et al., 2011). Wernicke and Broca’s areas in the left hemi‐sphere are predominant components of language perception and speech production processes (Aminoff and Daroff, 2014). From the 1980s onwards, studies have produced ev‐idence implicating the right hemisphere in language. An important demonstration of this is the ‘Wada test’ (a unilat‐eral amobarbital injection) in chronic aphasia patients, and showed the right hemishere engaged in the language process (Krashen, 1973; Rapport et al., 1983). Additionally, aphasia patients with a lehemisphere lesion may exhibit improved language function, while those who suffer a stroke in the right hemisphere may lose all language ability (Cambier et al., 1983).
Aphasia—the acquired loss of language ability—is one of the most common and debilitating cognitive consequences of stroke, a ff ecting approximately 20—40% of stroke survivors and impacting approximately one million individuals in the US alone (Siirtola et al., 1977; Pedersen et al., 1995; Berthier, 2005).e stroke incidence in China is about 205—584 per ten thousand, and includes one third of patients with speech disorders (Tsai et al., 2013).e incidence of aphasia in the elderly population is increasing dramatically in China (Tsai et al., 2013), and motor aphasia is one of the most common types of post‐stroke aphasia. Patients with Broca’s aphasia usually retain greater comprehensionvs. expressive capacity, which decreases their ability to communicate with others (Hamilton, 2016).is, along with a high degree of depen‐dence in daily life can lead to loneliness, passiveness, and de‐pression (Hamilton, 2016), as well as further serious psycho‐logical problems.us, aphasia represents a heavy burden to families and society.
Functional magnetic resonance imaging (fMRI) and pos‐itron emission tomography (PET) have been widely used to explore brain functional networks (Bottini et al., 1994; Jang and Jang, 2016; Park and Park, 2016). For instance, these tools have been used to explore the role of the right hemi‐sphere in language in healthy and aphasia groups (Bottini et al., 1994). Data from follow‐up studies of aphasia patients suggest that dynamic changes can occur in the bilateral hemispheres, such as spontaneous recovery of language (Müller et al., 1999; Saur et al., 2006; Winhuisen et al., 2007), as well as functional changes in the brain following language therapy at specific recovery stages (Rochon et al., 2010; Sza fl arski et al., 2011; Mattioli et al., 2014).ese data high‐light the involvement of the right hemisphere in post‐stroke language function. However, evidence from neuroimaging studies has been somewhat inconsistent (Finger et al., 2003; Hillis, 2007), especially with regard to (1) aphasia type, (2) fMRI task, and (3) changes during aphasia recovery. With regard to aphasia type, Thomas et al. (1997) showed that the activation pattern in the bilateral hemisphere in indi‐ viduals with Broca’s aphasia tended to shitowards the lehemisphere a few months after onset, while patients with Wernicke’s aphasia did not show this tendency. With regard to fMRI task, Hamilton et al. (2011) investigated the rela‐tionship between bilateral hemisphere activation in aphasia patients during language tasks by calculating 240 activation foci from 104 aphasia patients and 197 foci from 129 con‐trols using fMRI and PET. They found that activation in the right inferior frontal gyrus was reliably associated with language production tasks, while that in the right middle temporal gyrus was associated with comprehension tasks. Finally, some studies have revealed inconstancies in the de‐velopment from the acute to chronic phase of aphasia. Saur et al. (2006) used fMRI with a parallel auditory comprehen‐sion task to assess changes as participants progressed from the acute to the chronic stage of aphasia. In the acute phase (mean: 1.8 days), they observed weak activation in the per‐ilesional structures in the left hemisphere. In the subacute phase (mean: 12.1 days), they found that activation of the bilateral hemispheres corresponded with the peak values in the right Broca‐homologues. In the chronic phase (mean: 321 days), shifting of the activation peak back to the left hemisphere was associated with further language improve‐ment. A recent review by Anglade et al. (2014) described neuroplastic reorganization in the bilateral hemispheres of aphasia patients in different stages, suggesting that ac‐tivation in the leand right hemispheres varies with time from onset. Non‐invasive stimulation techniques such as repetitive transcranial magnetic stimulation (rTMS) and transcranial direct current stimulation (tDCS) have enabled further exploration of the mechanisms underlying recovery in the left and right hemispheres. Low‐frequency rTMS has been used to inhibit the right‐hemisphere homotopic language area in a patient with chronic aphasia, improving language ability (Chie ff o et al., 2014). Additionally, a study using longitudinal rTMS combined with PET showed that language‐related brain areas in the left hemisphere are essential during aphasia recovery; the right hemisphere is also activated, but it is not critical to promote language improvement (Winhuisen et al., 2007). However, the role of the bilateral hemispheres during aphasia recovery is still unclear. Naming, as an essential function in daily life, is re‐ported to be the earliest spontaneous post‐stroke language function.us, naming may be a useful tool for examining activation in language‐related areas in patients with sub‐acute Broca’s aphasia.
Although many treatment techniques have been used to treat individuals with Broca’s aphasia, including behavior‐ally‐based speech and language therapies and non‐invasive brain stimulation, it is di fficult for these patients to recover speech‐language function (Hamilton et al., 2011; Koyuncu et al., 2016). A more comprehensive understanding of the mechanisms of aphasia will thus be useful in developing more efficacious treatment options. In this study, we used blood oxygenation level‐dependent fMRI to evaluate bilat‐eral cortex activation in patients with Broca’s aphasia 1 to 3 months aer stroke.
