
2016年春季西太平洋M2海山浮游植物粒级结构与初级生产力研究*
2017-03-31 06:18孙晓霞梁俊华田梓杨
海洋与湖沼 2017年6期
代 晟 孙晓霞, 梁俊华 田梓杨 刘 涛
(1. 中国科学院海洋研究所 山东胶州湾海洋生态系统国家野外科学观测研究站 青岛 266071; 2. 中国科学院大学 北京100049; 3. 青岛海洋科学与技术国家实验室 海洋生态与环境科学功能实验室 青岛 266071)
海山(seamount)通常是指深海大洋中位于水面以下,高度大于1000m的隆起地形(Eppet al, 1989)。海山是研究海洋生物活动的热点区域, 全球范围内估计有 3万座海山(张均龙等, 2013), 但有研究记录的只有不到 1%(Clarket al, 2012; Djurhuuset al, 2017; Readet al, 2017)。由于海山独特的地形特征, 一些学者认为洋流在流经海山的复杂地形时, 形成内波、上升流和泰勒柱(Geninet al, 1985; Doweret al, 1992)等物理过程,促进了海水的垂直混合, 有利于浮游植物的生长(Whiteet al, 2007), 进而支撑起了海山区域较高的浮游生物、底栖动物和鱼类的生物量, 特别是在一些寡营养海区(Clarket al, 2010; Letessieret al, 2017), 这一理论被称为“海山效应”的经典假说(Doweret al,1996)。在 Cobb Seamount (Doweret al, 1992)和 Minamikasuga Seamount (Geninet al, 1985)海山山顶附近发现了由泰勒柱引起的浮游植物生物量和初级生产力升高的现象, 但该现象在同一海山并不长期存在, 如Cobb Seamount (Comeauet al, 1995), 还有一些海山如Britannia Seamount (Younget al, 2011)、Vercelli Seamount (Misicet al, 2012)和Komahashi No.2 Seamount (Odateet al, 1998)等的研究并不支持该假说。
热带西太平洋是海山系统分布最集中的海区之一(张均龙等, 2013), 该海域长期存在物理分层现象,表层具有高温寡营养盐和低生产力的特征(Messiéet al, 2006), 受营养盐和光照的限制, 叶绿素最大层(deep chlorophyll maximum layer, DCML)常出现在温跃层上层附近 (Cordeiroet al, 2013)。以往研究表明该海区初级生产者主要为微型和微微型浮游植物(Zhanget al, 2012), 聚球藻和原绿球藻为优势藻类(Le Bouteilleret al, 1992)。该区域的海山属于典型的寡营养海域海山, 目前对该区域海山的各项研究尚处于起步阶段, 浮游植物与初级生产力的研究鲜有报道, 海山是否会引起生产力升高等现象尚不明确。
本研究于 2016年 3月热带西太平洋 M2海山(11.19°—11.49°N, 139.13°—139.55°E)开展环境因子、分级叶绿素a浓度和初级生产力等研究, 首次对 M2海山区浮游植物分布特征和粒级结构, 初级生产力结构特征进行了分析, 并主要与张文静等(2016)报道的同海域的雅浦Y3海山的浮游植物粒级结构与环境因素进行了对比, 以期了解M2海山区浮游植物粒级结构、分布特征与环境因子之间的关系, M2海山的初级生产力情况, 以及导致不同海山之间叶绿素a浓度和粒级结构差异的原因, 为深入了解热带寡营养海域海山生态系统特征与初级生产过程奠定基础。
1 材料与方法
1.1 采样站位
2016年3月14—24日在热带西太平洋近马里亚纳 海 沟 的M2 海 山 (11.19°—11.49°N, 139.13°—139.55°E)分 A、B两个断面, 共 21个站位对分粒级叶绿素a浓度和分粒级初级生产力进行了调查。采样站位如图1所示。
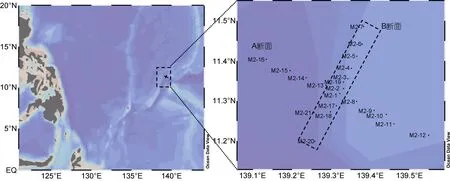
图1 M2海山采样站位图(右图方框代表B断面分布范围)Fig.1 Sampling stations of Seamount M2 (Transections A and B)
1.2 研究方法
各站温度、盐度的垂直剖面数据用 CTD(SBE911)进行测量, 其中 A 断面温盐数据在采集时,CTD传感器出现故障, 导致该断面温盐数据缺失。
