
Novel insights into the role of NF-κB p50 in astrocytemediated fate specification of adult neural progenitor cells
2017-04-07 03:36
中国神经再生研究(英文版) 2017年3期
Laboratory of Neuroplasticity, Department of Pharmaceutical Sciences, University of Piemonte Orientale, Novara, Italy
Novel insights into the role of NF-κB p50 in astrocytemediated fate specification of adult neural progenitor cells
Valeria Bortolotto, Mariagrazia Grilli*
Laboratory of Neuroplasticity, Department of Pharmaceutical Sciences, University of Piemonte Orientale, Novara, Italy
How to cite this article:Bortolotto V, Grilli M (2017) Novel insights into the role of NF-κB p50 in astrocyte-mediated fate specification of adult neural progenitor cells. Neural Regen Res 12(3):354-357.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
Within the CNS nuclear factor-kappa B (NF-κB) transcription factors are involved in a wide range of functions both in homeostasis and in pathology. Over the years, our and other groups produced a vast array of information on the complex involvement of NF-κB proteins in different aspects of postnatal neurogenesis. In particular, several extracellular signals and membrane receptors have been identified as being able to affect neural progenitor cells (NPC) and their progenyviaNF-κB activation. A crucial role in the regulation of neuronal fate specification in adult hippocampal NPC is played by the NF-κB p50 subunit. NF-κB p50KO mice display a remarkable reduction in adult hippocampal neurogenesis which correlates with a selective defect in hippocampal-dependent short-term memory. Moreover absence of NF-κB p50 can profoundly affect thein vitroproneurogenic response of adult hippocampal NPC (ahNPC) to several endogenous signals and drugs. Herein we briefly review the current knowledge on the pivotal role of NF-κB p50 in the regulation of adult hippocampal neurogenesis. In addition we discuss more recent data that further extend the relevance of NF-κB p50 to novel astroglia-derived signals which can influence neuronal specification of ahNPC and to astrocyte-NPC cross-talk.
adult neurogenesis; hippocampus; astrocyte; neural progenitor cell; NGAL/LCN-2; conditioned medium; 24p3R
Introduction
The generation of new neurons in adulthood, referred as to adult neurogenesis, occurs in restricted areas of mammalian brain, including the subgranular zone (SGZ) in the dentat gyrus (DG) of the hippocampus. In this region residen neural stem/progenitor cells (NSC/NPC) are present, can self-renew and give rise to nestin-positive transiently amplifying progenitor cells which, in turn, can generate neuroblasts capable of terminal neuronal differentiation. Finally the post-mitotic neuronal progeny of GC can be functionally integrated into the adult hippocampal circuitry (Bond e al., 2015; Kempermann et al., 2015). Adult hippocampa neurogenesis (ahNG) has recently attracted interest due to its involvement in important functions such as cognition and memory, mood and emotional behavior, stress respons (Aimone et al., 2010, 2014; Eisch and Petrik, 2012; Bortolotto et al., 2014). In addition to its potential role in brain homeostasis, ahNG is also profoundly dysregulated in severa neuropsychiatric/neurodegenerative opening the question whether this process may participate in the pathophysiology of such diseases or, at least, contribute to some of their associated symptoms, such as dementia and depressed mood (Grilli and Meneghini, 2012; Bortolotto and Grilli, 2016a Yun et al., 2016). Based on such findings, there is a need fo a better understanding of the molecular pathways and receptors which are responsible for modulation of ahNG both under physiological and pathological conditions.
NF-κB Signaling Pathway in the Regulation of ahNG
Within the CNS, nuclear factor-kappa B (NF-κB) transcription factors have been involved in a wide range of functions both in homeostasis and in pathology (Grilli et al., 1996; Grilli and Memo, 1997; Grilli and Memo, 1999; Kaltschmidt and Kaltschmidt, 2009; Bonini et al., 2016). NF-κB proteins are also highly expressed in areas of active neurogenesis in the postnatal and adult mouse brain (Denis-Donini et al., 2005; Meneghini et al., 2013). Over the years, our and other groups have produced a vast array of information on the complex involvement of NF-κB proteins in different aspects of postnatal neurogenesis (Grilli and Meneghini, 2012; Bortolotto and Grilli, 2014). In particular, several extracellular signals and membrane receptors have been identified as being able to affect neural stem cells/neural progenitor cells (NSC/NPC) and their progenyviaNF-κB activation (Meneghini et al., 2010, 2013; Valente et al., 2015; Cuccurazzu et al., 2013; Bortolotto and Grilli, 2016a, b).