Participants and Methods
Participants
Ten patients with aphasia (three females and seven males) were recruited from the Department of Rehabilitation at theird A ffiliated Hospital of Sun Yat‐sen University in China between December 2013 and April 2016.e control group contained ten healthy volunteers (three females and seven males).e two groups were matched in terms of age (aphasia group: 40—70 years, mean ± SD: 55.89 ± 13.37 years; control group: 45—70 years, 55.89 ± 11.78 years) and years of edu‐cation (aphasia group: 11.9 ± 1.4 years; control group: 12.2 ± 0.9 years).e demographic data from the two groups are presented inable 1.
In this study, we only included individuals presenting with stroke‐induced aphasia who had no previous history of aphasia and no prior history of speech, hearing, or neuro‐logical disorders. At enrollment, the duration of onset was 1—3 months post‐stroke. The ten patients were diagnosed with Broca’s aphasiaviathe Western Aphasia Battery (Wang, 1997a, b). We established that they had no severe cognitive impairmentsviathe Mini Mental State Examination (Cock‐rell and Folstein, 1988) (22.7 ± 1.3 scores). All participants met the diagnostic criteria for cerebral infarction or cerebral hemorrhage (Chinese Neurosurgical Society, 1996), as es‐tablished at a national cerebrovascular conference. Single unilateral left‐hemisphere lesion was confirmed by head computed tomography (CT) or (MRI) imaging. The exclu‐sion criteria for aphasia patients included vision and hearing disabilities, MRI contraindication, dysarthria, and stuttering before onset. We also excluded participants with lesions in‐volving bilateral hemispheres and individuals who were not cognitively fi t to complete the study.
The two groups comprised exclusively right handed na‐tive Mandarin speakers (Li, 1983). The study protocol was approved by the Ethics Committee of the Third Affiliated Hospital of Sun Yat‐sen University in China (No. [2015]2‐78). All participants provided written informed consent prior to engaging in the study.
Stimuli
We extracted a set of black and white line drawings from the normalized data set used by Snodgrass and Vanderwat (Snodgrass and Vanderwart, 1980). We randomly chose 27 images (Supplementary Figure 1), including animals and common objects, for use in the picture‐naming task. Before the scanning sessions, we used a di ff erent set of pictures to train the participants in the procedure and ensure that they understood the fMRI paradigm.
Picture-naming task
The experiment had a block design such that a 30‐sec‐ond task (overt picture‐naming) period alternated with a 30‐second rest ( fi xation) period. We used E‐prime soware to present the stimuli on a computer screen (Psychology Soware Tools, Inc., Pittsburgh, PA, USA).ree trials com‐prised a session, and there were three sessions in total. Each session lasted 3 minutes and 6 seconds. Between each ses‐sion, a fi xation cross was shown in the center of the screen for 6 seconds. In the picture‐naming condition, a picture cue was shown for 2 seconds. Afterwards, a fixation cross was presented in the center of the screen for 7 seconds, during which the participants were asked to name the object depict‐ed by the picture. Aer a response had been made, a blank screen was presented for 1 second. This acted as a buffer between adjacent target pictures.e task sequence is shown in Figure 1.
fMRI image acquisition
We used a GE Signa 1.5 Tesla scanner (version 5.6) (General Electric Company, Milwaukee, WI, USA) with a gradient Echo Planar Imaging sequence to acquire image data. Scan‐ning parameters for conventional T1‐weighted imaging, transverse sections: reaction time = 530 ms, echo time = 13 ms. Scanning parameters for T2‐weighted imaging, trans‐verse sections: reaction time = 4,500 ms, echo time = 102 ms. FLAIR sequence scanning parameters: reaction time = 8,800 ms, echo time = 120 ms, fl ip angle = 90°. Slice thickness of scanning = 5 mm, slice gap = 1 mm, fi eld of view = 240 × 240 mm, matrix = 320 × 256. Transverse sections were based on the orbitomeatal line, with a total of 12 slices. Coronal sections were vertical to the orbitomeatal line, with a total of 9 slices. 3D image scanning parameters: fast spoiled gradient recalled sequence, echo time = 1.8 ms, reaction time = 8.5 ms, fl ip angle = 15°, fi eld of view = 240 × 240 mm, matrix = 256 × 192, slice thickness = 1.4 mm. For BOLD‐fMRI im‐ages, we used a gradient recalled echo‐echo planar imaging sequence, with a scanning time of 186 seconds. Scanning pa‐rameters were: reaction time = 3,000 ms, echo time = 40 ms, fl ip angle = 60°, fi eld of view = 240 × 240 mm, matrix = 64 × 64, slice thickness = 7 mm, slice gap = 2 mm. Scan range: AC‐PC line as baseline, for a total of 12 slices.e top edge included the frontal lobe and parietal lobe.e bottom edge was locationed around the tentorium cerebelli.
Image postprocessing
Brain structure and functional images were produced using Viewforum (General Electric Company, Milwaukee, WI, USA). The DICOM images were transformed to img. We used SPM8 soware (Wellcome Trust Centre for Neuroim‐aging, London, UK) for data analysis. We used the Talairach Client (www.talairach.org, the University of Texas Health Sciences Center San Antonio (UTHSCSA), San Antonio, TX, USA) to transform the active brain area into Talairach coordinates. Finally, we located the Brodmann (BA) areas.