根据海洋调查规范(GB/T12763.6-2007), 采用Niskin采水器分 0、15、30、50、75、100、150、200m共8层采集水样后, 先用孔径为200µm的筛绢过滤,去除水体中的浮游动物, 之后每层取 2000mL水, 分别用 20µm 筛绢、2µm Whatman Nuclepore Track-Etch滤膜和 0.7µm Whatman GF/F 滤膜过滤, 取得 20—200µm (小型浮游植物, micro-phytoplankton)、2—20µm (微型浮游植物, nano-phytoplankton)和 0.7—2µm (微微型浮游植物, pico-phytoplankton)的叶绿素a样品, 同时取 2000mL水, 用 0.7µm Whatman GF/F滤膜过滤得到总叶绿素a样品。样品避光-20°C冷冻保存带回实验室经90%丙酮萃取24h后, 用Turner-Designs-Model 10荧光计测定并计算叶绿素a浓度。
硝酸盐、亚硝酸盐、磷酸盐、硅酸盐样品分别在0、15、30、50、75、100、150和 200m 水层取水 250ml,按照海洋调查规范(GB12763.4-2007)进行测定, 用0.45µm的混合纤维滤膜过滤后装入250mL高密度聚乙烯瓶中, 用占水样体积千分之二的氯仿固定后于-20°下冷冻保存, 在实验室用连续流动分析仪(QuAAtro, Germany)分析测定。
根据海洋调查规范(GB/T12763.6-2007), 分别在光强为表层的100%、50%、30%、10%、5%和1%的深度上采水, 水样用200μm筛绢过滤, 然后分别加入0.037kBq的14C工作液, 用中性衰光材料控制光照强度, 在具有流动的海水水槽内培养 4—6h, 培养体积为250cm3。培养结束后, 按与叶绿素相同的分级方法过滤样品。滤膜用浓盐酸熏蒸 15min装入闪烁瓶,-20ºC冰冻保存, 带回实验室用XH-6925液体闪烁计数器测定样品的14C放射性活度。按海洋调查规范的方法中的公式计算每一水层初级生产力, 然后根据(公式 1)计算水柱初级生产力, 最后测定水柱叶绿素a浓度, 根据(公式2)计算水柱叶绿素同化系数。
公式1:
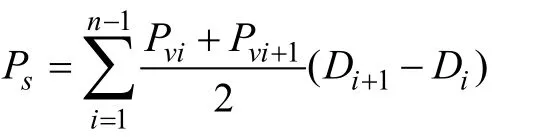
式中,Ps为水柱初级生产力, mgC/(m2⋅d);Pvi为第i层初级生产力, mgC/(m3⋅d);Di为第i层深度, m;n为采样层次数(1≤I≤n-1)。
公式2:
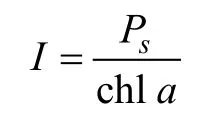
式中,I为水柱叶绿素同化系数, mgC/(mg chla⋅h);Ps为水柱初级生产力, mgC/(m2⋅h); chla为水柱叶绿素a浓度, mg/m2。
1.3 数据分析
使用Ocean Data View(4.5.5)绘制图, 采用Microsoft Office Excel 2016绘制表格, 用R(3.4.0)进行双侧显著性T检验和主成分分析。
2 结果
2.1 温盐分布特征
M2海山B断面温度、盐度分布见图2。总体上,B断面温度和盐度的垂直分布较为相似, 呈现明显的分层现象。且海山东北部靠近山体的区域表现为温度和盐度的涌升。如图 2a所示, 表层水温(<50m)达到了28ºC, 在75m深度处, 开始出现温跃层; 海山山顶的东北部等温线有明显波动, 在 75m水层 M2-03站位温度最低为 26.39ºC, 出现等温线抬升的现象; 东北部等温线总体比西南部密集, 在海山东北部表层盐度出现低值区, 小于 34.6; 盐度最大层均位于100m 附近(图 2b), 对应在 20—26ºC等温线之间, 其波动情况与等温线相吻合。

图2 M2海山B断面温度、盐度垂直分布图Fig.2 Vertical distribution of temperature (℃) and salinity in Seamount M2 across Transection B
2.2 叶绿素a分布特征
2.2.1总叶绿素a分布特征A断面叶绿素a浓度的平均值为(0.094±0.010)mg/m3, 各水层浓度的变化范围为 0.011—0.304mg/m3。表层叶绿素a浓度的平均值为(0.042±0.012)mg/m3, 最高浓度值为0.068mg/m3, 出现在M2海山的M2-12站位, 最低浓度为0.027mg/m3, 出现在M2-08站位。各水层叶绿素a平均浓度最大值为 0.236mg/m3, 位于 100m水层,变化范围为0.177—0.304mg/m3, DCML均在100m水深附近(图 3a)。水柱叶绿素a浓度范围为 15.355—21.367mg/m2, 平均值为(18.898±1.994)mg/m2。A 断面水体平均叶绿素a浓度变化范围为0.077—0.107mg/m3, 在山顶附近的M2-10站位平均叶绿素a浓度最高, 东南部浓度高于西北部。