Since the initial observation of NF-κB presence in adult neurogenic areas, our group further explored the role of these regulatory proteins in adult neurogenesis with a spe-cific focus on the NF-κB1 (p50) subunit. The availability of p50KO mice and their wild type counterpart (WT) allowed us to demonstrate that absence of p50 can profoundly affect thein vitroresponse of adult hippocampal NPC (ahNPC) to several endogenous signals and to drugs (Valente et al., 2012, 2015; Meneghini et al., 2013; Nafez et al., 2015). In addition to that, p50KO mice display a remarkable reduction in adult hippocampal neurogenesis which correlates with a selective defect in hippocampal-dependent short-term memory (Denis-Donini et al., 2008). A phenotypic characterization of the mutant mouse model suggested that lack of p50 results in defects in the late maturation of newly generated neurons, in absence of significant alterations in survival and proliferation rates of ahNPC (Denis-Donini et al., 2008). Interestingly, when culturedin vitro, ahNPC from WT and p50KO mice do not display different neurogenic potential, suggesting a role of the neurogenic niche in defective hippocampal neurogenesis observed in mutant micein vivo.
The Cross-Talk Between Astrocytes and ahNPC in the Modulation of Adult Neurogenesis: A Role for NF-κB p50
It is well established that homeostasis of adult neurogenesis requires permissive and instructive signals for ahNPC, including the ones from the local microenvironment, the so called “neurogenic niche”. A few years ago, a pivotal work demonstrated that astroglial cells, an important component of the neurogenic niche, can indeed instruct stem/progenitor cells to adopt a neuronal fate (Song et al., 2002). Since then the role of astrocyte-mediated signaling in ahNPC regulation has been extensively confirmed.
Recently we identified a novel astrocyte-secreted molecule which can affect cell fate specification of neural progenitors. We wanted to dissect the role of NF-κB p50 in the cross-talk between adult neural progenitor cells and astrocytes. In particular, we aimed at understanding the potential influence of astroglia-generated soluble factors on aNPC fate specification and the cell autonomous or non cell-autonomous role played by NF-κB p50 signaling in that context. To this purpose, we exposed both WT and p50KO hippocampal NPC to conditioned media from WT and p50KO astrocytes. These efforts allowed us to suggest that dysregulated NPC-astrocyte communication may indeed contribute to reduced hippocampal neurogenesis in p50KO micein vivo(Cvijetic et al., 2017).
Lack of NF-κB p50 and Its Effect on Astrocyte Secretory Profile
In line with previously published data, WT Astrocyte Conditioned Medium (ACM) tested on WT ahNPC promoted their neuronal differentiation. When grown in culture in absence of growth factors, ahNPC spontaneously differentiate not only toward the neuronal but also toward non neuronal lineages (Meneghini et al., 2014). When ahNPC were exposed to p50KO ACM, astrogliogenic but not proneurogenic effects could be observed (Cvijetic et al., 2017). The lack of proneurogenic effects elicited by p50KO ACM could be ascribed to alterations in the secretory profile of astrocytes, occurring in absence of the NF-κB subunit. In particular, primary astrocytes lacking p50 could either become defective in the production of pro-neurogenic molecule(s) or, alternatively, produce antineurogenic molecule(s). These changes may occur because in absence of p50 a profound rearrangement occurs in the available pool of NF-κB transcriptional dimeric complexes, which may in turn result in activation and/or repression of target gene(s) (Grilli and Memo, 1999). Another possibility, that at present we cannot exclude, is that p50 absence may affect astrocyte secretory profile by altering the expression of genes encoding proteins which may influence the activity of secreted proteinsviaposttranscriptional modifications,i.e., kinases, phosphatases, proteases.
We decided to actively search for proteins differentially secreted by WT and p50KO astrocytes and correlated with the lack of pro-neurogenic effects observed in p50KO ACM. A proteomic approach allowed us to identify two soluble molecules which are upregulated in p50KO compared to WT ACM, namely C-C motif chemochine-2 (CCL2) and neutrophil gelatinase-associated lipocalin/lipocalin-2 (NGAL/LCN-2).