Assignment of brain activity
We measured the volume and intensity of the active brain regions. We used group analysis methods to calculate acti‐vation at a group level. In our individual level analysis, the threshold of brain activity wasP< 0.005, and the activity volume was set at 10 voxels.e level result in each groupwas analyzedviaa one‐samplet‐test. We used two‐samplet‐tests (P< 0.05, voxel = 10) to compare activities in the two groups.
Results
Brain activation in normal control participants in the picture-naming task
When comparing the picture‐naming condition with the fi x‐ation condition (task > baseline) in normal participants at an individual level, we found brain activation predominantly in the lehemisphere, and both the leand right inferior fron‐tal gyri were activated in all participants. In addition, the lemiddle, superior frontal, and superior temporal gyri, as well as the right superior temporal gyrus, were activated in 90% of participants. Comparing these conditions (task > baseline) on a group level, multiple brain areas were engaged during the picture‐naming process. We found substantially stronger activity in the lecompared with the right hemisphere. Acti‐vated regions included the bilateral inferior frontal and supe‐rior temporal gyri, insula, basal ganglia, leanterior central gyrus, thalamus, and superior frontal, middle temporal, and middle frontal gyri (Figure 2 andable 2).
Brain activation in Broca’s aphasia patients in the picture-naming task
Whole‐brain analyses of patients on an individual level re‐vealed: (1) Activation in the visual processing cortices related to visual perception in both the left and right hemispheres (middle frontal, middle temporal, lingualis, and fusiform gyri) in all patients. (2) Areas activated during picture‐naming were located in the leinferior frontal gyrus (i.e., Brodmann area 44), right inferior frontal gyrus, and bilateral superior temporal gyrus. (3) All patients showed less activation volume in the lecompared with the right hemisphere, although this was not signi fi cant. Group level analysis showed a signi fi cant decrease in brain activation (mainly in the lebasal ganglia and insula) in the left hemisphere compared with controls. The activated brain areas in the right hemisphere were the superior temporal, fusiform, superior frontal, middle tempo‐ral, parahippocampal, and middle frontal gyri.e activated bilateral areas are presented in Figure 3 andable 2.
Activation in Broca’s aphasiavs. control group
When comparing the activated areas in the two groups, we found that, in the control group, the left hemisphere was dominant during the picture‐naming process, speci fi cally the anterior and posterior frontal area. Compared with the control group, we mainly observed a decrease in the lehemisphere, including the left posterior central, middle frontal, inferior frontal, and superior frontal gyri in the patient group. Simulta‐neously, the right inferior frontal and middle frontal gyri were more strongly activated in the controls compared with the patients. For the patients, the bilateral temporal‐parietal lobes played an important role in the picture‐naming task (able 2).
Activation volume of left and right hemispheres
Comparing the activated volume between the aphasia and control groups revealed that the activated volume was sig‐ni fi cantly less in the right hemisphere than in the lehemi‐sphere (t= 2.715,P= 0.024 < 0.05). No statistically signi fi‐cant di ff erence was found in the aphasia group (t= –1.975,P= 0.098 > 0.05).e activated volume in the ipsilateral lehemisphere was signi fi cantly less in the aphasia group than in the control group (t= 3.583,P= 0.004 < 0.01). We did not fi nd any di ff erences between the two groups in terms of the activated volume of the right hemisphere (t= 1.931,P= 0.070> 0.05) (able 3).
Discussion
Language region changes induced by stroke
In many clinical cases, post‐stroke aphasia is associated with infarction near the left middle cerebral artery. Indeed, the bulk of behavioral and imaging aphasia studies have exam‐ined patients with language deficits that are attributable to lesions in this vascular territory.e notion that language is represented in the brain by a network of functionally distinct interacting processing centers dates back to the 19thcentury with the pioneering discoveries of Broca and Wernicker. Initially, the language network was envisioned as comprising a simple expression area and an understanding area. For most healthy individuals, language is predominantly but not exclusively represented in a network of regions in the left hemisphere surrounding Broca’s area and Wernicke’s area. The idea that lesions leading to aphasia are located in the leside of the brain corresponded with this classical model. Strokes that result in aphasia typically damage this leperi‐sylvian network (the regions around fi ssure of sylvius). Cur‐rent models of language representation also emphasize the role of dorsal and ventral processing streams, and indicate that post‐stroke aphasia can result from damage to either or both stream.
Advances in functional neuroimaging have yielded a num‐ber of insights into the changes that occur in brain activity during language tasks in patients with post‐stroke aphasia. Stroke recovery mechanisms mainly involve neural plasticity,i.e., the activation of regions peripheral to the lesion area or the contralateral brain regions. Similarly, post‐stroke aphasia recovery mechanisms involve neural plasticity. Most studies investigating language reorganization in patients with large perinatal vascular lesions have suggested that the right hemi‐sphere network mirrors that of the normal lehemisphere, both in the classical language areas (Staudt et al., 2002; Til‐lema et al., 2008) and other regions, such as the cerebellum (Lidzba et al., 2008). However, increased activity in periph‐eral regions has been noted in aphasia patients with small lesions. Interestingly, a study (Sebastian et al., 2016) found that improvements in naming accuracy from the acute stage to the chronic stage of aphasia following stoke corresponded with increased connectivity within and between language re‐gions in the leand right hemispheres.ese fi ndings point to the importance of reorganizing patterns of brain activity and connectivity aer stoke to make relevant connections in the language network.

Figure 1imeline of task design in one session.