B断面绿素a浓度的平均值为(0.094±0.008)mg/m3,各水层浓度的变化范围为 0.004—0.262mg/m3。表层叶绿素a浓度的平均值为(0.040±0.005)mg/m3, 最高浓度为 0.047mg/m3, 位于 M2-18站位, 最低为0.029mg/m3, 位于M2-02站位, 各水层叶绿素a平均浓度最大值为 0.231mg/m3, 位于 100m水层, 除M2-20站位外, DCML均为100m(图3b)。水柱叶绿素a浓度范围为16.338—21.197mg/m2, 平均值为(18.710±1.617)mg/m2。B断面水体平均叶绿素a浓度变化范围为 0.082—0.106mg/m3, 在海山的西南部M2-20站位平均叶绿素a浓度最高, 东北部DCML的叶绿素a浓度高于西南部(图3b)。
M2-01站位位于山顶处, 水深只有37m, 没有出现DCML, 表层叶绿素a浓度为0.533mg/m3, 水柱叶绿素a浓度为1.098mg/m2。
海山各个水层叶绿素a浓度分布都不均匀, 最小值在200m水层, 最大层为最小层的10—20倍。A、B断面表层叶绿素a浓度相当。M2海山的叶绿素分布呈现西南部高于东北部, 东南部高于西北部的趋势; 垂直分布上, 随深度的增加, 各个站位的叶绿素a浓度均出现先增加后减小的趋势, DCML多出现在100m 水层, 其波动情况与等温线波动一致, 在海山M2-03站位出现抬升的情况, 整体分布与盐度最大层基本吻合(图2, 图3b)。

图3 M2海山A、B断面分粒级叶绿素a垂直分布情况Fig.3 Vertical distribution of size-fractionated Chl a in Seamount M2 (Transections A and B)
2.2.2分粒级叶绿素a浓度分布海山A断面小-型浮游植物叶绿素a浓度的平均值为0.003mg/m3, 各水层的变化范围为0.0001—0.010mg/m3; 微型浮游植物叶绿素a浓度平均值为 0.011mg/m3, 变化范围为0.0001—0.032mg/m3, 微微型浮游植物叶绿素a浓度平均值为 0.085mg/m3, 各水层的变化范围为0.0113—0.274mg/m3。海山B断面各粒径级叶绿素a的结构与A断面相似, 其浓度也相近, 平均浓度分别是 0.003、0.009、0.073mg/m3, 各水层的变化范围分别为 0.0001—0.011、0.0012—0.024、0.0017—0.248mg/m3。三种粒级的浮游植物叶绿素a浓度最大层均与总叶绿素a浓度最大层相吻合, 在100m附近。其中小型浮游植物和微型浮游植物叶绿素a浓度在海山斜坡附近出现高值(图3c, d, e, f)。
两个断面的浮游植物粒级结构相似, 微微型浮游植物的叶绿素a浓度贡献率最高, 均占了总叶绿素a浓度的 85%以上, 其次是微型浮游植物, 贡献最小的都是小型浮游植物, 贡献率不足 5%。100m以上水层, 微微型浮游植物叶绿素a的贡献率最高,随水深的增加, 其贡献率开始下降, 在 150—200m水层, 微型浮游植物叶绿素a的贡献率提高到 20%以上(表 1)。
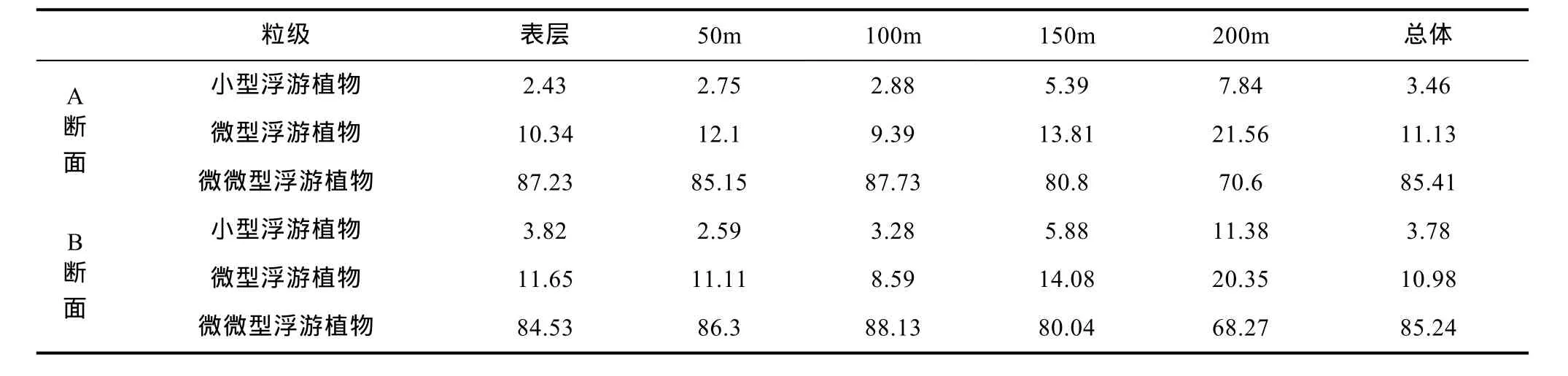
表1 M2海山不同水层分粒级叶绿素a浓度贡献率(%)Tab.1 Contribution (in percentage) of size-fractionated Chl a in different depths of every transection
2.3 M2海山初级生产力
M2海山的叶绿素同化系数为 1.3587mgC/(mg chla⋅h), 水柱初级生产力为 71.31mgC/(m2⋅d)。海山南部和北部初级生产力较高, 南部高于北部, 东部和西部生产力较低, 山顶处由于水深只有 37m, 没有出现叶绿素最大层, 因此初级生产力最低(图 4)。初级生产力组成主要为微型和微微型浮游植物, 其中微型浮游植物贡献率达到了 72%, 微微型浮游植物贡献率为28%, 小型浮游植物可以忽略不计。
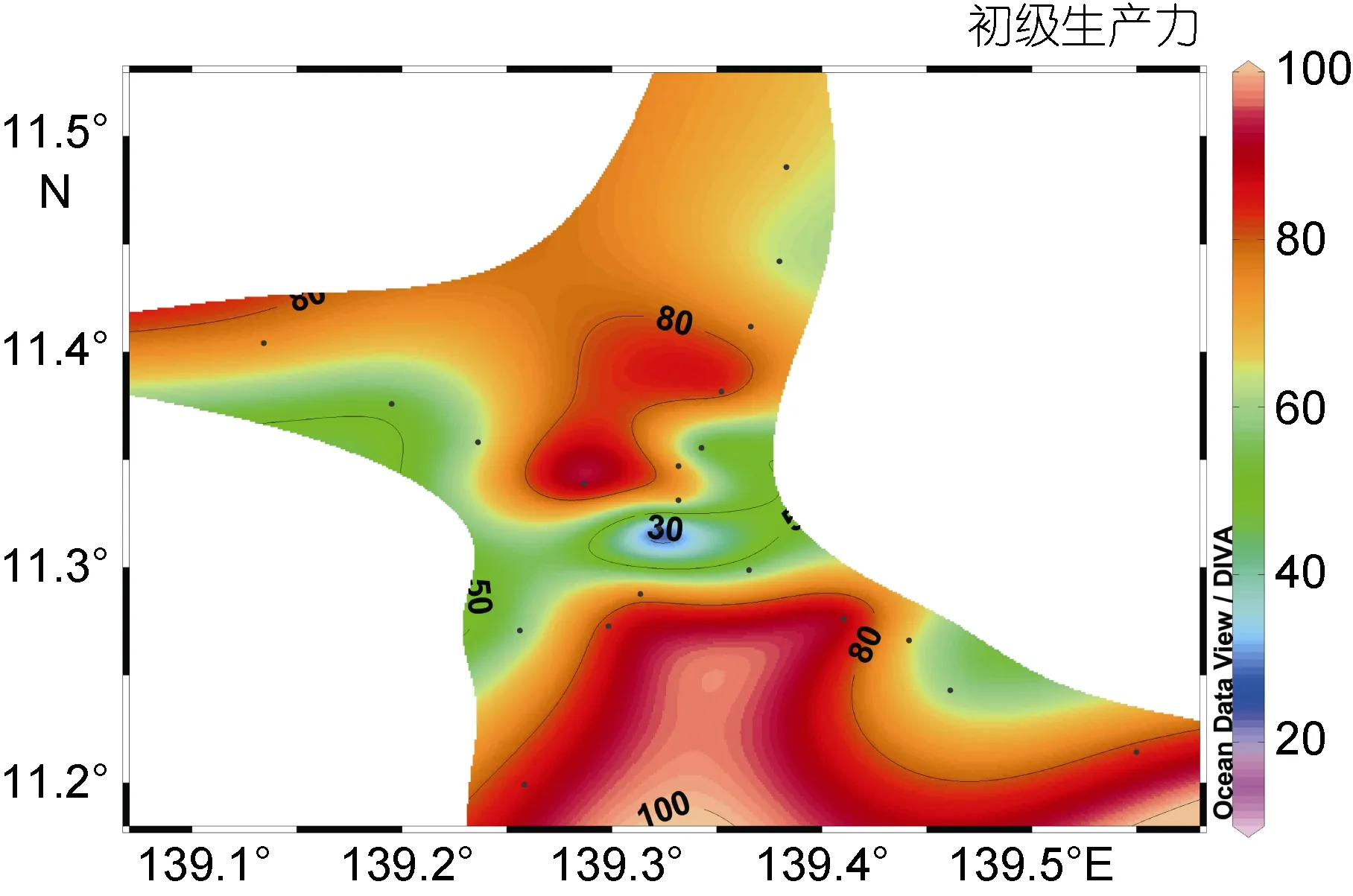
图4 M2海山区水柱初级生产力分布图(单位:mgC/(m2⋅d))Fig.4 The distribution of primary productivity in Seamount M2(mgC/(m2⋅d))
2.4 M2海山营养盐
由表2可以看出, M2海山的主要营养盐如磷酸盐、硝酸盐、硅酸盐等在75m及以上水层浓度较低,尤其是硝酸盐浓度在0m和15m水层不到1μg/L; 在DCML以下, 磷酸盐、硝酸盐和硅酸盐的的浓度开始随水深增加而升高。200m以上, 亚硝酸盐浓度均低于1μg/L, 氨氮在200m水深处浓度大约是150m以上水层的10倍。