NGAL/LCN-2 as a Novel Proneurogenic Astrocyte-Derived Factor
Absence of NF-κB p50 is commonly linked to inflammation in brain (Bernal et al., 2014; Bortolotto et al., 2014; Rolova et al., 2014) and both CCL-2 and NGAL/LCN-2, which are encoded by NF-κB target genes, share an established role in inflammatory processes (Ueda et al., 1994; Kohda et al., 2014). Since no information was yet available on the role of NGAL/ LCN-2 on adult neurogenesis, we decided to concentrate our attention on this protein, whose expression is increased by about threefold in p50KO compared to WT ACM. NGAL/ LCN-2 is an iron-related protein whose modulatory role in the CNS is not completely understood, but it is commonly regarded as a deleterious signal (Ferreira et al., 2015). To our surprise we discovered that NGAL/LCN-2 can promote, in a concentration-dependent manner, neuronal differentiation of WT ahNPC. Interestingly, under the same experimental conditions, NGAL/LCN-2 had no or little effect on neuronal differentiation of p50KO NPC. Altogether, these data propose NGAL/LCN-2 as a novel astroglial-derived signal able to promote neuronal fate specification of ahNPC (Figure 1). We also discovered that p50KO NPC are less responsive to NGAL/LCN-2 and we could correlate such reduced sensitivity with downregulated expression of the LCN-2 receptor 24p3R, in mutant ahNPC. Although it has been demonstrated that 24p3R is constitutively expressed at high levels in brain (Chia et al., 2015), including the hippocampus, this is the first demonstration that the receptor is functionally expressed also in ahNPC.
Cell Autonomous Changes Also Occur in ahNPC in Absence of NF-κB p50
The reduced expression of 24p3R in p50KO NPC prompted us to further investigate the phenotypic changes that mayalso occur in absence of p50 in adult neural progenitors. We therefore tested the effect of WT and p50KO ACM also on p50KO NPC differentiation. Surprisingly, neither WT nor p50KO ACM promoted neuronal and also astroglial differentiation of p50KO NPC over basal conditions. These findings suggest that, in absence of the NF-κB p50 subunit, cell autonomous changes may also occur and affect responsiveness of ahNPC to astrocyte-derived pro-neurogenic and astrogliogenic signals (Cvijetic et al., 2017).
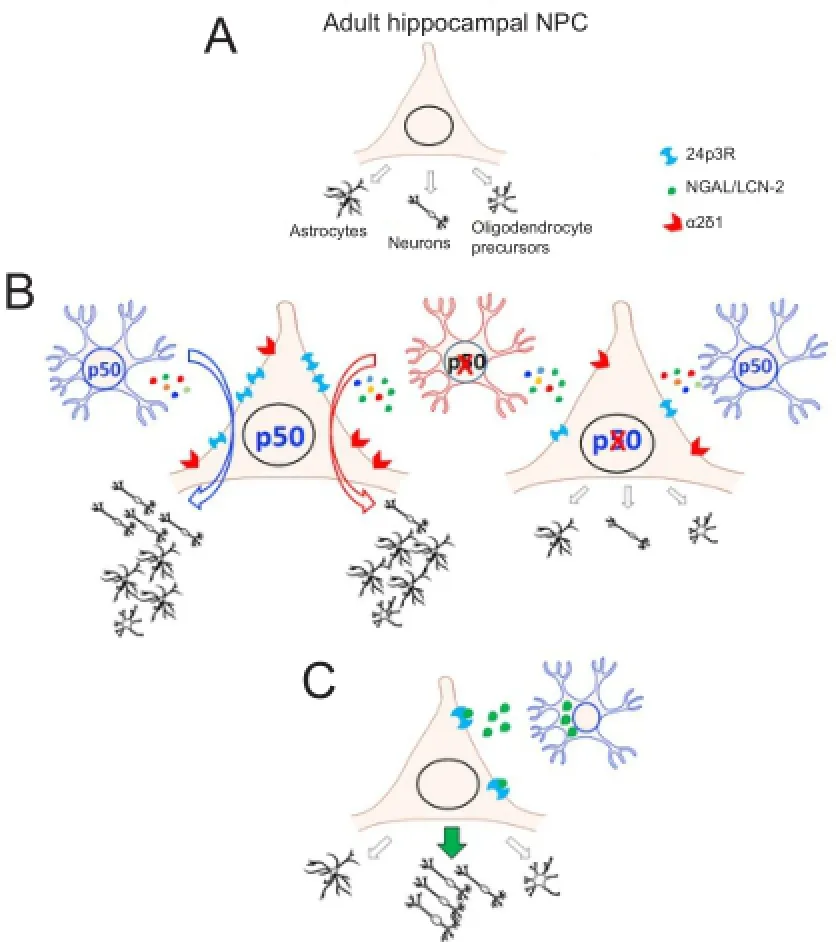
Figure 1 Cell autonomous and non cell-autonomous role of nuclear factor-kappa B (NF-κB) p50 in astrocyte-mediated fate specification of adult hippocampal neural progenitor cells.