Figure 2 Activation map of the whole-brain analyses related to the main task e ff ect by functional magnetic resonance imaging (task >baseline) in control group.
How and where brain regions involved in language trans‐fer may depend in part on the stage of neural development at the time of injury aer stoke.e mechanisms by which language reorganizes are still not well understood.
Findings of the present study
In this study, we examined the topography of language re‐organization by studying healthy adults and patients with Broca’s aphasia. We found di ff erences in the features of brain activation in the two groups.
Language-related brain activation in healthy controls
Naming is an important mode of expression. Picture‐naming is commonly used in fMRI studies. Picture‐naming requires the perception of the object depicted in the picture, as well as the extraction of information about the object, including: (1) visual analysis of the object; (2) matching the results of the visual analysis with the structural features of objects stored in memory; (3) search for a semantic description of the ob‐ject, followed by speech coding; (4) speaking the name of theobject under the control of motor voice‐related brain areas (Menke et al., 2009).
able 1 Demographic data from patients with Broca’s aphasia and normal controls
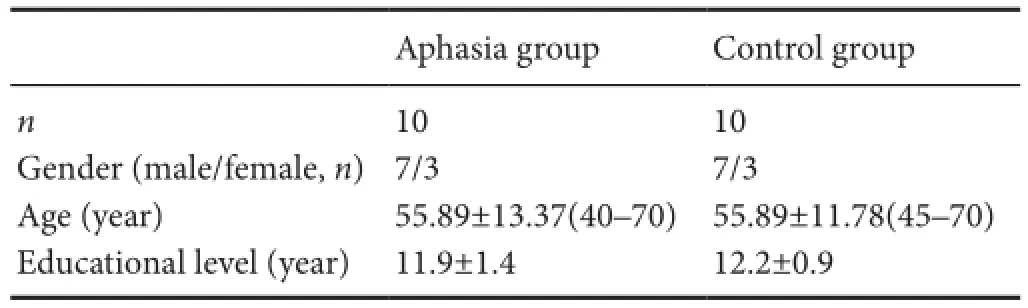
able 1 Demographic data from patients with Broca’s aphasia and normal controls
Age and educational level are expressed as the mean ± SD.
Aphasia group Control groupn10 10 Gender (male/female,n)7/3 7/3 Age (year) 55.89±13.37(40–70) 55.89±11.78(45–70) Educational level (year) 11.9±1.4 12.2±0.9
able 3 Activated volume in bilateral hemispheres of both groups

able 3 Activated volume in bilateral hemispheres of both groups
Data are expressed as the mean ± SD, with 10 participants in each group. **P< 0.01,vs. aphasia group (two‐samplet‐test); #P< 0.05,vs. lehemisphere (one‐samplet‐test).
Group Lehemisphere (voxels) Right hemisphere (voxels) Control 1,006.00±635.62**248.33±197.07#Aphasia 754.60±423.80 414.33±332.23

Figure 3 Activation map of the whole-brain analyses related to the main task e ff ect by fMRI (task > baseline) in the aphasia group.
Cabeza and Nyberg (2000) reported that the visual process used during picture‐naming is related to the lemiddle frontal gyrus, bilateral middle temporal gyrus, and the lower‐middle part of the occipitotemporal cortex.e middle frontal, tempo‐ralis medius, and fusiform gyri, as well as the occipital lobe, are engaged during visual processing of an object. Among these, the left fusiform gyrus may participate in image recognition (Xue et al., 2006). Broca’s area is known for its input and output functions. Broca’s area covers not only the lepars opercularis gyri, but also the middle frontal and anterior central gyri (Cat‐ani et al., 2005, 2007). Previously, we found signi fi cant activa‐tion in the leinferior frontal, lemiddle frontal, leanterior central, and lesuperior frontal gyri during picture‐naming in healthy adults (Xie, 2011), which supports the viewpoint that Broca’s area, along with multiple frontal areas, is engaged in phonologic and semantic processes.
fMRI studies have shown that the bilateral superior temporal cortex is activated during phoneme extraction.Additionally, the lesuperior temporal cortex is implicated in phonological function. The left middle temporal gyrus and inferior temporal gyrus are correlated with semantic processes, and are connectedviaBrodmann area 37 (Xie, 2011). In this study, the left superior temporal gyrus, in‐ferior parietal lobule, and middle temporal gyrus were activated in most of the healthy control participants.ese areas might be involved in phonologic coding and semantic extraction.