3 讨论
3.1 M2海山区水文环境
M2海山区位于热带西太平洋暖池区, 表层水温常年在 28ºC以上, 存在严重的海水分层现象(Messiéet al, 2006), 从图2可以看出该海域温跃层较深, 接近 75m。北赤道流以20cm/s左右的流速自东向西流经M2海山(王凡等, 2001), 受到突起地形的影响, 上游的水流被压缩, 会导致气旋和反气旋漩涡的形成(Readet al, 2017), 进而在局部区域生成上升流, 使得水温较低、营养盐较为丰富的下层水上涌。如海山东北方向发现了等温线抬升的现象, 该现象暗示此处可能存在由涡引起的上升流等垂向运动物理过程。洋流在流经海山时, 和复杂的地形相互作用形成很多物理过程, 如内波、涡和泰勒柱等。Mouriño等(2001)在1992年的Great Meteor海山山顶处发现了泰勒柱现象, 但在 1999年对此海山的研究中没有发现到泰勒柱, 只观察到等温线抬升的现象, 等温线抬升的现象在多处海山的研究中均有发现(Comeauetal,1995; Cordeiroet al, 2013; 张文静等, 2016 )。
3.2 叶绿素a结构与分布特征
浮游植物的分布主要受温度、盐度、光照和营养盐等控制。对M2海山各站位的温度、盐度、硝酸盐、亚硝酸盐、磷酸盐、铵盐和硅酸盐进行主成分分析,前三个主成分的累计概率为91%, 结果如表3。其中第一主成分主要由温度、硝酸盐、磷酸盐和硅酸盐组成, 第二主成分主要由铵盐和亚硝酸盐组成, 第三部分主要由盐度和铵盐组成。其中深度直接影响温度,并间接通过温跃层影响营养盐的分布, 因此第一主成分主要代表了深度变化及其对温度和营养盐分布的影响; 第二主成分主要代表生物活动导致的铵盐和亚硝酸盐的变化; 第三主成分主要为洋流和密度分层引起的盐度变化。

表3 M2海山主要环境要素主成分分析结果Tab.3 The Principal Component Analysis of the major environment factors in Seamount M2
温度、光照和营养盐随深度变化, 进而影响浮游植物的垂直分布。本航次营养盐数据表明在 DCML及其以上的海水中各种营养盐的浓度都稳定在一个较低值, 尤其是硝酸盐浓度很低, 只有 10μg/L, 存在严重的营养盐限制; 在DCML以下, 主要营养盐的浓度随水深增加而增加。M2海山位于西太平洋暖池区,表层水温常年在 28ºC左右, 水体长期存在物理分层现象, 营养盐较为丰富的下层水无法与上层水进行交换, 上层水中营养盐成为浮游植物生长的限制因素, 并且该区域表层较强的光照会对浮游植物产生光抑制作用(李俊磊等, 2016), 在次表层, 浮游植物可以从下层水得到营养盐, 同时也可以得到适当的光照强度, 而真光层下方, 光照成为限制浮游植物生长的主要因素, 因此出现次表层叶绿素a浓度最大值的现象。洋流流经海山时产生的物理过程可以直接影响浮游植物的分布, 另外还可以打破原有稳定的分层, 使下层有较丰富营养盐、水温较低的海水上涌,从而促进了上层浮游植物的生长, 因此出现 DCML与等温线相一致的波动情况。这一现象在 Minami-Kasuga海山(Geninet al, 1985)及Komahashi No.2海山(Odateet al, 1998)也存在。
M2海山微微型浮游植物叶绿素a的贡献率达到了 85%, 稍高于 Zhang(2012)等人春季对该海区的调查, 与刘子琳等(2006)在 2002年春季黑潮源区的浮游植物粒级结构相吻合。浮游植物的结构和分布主要受营养盐结构和光照的影响。微微型浮游植物之所以占据绝对优势, 是因为调查区域所处的热带西太平洋海域是寡营养盐区, 表层营养盐尤其是硝酸盐浓度很低, 限制了小型和微型浮游植物的生长, 而微微型的聚球藻可以通过固氮对硝酸盐进行补偿, 一些微微型浮游植物可以适应深层较低光照强度(Mackeyet al, 2002), 如在南太平洋的调查中发现原绿球藻在1%光照强度下处于优势地位(DiTullioet al, 2003)。此次调查发现150—200m水层中, 随水深增加, 总叶绿素a浓度迅速下降, 但相对来说微型浮游植物贡献率有所提高(表 2)。Bouteiller等(1992)在该海区的研究结果表明, 该海域 100m以上水层, 微微型蓝藻为优势藻种, 100m以下水层, 聚球藻渐渐被微微型原绿球藻所取代, 整个真光层中微型金藻(主要为颗石藻)垂直分布较均匀, 该研究结果与此次调查结果相吻合。
3.3 初级生产力