While searching for phenotypic changes in p50KO NPC which may explain their reduced responsiveness to astrocyte-derived pro-neurogenic signals we evaluated the possibility that mutant cells may be deficient not only in 24p3R but also in other membrane-associated receptors for astrocyte-derived pro-neurogenic molecules. RAGE, the Receptor for Advanced Glycation End-products, is functionally expressed by adult NPC where it mediates the pro-neurogenic effects of several molecules, like HMGB-1 and S100B, which are known to be secreted by astrocytes (Meneghini et al., 2013; Bortolotto and Grilli, 2016a, b). Since in presence of a neutralizing anti-RAGE antibody WT ACM pro-neurogenic effects were unaffected, we could exclude the involvement of this receptor in astrocyte-mediated neuronal differentiation of ahNPC.
Trombospondin-1 (TSP-1) is another established astrocyte-derived pro-neurogenic factor (Lu and Kipnis, 2010). The voltage-gated calcium channel α2δ1 subunit has been demonstrated to be a TSP-1 receptor (Eroglu et al., 2009) which is functionally expressed also by ahNPC (Valente et al., 2012). Interestingly, we discovered that α2δ1 expression levels were also significantly reduced in p50KO ahNPC, compared to their WT counterpart. In agreement with this observation, TSP-1 promoted an increase in the percentage of newly formed neurons in WT, but not in p50KO, NPC cultures. Altogether these data suggested that, in absence of p50, ahNPC responsiveness to distinct astrocyte-derived pro-neurogenic signals, and in particular to TSP-1 and LCN-2, was reducedviadownregulated expression of their membrane receptors (Cvijetic et al., 2017).
Conclusions and Perspectives
At present we have not yet identified a culprit for reduced pro-neurogenic effects of p50KO ACM on WT NPC. Indeed, in our cellular model also CCL-2, the other protein whose expression is significantly upregulated in p50KO ACM, exerted pro-neurogenic activity on WT ahNPC (unpublished data). Of course it is possible that LC-MS/MS analysis of ACM identified only a fraction of all astrocyte-secreted proteins, so we may have missed other contributors to the reduced proneurogenic potential of p50KO ACM.
In our experimental setting p50KO NPC were unresponsive not only to pro-neurogenic but also to astrogliogenic astrocyte-derived signals. In the future efforts will be devoted to identify astrocyte-secreted signals that promote astrogliogenesis from ahNPC as well as additional cell-autonomous changes that may take place in progenitors in absence of the NF-κB p50 subunit. Of course we are aware that the inability of p50KO NPC to respond to both pro-neurogenic and astrogliogenic signals may also depend on p50-requirement in the downstream signaling pathway triggered by exposure of NPC to ACM treatment. For example, NF-κB signaling activation has also been shown to occur downstream of LCN-2/24p3R activation (Ferreira et al., 2015).
Altogether based on our recent data, it can be envisioned that reduced hippocampal neurogenesis in p50KO mice (Denis-Donini et al., 2008) could be ascribed to complex defects in the cross-talk between astrocytes and ahNPC. Additionally, since microglial cells are well known contributors to hippocampal NPC specification we cannot exclude their contribution in the reduction of hippocampal neurogenesis observed in p50KO mice.
Our experimental work demonstrates that complex cell autonomous and non cell-autonomous changes can affect NPC fate specification in absence of NF-κB p50, as summarized inFigure 1. We also showed, for the first time, that LCN-2 is an astroglia-derived signal which promotes neuronal fate specification of ahNPC and whose activity, in parallel with a downregulation of 24p3R, is strongly reduced in p50KO NPC.
Altogether, these data add further complexity to a growing body of data suggesting the relevance of NF-κB signaling in the modulation of adult hippocampal neurogenesis and the key role of astrocytes in the neurogenic niche.
Acknowledgments:This work was supported by a grant from Fondazione Cariplo to MG.
Author contributions:Both authors wrote the review.
Conflicts of interest:None declared.
Aimone JB, Deng W, Gage FH (2010) Adult neurogenesis: integrating theories and separating functions. Trends Cogn Sci 14:325-337.
Aimone JB, Li Y, Lee SW, Clemenson GD, Deng W, Gage FH (2014) Regulation and function of adult neurogenesis: from genes to cognition. Physiol Rev 94:991-1026.