able 2 Activation during picture-naming task in aphasia and control groups (task > baseline)
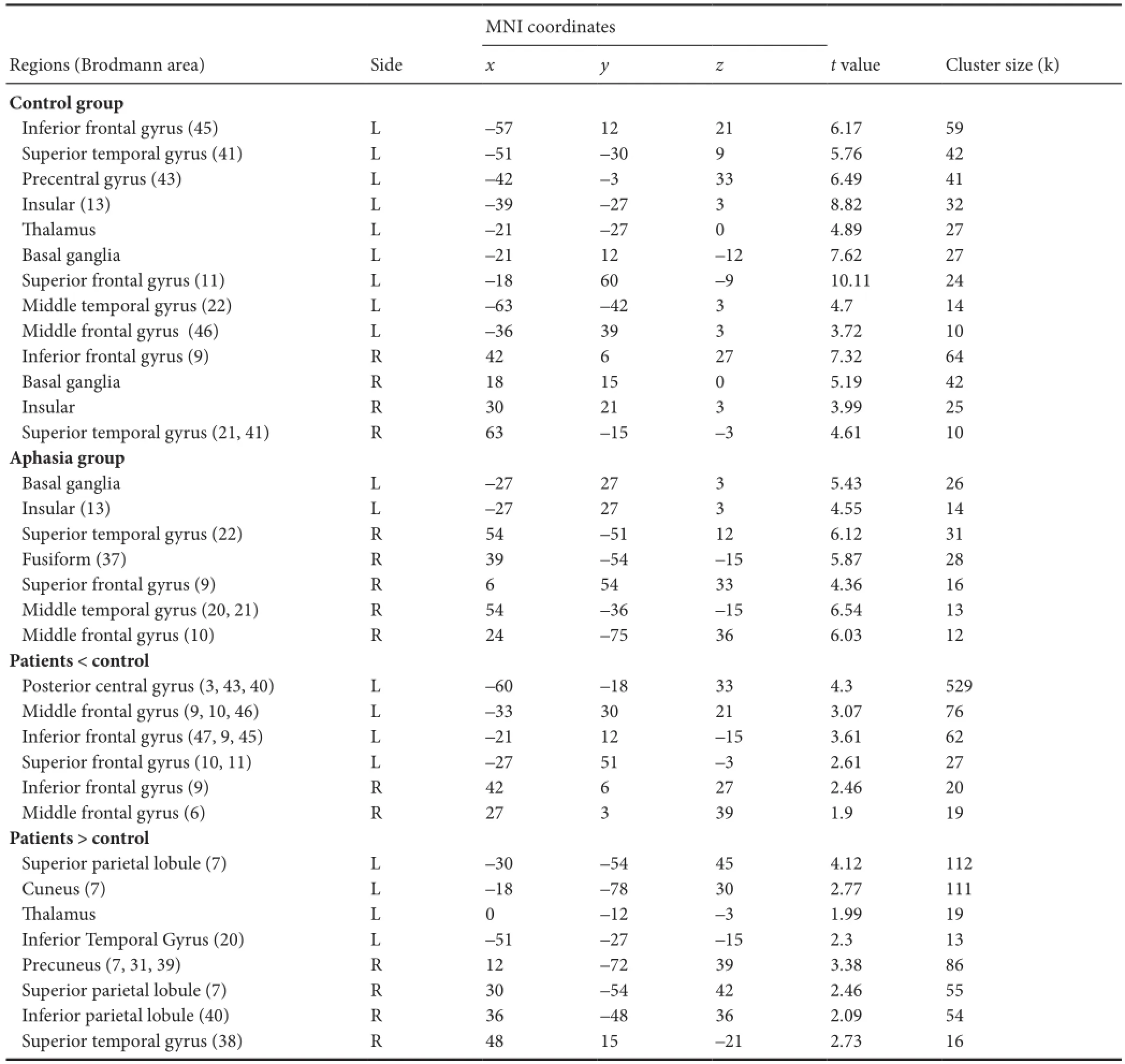
able 2 Activation during picture-naming task in aphasia and control groups (task > baseline)
Multiple comparisons: Family‐wise error (FWE) cluster‐corrected atP< 0.05 determined by Monte Carlo simulation.
MNI coordinates Regions (Brodmann area) Sidexyztvalue Cluster size (k) Control group Inferior frontal gyrus (45) L –57 12 21 6.17 59 Superior temporal gyrus (41) L –51 –30 9 5.76 42 Precentral gyrus (43) L –42 –3 33 6.49 41 Insular (13) L –39 –27 3 8.82 32alamus L –21 –27 0 4.89 27 Basal ganglia L –21 12 –12 7.62 27 Superior frontal gyrus (11) L –18 60 –9 10.11 24 Middle temporal gyrus (22) L –63 –42 3 4.7 14 Middle frontal gyrus (46) L –36 39 3 3.72 10 Inferior frontal gyrus (9) R 42 6 27 7.32 64 Basal ganglia R 18 15 0 5.19 42 Insular R 30 21 3 3.99 25 Superior temporal gyrus (21, 41) R 63 –15 –3 4.61 10 Aphasia group Basal ganglia L –27 27 3 5.43 26 Insular (13) L –27 27 3 4.55 14 Superior temporal gyrus (22) R 54 –51 12 6.12 31 Fusiform (37) R 39 –54 –15 5.87 28 Superior frontal gyrus (9) R 6 54 33 4.36 16 Middle temporal gyrus (20, 21) R 54 –36 –15 6.54 13 Middle frontal gyrus (10) R 24 –75 36 6.03 12 Patients < control Posterior central gyrus (3, 43, 40) L –60 –18 33 4.3 529 Middle frontal gyrus (9, 10, 46) L –33 30 21 3.07 76 Inferior frontal gyrus (47, 9, 45) L –21 12 –15 3.61 62 Superior frontal gyrus (10, 11) L –27 51 –3 2.61 27 Inferior frontal gyrus (9) R 42 6 27 2.46 20 Middle frontal gyrus (6) R 27 3 39 1.9 19 Patients > control Superior parietal lobule (7) L –30 –54 45 4.12 112 Cuneus (7) L –18 –78 30 2.77 111alamus L 0 –12 –3 1.99 19 Inferior Temporal Gyrus (20) L –51 –27 –15 2.3 13 Precuneus (7, 31, 39) R 12 –72 39 3.38 86 Superior parietal lobule (7) R 30 –54 42 2.46 55 Inferior parietal lobule (40) R 36 –48 36 2.09 54 Superior temporal gyrus (38) R 48 15 –21 2.73 16
Roles of the right hemisphere in the picture-naming process
In sum, the language functional area is a whole brain network, where, in addition to Broca’s area, the lesupe‐rior frontal gyrus, middle frontal gyrus, inferior parietal lobule, right inferior frontal gyrus, superior temporal gyrus, and middle temporal gyrus play a role in language expression.