该海区表层水体营养盐耗尽, 初级生产力受营养盐限制, 而DCML营养盐相对丰富, 初级生产力却受到光限制, 因此整体初级生产力水平较低。本次调查M2海山的平均初级生产力为71.31mgC/(m2⋅d), 钱宏林等(1986)和 Friedrichs等(2009)对该海域的生产力的估算结果为 50—100mgC/(m2⋅d), 与本次调查结果较为一致。Genin等(1985)和 Dower等(1992)等分别在Minamikasuga海山和Cobb海山山顶附近均发现了初级生产力升高的现象, 并由此提出了“海山效应”的经典假说(Doweret al, 1996), 但是此次对M2海山的调查结果并不支持该假说, Comeau等(1995), White等(2007), Young等(2011)和 Misic等(2012)对海山的调查中也没有发现明显的初级生产力升高的现象。
此次调查结果表明在M2海山附近, 虽然微微型浮游植物对叶绿素a浓度的贡献率达到了85%, 但是其对初级生产力的贡献率只有 30%左右, 初级生产力的主要贡献者为微型浮游植物。以往研究表明微微型浮游植物初级生产力的贡献率远远低于叶绿素a的贡献率, 尤其是在寡营养海区(Fernándezet al,2003), 如热带北大西洋寡营养水体的研究表明微微型浮游植物虽然占浮游植物总生物量的 75%, 但是却只贡献了一半左右的初级生产力(Fernándezet al,2003)。
3.4 M2海山与Y3海山比较
本次调查的M2海山与张文静等(2016)报道的雅浦Y3海山都位于热带西太平洋区域, 均发现了等温线波动的现象, 但是均未在山顶处发现泰勒柱存在。原因可能是地球自转偏向力在赤道附近很小, 不足以形成泰勒柱(Whiteet al, 2007), 也可能是该区域存在速率较快的向西流动的北赤道海流, 阻止了海山上形成稳定循环流动(Readet al, 2017)。
此次调查春季M2海山A、B断面平均叶绿素a浓度均为0.094mg/m3, 明显高于冬季Y3海山的0.057和 0.054mg/m3(张文静等, 2016)。这可能是两座海山的山顶距海平面深浅程度不同引起的差异: M2海山山顶深度仅为37m, 而Y3海山山顶深度接近300m。通常较浅的海山浮游植物生物量相对较高。Sonnekus等(2016)在西南印度洋的六座海山调查中发现, 浅的海山(深度<200m)对浮游植物的生物量有促进作用。较浅的M2海山可能在潮汐能和内波的作用下, 在海山山顶部存在流速加快的现象, 引起有机颗粒的再悬浮(Eriksen, 1991; Misicet al, 2012), 使得M2海山附近真光层中有较高的物质周转效率; 另外M2海山真光层中可能存在底栖群落与浮游群落的直接作用(Cordeiroet al, 2013), 也会加快物质的循环, 从而促进各粒级浮游植物的生长。对两座海山的对比研究中发现春季M2海山的表层盐度(深度<30m)显著高于雅浦Y3海山(df=51,P<0.05), M2海山DCML磷酸盐浓度极显著高于Y3海山(M2: 10.96μg/L, Y3: 7.67μg/L,df=29,P<0.01), M2海山DCML硅酸盐浓度显著高于Y3海 山 (M2: 91.75μg/L, Y3: 69.98μg/L, df=15,P<0.05), 可能是M2海山浮游植物叶绿素a浓度较高的直接原因。
M2海山微微型浮游植物对总叶绿素a的贡献率明显高于Y3海山。冬季雅浦Y3海山(张文静等, 2016)虽然也是微微型浮游植物具有优势, 但其对总叶绿素a的贡献率在A断面为60%左右, B断面只有50%左右。与雅浦 Y3海山的温度数据进行对比发现, 春季 M2海山的温跃层深度更深, 在 75m 附近, 与Zhang等(2012)和刘子琳等(2006)等人的调查相同,而Y3海山的温跃层在50m附近, 推测分层深度的增加, 会使得下层水的光照限制更加严重。M2海山的DCML多位于100m以深, 小型和微型浮游植物难以适应该水深的光照强度, 而微微型浮游植物如原绿球藻则能在较低的光照下生存, 因此微微型浮游植物在M2海山区贡献率更高。
Y3海山的初级生产力为 35.31mgC/(m2⋅d), 其中微型浮游植物贡献率最高, 接近80%, 微微型浮游植物贡献率约为 17%(Y3海山未发表数据)。以往研究表明, 较大的浮游植物(>2μm)拥有更高的光合作用效率, 尤其是在寡营养海区(Fernándezet al, 2003),与 M2和 Y3调查结果相吻合。M2海山的初级生产力约为Y3海山的2倍, 两座海山的对比结果表明, 浮游植物在较浅的海山有更高的生物量和初级生产力。
4 结论
(1) M2海山存在物理过程引起的等温线波动的现象, 该过程促进了下层水和上层水的垂直交换, 进而影响浮游植物的分布, 导致DCML的波动。
(2) M2海山微微型浮游植物叶绿素a对总叶绿素a的贡献最大, 达到了 85%以上; 初级生产力为71.31mgC/(m2⋅d), 其中微型浮游植物对总初级生产力的贡献最高, 为72%。
(3) M2海山的海山效应不明显。
(4) M2海山的叶绿素a浓度和初级生产力接近Y3海山两倍, 浅的海山(<200)可能对浮游植物生物量起促进作用。M2海山微微型浮游植物的贡献率更高, 两座海山粒级结构的差异主要是由温跃层深度决定的。
(5) 目前针对西太平洋海山的浮游生态学研究较少, 建议增加不同季节的航次, 对不同深度的海山进行系统研究。
致谢本研究部分样品采集由本课题组成员陈芸燕、张文静、李俊磊和孙玉娥等人完成, 温盐数据由中国科学院海洋研究所海洋专项办公室提供, 营养盐数据由李学刚老师提供, 此次海上调查得到了“科学号”考察船全体工作人员的支持, 谨致谢忱。
王 凡, 张 平, 胡敦欣等, 2001. 热带西太平洋环流及其季节变化. 科学通报, 46(23): 1998—2002
刘子琳, 陈忠元, 周蓓锋等, 2006. 黑潮源区及其邻近海域叶绿素a浓度的季节分布. 海洋湖沼通报, (1): 58—63
李俊磊, 孙晓霞, 张文静等, 2016. 2014年冬季西太平洋浮游植物光合作用特征. 海洋与湖沼, 47(5): 935—944
张文静, 孙晓霞, 陈芸燕等, 2016. 2014年冬季热带西太平洋雅浦Y3海山浮游植物叶绿素a浓度及粒级结构. 海洋与湖沼, 47(4): 739—747
张均龙, 徐奎栋, 2013. 海山生物多样性研究进展与展望.地球科学进展, 28(11): 1209—121
钱宏林, 齐雨藻, 1992. 1986年11—12月热带西太平洋初级生产力的估算. 暨南大学学报(自然科学), 13(3): 124—129
Clark M R, Rowden A A, Schlacher Tet al, 2010. The ecology of seamounts: structure, function, and human impacts. Annual Review of Marine Science, 2: 253—278
Clark M R, Schlacher T A, Rowden A Aet al, 2012. Science priorities for seamounts: research links to conservation and management. PLoS One, 7(1): e29232
Comeau L A, Vézina A F, Bourgeois Met al, 1995. Relationship between phytoplankton production and the physical structure of the water column near Cobb Seamount, Northeast Pacific.Deep Sea Research Part I: Oceanographic Research Papers,42(6): 993—1005
Cordeiro T A, Brandini F P, Rosa R Set al, 2013. Deep chlorophyll maximum in western equatorial Atlantic——How does it interact with islands slopes and seamounts?.Marine Science, 3(1): 30—37
DiTullio G R, Geesey M E, Jones D Ret al, 2003. Phytoplankton assemblage structure and primary productivity along 170°W in the South Pacific Ocean. Marine Ecology-Progress Series,255: 55—80
Djurhuus A, Read J F, Rogers A D, 2017. The spatial distribution of particulate organic carbon and microorganisms on seamounts of the South West Indian Ridge. Deep Sea Research Part II: Topical Studies in Oceanography, 136:73—84