Role of the right hemisphere in picture-naming in aphasia patients
We found that activated brain areas in aphasia patients were reduced significantly compared with healthy participants, which is consistent with a previous study (Menke et al., 2009). Our data showed that the visual processing cortices were activated in all participants, such as the fourth visual cortex (BA17–20), left middle frontal gyrus (BA9), lingual gyrus (BA19), and fusiform gyrus (BA37). Nevertheless, the intensity of activation was lower than that in normal partic‐ipants. This indicates that aphasia patients may have weak connections among visual perception areas that process ob‐ject information. For instance, they may be able to obtain the shape of an object visually, but may fi nd it di ffi cult to match the speci fi c information stored in the brain with the object (Warrington and James, 1988).
Aphasia is caused by brain lesions that directly or directly hinder normal language function (Price et al., 2001). The speech deficit caused by a specific lesion in Broca’s area is mild and recovery may be faster than that for the lesions seen in our patient group. Indeed, the lesions leading to a severe speech de fi cit generally include, in addition to Broca’s area (Mohr et al., 1978), the lepars opercularis, lemiddle frontal gyrus, and left central motor cortex (Catani et al., 2005, 2007). In our study, 10 patients in the aphasia group were diagnosed as having moderate to severe aphasia, and they experienced multiple brain lesions including Broca’s area.us, we were not surprised to fi nd that bilateral brain activated volumes were lower in patients compared with controls. We found a significant decrease in volume in the lehemisphere of patients, but not in the right hemisphere.e activated volume in the right hemisphere was reduced in aphasia patients, although this was not statistically sig‐nificant. Specifically, we found decreased activation in the inferior frontal gyrus, parahippocampal gyrus, and middle frontal gyrus in the right hemisphere. However, we observed normal and increased activation in other right hemisphere brain areas.us, language‐related areas in the right hemi‐sphere appear to be a ff ected by distant e ff ects of lehemi‐sphere lesions.
Some studies have reported that the right inferior frontal gyrus is strongly activated in aphasia patients. In this popu‐lation, language function is improved when the right inferior frontal gyrus is inhibited (Naeser et al., 2004). However, right hemisphere activation could promote recovery (Blank et al., 2003). Interestingly, previous studies have not shown decreased activation in the right inferior frontal gyrus in ear‐ly stage aphasia patients compared with control participants.us, this fi nding is not consistent with previous results that focus on chronic aphasia patients rather than those in the early stages.is fi nding suggests that the right inferior fron‐tal gyrus plays di ff erent roles during language recovery. One of the possible mechanisms is the involvement of the right inferior frontal gyrus in normal language processes.e dis‐tant e ff ect of the lesion would decrease activation in the early stage of recovery. As the recovery process continues, the right inferior frontal gyrus compensates for the original lan‐guage function. In contrast, in the chronic phase, activation in this region would be too high to inhibit lesion recovery in the lehemisphere through the corpus callosum.is is a valuable question for future investigations with increased samples and a longitudinal design, which could evaluate the mechanisms of reconstruction of functional paths in di ff er‐ent recovery phases.
Limitations
Implications of the current literature for further research regarding mechanisms of plasticity and reorganization
In this study, we found that individuals with subacute Broca’s aphasia exhibited a decrease in left hemisphere and right inferior frontal gyrus activation during a picture‐naming task. Additionally, we observed relative increases in certain activated brain regions in the right hemisphere in the patient population. Our fi ndings are consistent with a comprehen‐sive neuropsychological model of recovery mechanisms underlying the progression of aphasia. The reorganization that occurs in the early stages of Broca’s aphasia may be dif‐ferent from that in chronic aphasia. Our data indicate that future research examining temporal changes during aphasia recovery may be bene fi cial, particularly for guiding non‐in‐vasive stimulation therapies (rTMS, tDCS), which are based on brain activation patterns and corresponding functions, in the early stages of post‐stroke aphasia.
Conclusions
To the best of our knowledge, this is the fi rst study to use functional imaging to examine brain activation in patients with subacute Broca’s aphasia during a picture‐naming task. Our results show that language expression is associated with multiple brain areas.e right hemisphere is engaged in normal speech generation. We found that brain lesion and associated remote e ff ects led to a decrease in activity in language functional areas in post‐stroke aphasia patients. Particularly, the left hemisphere and right inferior frontal gyrus showed a clear decrease in activation. However, acti‐vation in sections of the right hemisphere increased in this population, compared with healthy controls.e right infe‐rior frontal gyrus plays multiple roles in language recovery during di ff erent periods in post‐stroke aphasia. Further in‐vestigation regarding the mechanisms of aphasia will likely lead to improved treatments options, thus reducing family and social burden.
Declaration of patient consent:
Author contributions:WHQ served as a guarantor of integrity of the entire paper, study design, paper editing and paper review. HXW participated in clinical studies, data acquisition, statistical analysis and paper editing. QLY was in charge of data analysis, paper preparation and paper editing. ZK participated in clinical studies and data acquisition. KL served as a guarantor of study integrity. GRQ participated in paper editing. ZCC participated in paper editing and paper review. CQX participated in literature research. GFW participated in de fi nition of intellectual content. SQC participated in study concept. All authors approved the fi nal version of the paper.