Dower J F, Mackas D L, 1996. “Seamount effects” in the zooplankton community near Cobb Seamount. Deep Sea Research Part I: Oceanographic Research Papers, 43(6):837—858
Dower J, Freeland H, Juniper K, 1992. A strong biological response to oceanic flow past Cobb seamount. Deep Sea Research Part A. Oceanographic Research Papers, 39(7—8):1139—1145
Epp D, Smoot N C, 1989. Distribution of seamounts in the North-Atlantic. Nature, 337(6204): 254—257
Eriksen C C, 1991. Observations of amplified flows atop a large seamount. Journal of Geophysical Research Oceans, 96(C8):15227—15236
Fernández E, Marañón E, Morán X A Get al, 2003. Potential causes for the unequal contribution of picophytoplankton to total biomass and productivity in oligotrophic waters.Marine Ecology Progress, 254: 101—109
Friedrichs M A M, Carr M-E, Barber R Tet al, 2009. Assessing the uncertainties of model estimates of primary productivity in the tropical Pacific Ocean. Journal of Marine Systems,76(1—2): 113—133
Genin A, Boehlert G W, 1985. Dynamics of temperature and chlorophyll structures above a seamount: an oceanic experiment. Journal of Marine Research, 43(4): 907—924
Le Bouteiller A, Blanchot J, Rodier M, 1992. Size distribution patterns of phytoplankton in the western Pacific: towards a generalization for the tropical open ocean. Deep Sea Research Part A. Oceanographic Research Papers, 39(5):805—823
Letessier T B, De Grave S, Boersch-Supan P Het al, 2017.Seamount influences on mid-water shrimps (Decapoda) and gnathophausiids (Lophogastridea) of the south-west Indian ridge. Deep Sea Research Part II: Topical Studies in Oceanography, 136: 85—97
Mackey D J, Blanchot J, Higgins H Wet al, 2002. Phytoplankton abundances and community structure in the equatorial Pacific. Deep Sea Research Part II: Topical Studies in Oceanography, 49(13—14): 2561—2582
Messié M, Radenac M-H, 2006. Seasonal variability of the surface chlorophyll in the western tropical Pacific from SeaWiFS data. Deep Sea Research Part I: Oceanographic Research Papers, 53(10): 1581—1600