Con fl icts of interest:None declared.
Supplementary information:Supplementary data associated with this article can be found, in the online version, by visiting www.nrronline.org.
Plagiarism check:This paper was screened twice using CrossCheck to verify originality before publication.
Peer review:
Amino ff MJ, Daro ff RB (2014) Encyclopedia of the Neurological Sci‐ences. Amsterdam: Elsevier.
Benvenuti SM, Zanatta P, Longo C, Mazzarolo AP, Palomba D (2012) Preoperative cerebral hypoperfusion in the le, not in the right, hemisphere is associated with cognitive decline aer cardiac sur‐gery. Psychosom Med 74:73‐80.
Berthier ML (2005) Poststroke aphasia : epidemiology, pathophysiology and treatment. Drugs Aging 22:163‐182.
Blank SC, Bird H, Turkheimer F, Wise RJ (2003) Speech production after stroke: The role of the right pars opercularis. Ann Neurol 54:310‐320.
Bottini G, Corcoran R, Sterzi R, Paulesu E, Schenone P, Scarpa P, Frac‐kowiak RS, Frith D (1994)e role of the right hemisphere in the interpretation of fi gurative aspects of language A positron emis‐sion tomography activation study. Brain 117:1241‐1253.
Cabeza R, Nyberg L (2000) Imaging cognition II: An empirical review of 275 PET and fMRI studies. J Cogn Neurosci 12:1‐47.
Cambier J, Elghozi D, Signoret JL, Henin D (1983) Contribution of the right hemisphere to language in aphasic patients. Disappearance of this language aer a right‐sided lesion. Rev Neurol (Paris) 139:55‐63.
Catani M, Jones DK, ff ytche DH (2005) Perisylvian language networks of the human brain. Ann Neurol 57:8‐16.
Catani M, Allin MPG, Husain M, Pugliese L, Mesulam MM, Mur‐ray RM, Jones DK (2007) Symmetries in human brain language pathways correlate with verbal recall. Proc Natl Acad Sci U S A 104:17163‐17168.
Chie ff o R, Ferrari F, Battista P, Houdayer E, Nuara A, Alemanno F, Abu‐talebi J, Zangen A, Comi G, Cappa SF, Leocani L (2014) Excitatory deep transcranial magnetic stimulation with H‐coil over the right homologous Broca’s region improves naming in chronic post‐stroke aphasia. Neurorehabil Neural Repair 28:291—298.
Chinese Neurosurgical Society (1996)e fourth National Conference of cerebrovascular disease.e diagnosis of cerebrovascular dis‐ease academic standards. Zhonghua Shenjing Ke Zazhi 3:10‐11.
Cockrell JR, Folstein MF (1988) Mini‐mental state examination (MMSE). Psychopharmacol Bull 24:689‐692.
Crosson B, Moore AB, Gopinath K, White KD, Wierenga CE, Gaiefsky ME, Fabrizio KS, Peck KK, Soltysik D, Milsted C, Briggs RW, Con‐way TW, Rothi LJ (2005) Role of the right and lehemispheres in recovery of function during treatment of intention in aphasia. J Cognit Neurosci 17:392‐406.
Finger S, Buckner RL, Buckingham H (2003) Does the right hemi‐sphere take over aer damage to Broca’s area? the Barlow case of 1877 and its history. Brain Lang 85:385‐395.
Hamilton RH (2016) Neuroplasticity in the language system: Reorga‐nization in post‐stroke aphasia and in neuromodulation interven‐tions. Restor Neurol Neurosci 34:467‐471.
Hamilton RH, Chrysikou EG, Coslett B (2011) Mechanisms of aphasia recovery aer stroke and the role of noninvasive brain stimulation. Brain Lang 118:40‐50.
Hillis AE (2007) Aphasia: progress in the last quarter of a century. Neu‐rology 69:200‐213.
Jang SH, Jang WH (2016) Di ff erence in cortical activation during use of volar and dorsal hand splints: a functional magnetic resonance imaging study. Neural Regen Res 11:1274‐1277.
Krashen SD (1973) Lateralization, language learning, and the critical period: some new evidence. Lang Learn 23:63‐74.
Koyuncu E, Çam P, Altınok N, Çallı DE, Duman TY, Özgirgin N (2016) Speech and language therapy for aphasia following subacute stroke. Neural Regen Res 11:1591‐1594.
Lidzba K, Wilke M, Staudt M, Krägeloh‐Mann I, Grodd W (2008) Reorganization of the cerebro‐cerebellar network of language pro‐duction in patients with congenital le‐hemispheric brain lesions. Brain Lang 106:204‐210.
Müller Ra, Rothermel RD, Behen Me, Muzik O, Chakraborty PK, Chugani HT (1999) Language organization in patients with early and lateleft‐hemisphere lesion: a pet study. Neuropsychologia 37:545‐557.
Mattioli F, Ambrosi C, Mascaro L, Scarpazza C, Pasquali P, Frugoni M, Magoni M, Biagi L, Gasparotti R (2014) Early aphasia rehabilita‐tion is associated with functional reactivation of the leinferior frontal gyrus: a pilot study. Stroke 45:545‐552.
Menke R, Meinzer M, Kugel H, Deppe M, Baumgärtner A, Schiauer H,omas M, Kramer K, Lohmann H, Flöel A, Knecht S, Breiten‐stein C (2009) Imaging short‐ and long‐term training success in chronic aphasia. BMC Neurosci 10:118.
Mohr JP, Pessin MS, Finkelstein S, Funkenstein HH, Duncan GW, Da‐vis KR (1978) Broca aphasia: pathologic and clinical. Neurology 28:311‐324.
Naeser MA, Martin PI, Baker EH, Hodge SM, Sczerzenie SE, Nicholas M, Palumbo CL, Goodglass H, Wing fi eld A, Samaraweera R, Har‐ris G, Baird A, Renshaw P, Yurgelun‐Todd D (2004) Overt prop‐ositional speech in chronic nonfluent aphasia studied with the dynamic susceptibility contrast fMRI method. Neuroimage 22:29‐41.
Noppeney U, Price CJ, Duncan JS, Koepp MJ (2005) Reading skills after left anterior temporal lobe resection: an fMRI study. Brain 128:1377‐1385.
Park BY, Park H (2016) Connectivity differences between adult male and female patients with attention deficit hyperactivity disorder according to resting‐state functional MRI. Neural Regen Res 11:119‐125.
Pedersen PM, Jørgensen HS, Nakayama H, Raaschou HO, Olsen TS (1995) Aphasia in acute stroke: incidence, determinants, and re‐covery. Ann Neurol 38:659‐666.
Price CJ, Warburton EA, Moore CJ, Frackowiak RS, Friston KJ (2001) Dynamic diaschisis: anatomically remote and context‐sensitive human brain lesions. J Cogn Neurosci 13:419‐429.
Rapport RL, Tan CT, Whitaker HA (1983) Language function and dys‐function among Chinese‐ and English‐speaking polyglots: cortical stimulation, Wada testing, and clinical studies. Brain Lang 18:342‐366.
Rochon E, Leonard C, Burianova H, Laird L, Soros P, Graham S, Grady C (2010) Neural changes aer phonological treatment for anomia: An fMRI study. Brain Lang 114:164‐179.
Saur D, Lange R, Baumgaertner A, Schraknepper V, Willmes K, Rijntjes M, Weiller C (2006) Dynamics of language reorganization after stroke. Brain 129:1371‐1384.
Sebastian R, Long C, Purcell JJ, Faria AV, Lindquist M,Jarso S, Race D, Davis C, Posner J, Wright A, Hillis AE (2016). Imaging network level language recovery aer lePCA stroke. Restor Eurol Neuro‐sci 34:473‐489.
Siirtola M, Narva EV, Siirtola T (1977) On the occurrence and progno‐sis of aphasia in patients with cerebral infarction. Scand J Soc Med Suppl 14:128‐133.
Snodgrass JG, Vanderwart M (1980) A standardized set of 260 pictures: norms for name agreement, image agreement, familiarity, and vi‐sual complexity. J Exp Psychol Hum Learn 6:174‐215.
Staudt M, Lidzba K, Grodd W, Wildgruber D, Erb M, Krägeloh‐Mann I (2002) Right‐hemispheric organization of language following early le‐sided brain lesions: functional MRI topography. Neuroimage 16:954‐967.
Sza fl arski JP, Eaton K, Ball AL, Banks C, Vannest J, Allendorfer JB, Page S, Holland SK (2011) Post‐stroke aphasia recovery assessed with fMRI and a picture identi fi cation task. J Stroke Cerebrovasc Dis 20:336‐345.
Thiel A, Zumbansen A (2016) The pathophysiology of post‐stroke aphasia: A network approach. Restor Neurol Neurosci 34:507‐518.
Tillema JM, Byars AW, Jacola LM, Schapiro MB, Schmithorst VJ, Sza‐fl arski JP, Holland SK (2008) Cortical reorganization of language functioning following perinatal left MCA stroke. Brain Lang 105:99‐111.
Warrington EK, James M (1988) Visual apperceptive agnosia: a clini‐co‐anatomical study of three cases. Cortex 24:13‐32.
Winhuisen L, Thiel A, Schumacher B, Kessler J, Rudolf J, Haupt WF, Heiss WD (2007)e right inferior frontal gyrus and poststroke aphasia: a follow‐up investigation. Stroke 38:1286‐1292.
Xie Q (2011) Progresses in functional magnetic resonance imaging study of linguistic function areas. Yixue Zongshu 17:3315‐3317.
Xue G, Chen C, Jin Z, Dong Q (2006) Language experience shapes fusi‐form activation when processing a logographic arti fi cial language: An fMRI training study. Neuroimage 31:1315‐1326.
Copyedited by Koke S, Haase R, Yu J, Li CH, Qiu Y, Song LP, Zhao M
10.4103/1673-5374.198996
Accepted: 2016-11-10
*Correspondence to: Wei-hong Qiu, M.D. or Kui Li, q-weihong@163.com or likui3@126.com.

- 中国神经再生研究(英文版)的其它文章
- Restoring axonal localization and transport of transmembrane receptors to promote repair within the injured CNS: a critical step in CNS regeneration
- Information for Authors -Neural Regeneration Research
- A new computational approach for modeling diffusion tractography in the brain
- Celebration of the 10thAnniversary of Neural Regeneration Research
- Terapeutic potential of brain-derived neurotrophic factor (BDNF) and a small molecular mimics of BDNF for traumatic brain injury
- Blood microRNAs as potential diagnostic markers for hemorrhagic stroke