Misic C, Bavestrello G, Bo Met al, 2012. The “seamount effect”as revealed by organic matter dynamics around a shallow seamount in the Tyrrhenian Sea (Vercelli Seamount, western Mediterranean). Deep Sea Research Part I: Oceanographic Research Papers, 67: 1—11
Mouriño B, Fernández E, Serret Pet al, 2001. Variability and seasonality of physical and biological fields at the Great Meteor Tablemount (subtropical NE Atlantic). Oceanologica Acta, 24(2): 167—185
Odate T, Furuya K, 1998. Well-developed subsurface chlorophyll maximum near Komahashi No.2 Seamount in the summer of 1991. Deep Sea Research Part I: Oceanographic Research Papers, 45(10): 1595—1607
Read J, Pollard R, 2017. An introduction to the physical oceanography of six seamounts in the southwest Indian Ocean. Deep Sea Research Part II: Topical Studies in Oceanography, 136: 44—58
Sonnekus M J, Bornman T G, Campbell E E, 2016.Phytoplankton and nutrient dynamics of six South West Indian Ocean seamounts. Deep Sea Research Part II: Topical Studies in Oceanography, 136: 59—72
White M, Bashmachnikov I, Arístegui Jet al, 2007. Physical Processes and Seamount Productivity. In: Pitcher T J,Morato T, Hart P J Bet al, eds. Seamounts: Ecology,Fisheries & Conservation. Blackwell, Oxford, UK: Fish and Aquatic Resources Series, 62—84
Young J W, Hobday A J, Campbell R Aet al, 2011. The biological oceanography of the East Australian Current and surrounding waters in relation to tuna and billfish catches off eastern Australia. Deep Sea Research Part II: Topical Studies in Oceanography, 58(5): 720—733
Zhang D S, Wang C S, Liu Z Set al, 2012. Spatial and temporal variability and size fractionation of chlorophyllain the tropical and subtropical pacific ocean. Acta Oceanologica Sinica, 31(3): 120—131
猜你喜欢
南方水产科学(2021年3期)2021-06-23
Plasma Science and Technology(2020年9期)2020-09-14
37°女人(2019年4期)2019-04-20
吉林农业(2018年15期)2018-08-14
作品(2016年10期)2016-12-06
大江南北(2016年3期)2016-11-22
应用海洋学学报(2015年3期)2015-11-22
应用海洋学学报(2015年1期)2015-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
天然气勘探与开发(2014年4期)2014-02-28