Bernal GM, Wahlstrom JS, Crawley CD, Cahill KE, Pytel P, Liang H, Kang S, Weichselbaum RR, Yamini B (2014) Loss of NFκB1 leads to early onset aging. Aging 6:931-943.
Bond AM, Ming GL, Song H (2015) Adult mammalian stem cells and neurogenesis: five decades later. Cell Stem Cell 17:385-395.
Bonini SA, Mastinu A, Maccarinelli G, Mitola S, Premoli M, La Rosa LR, Ferrari-Toninelli G, Grilli M, Memo M (2016) Cortical structure alterations and social behavior impairment in p50-deficient mice. Cereb Cortex 26:2832-2849.
Bortolotto V, Cuccurazzu B, Canonico PL, Grilli M (2014) NF-κB mediated regulation of adult hippocampal neurogenesis: relevance to mood disorders and antidepressant activity. BioMed Res Int 2014:612798.
Bortolotto V, Grilli M (2016a) Every cloud has a silver lining: Proneurogenic effects of Aβ oligomers and HMGB-1 via activation of the RAGE-NF-κB axis. CNS Neurol Disord Drug Targets doi:10.2174/18 71527315666160803153459.
Bortolotto V, Grilli M (2016b) Not only a bad guy: potential proneurogenic role of the RAGE/ NF-κB axis in Alzheimer’s disease brain. Neural Regen Res 11:1924-1925.
Chia WJ, Tan FC, Ong WY, Dawe GS (2015) Expression and localization of brain-type organic cation transporter (BOCT/24p3R/LCN2R) in the normal rat hippocampus and after kainate-induced toxicity. Neurochem Int 87:43-59.
Cuccurazzu B, Bortolotto V, Valente MM, Ubezio F, Koverech A, Canonico PL, Grilli M (2013) Upregulation of mGlu2 receptors via NF-κB p65 acetylation is involved in the proneurogenic and antidepressant effects of acetyl-l-carnitine. Neuropsychopharmacology 38:2220-2230.
Cvijetic S, Bortolotto V, Manfredi M, Ranzato E, Marengo E, Salem R, Canonico PL, Grilli M (2017) Cell autonomous and noncell-autonomous role of NF-κB p50 in astrocyte-mediated fate specification of adult neural progenitor cells. Glia 65:169-181.
Denis-Donini S, Caprini A, Frassoni C, Grilli M (2005) Members of the NF-kappaB family expressed in zones of active neurogenesis in the postnatal and adult mouse brain. Brain Res Dev Brain Res 154:81-89.
Denis-Donini S, Dellarole A, Crociara P, Francese MT, Bortolotto V, Quadrato G, Canonico PL, Orsetti M, Ghi P, Memo M, Bonini SA, Ferrari-Toninelli G, Grilli M (2008) Impaired adult neurogenesis associated with short-term memory defects in NF-kappaB p50-deficient mice. J Neurosci 28:3911-3919.
Eisch AJ, Petrik D (2012) Depression and hippocampal neurogenesis: a road to remission? Science 338:72-75.
Eroglu C, Allen NJ, Susman MW, O’Rourke NA, Park CY, Ozkan E, Chakraborty C, Mulinyawe SB, Annis DS, Huberman AD, Green EM, Lawler J, Dolmetsch R, Garcia KC, Smith SJ, Luo ZD, Rosenthal A, Mosher DF, Barres BA (2009) Gabapentin receptor alpha2delta-1 is a neuronal thrombospondin receptor responsible for excitatory CNS synaptogenesis. Cell 139:380-392.
Ferreira AC, Dá Mesquita S, Sousa JC, Correia-Neves M, Sousa N, Palha JA, Marques F (2015) From the periphery to the brain: Lipocalin-2, friend or foe? Prog Neurobiol 131:120-136.
Grilli M, Memo M (1997) Transcriptional pharmacology of neurodegenerative disorders: novel venue towards neuroprotection against excitotoxicity? Mol Psychiatry 2:192-194.
Grilli M, Memo M (1999) Nuclear Factor-κB/Rel proteins: a point of convergence of signalling pathways relevant in neuronal function and dysfunction. Biochem Pharmacol 57: 1-7.
Grilli M, Meneghini V (2012) NF-κB proteins in adult neurogenesis: relevance for learning and memory in physiology and pathology. In: Transcription factors CREB and NF-κB: involvement in synaptic plasticity and memory formation (Albensi BC, Oak Park IL, eds), pp199-205. Bentham Science Publishers Ltd.
Grilli M, Pizzi M, Memo M, Spano PF (1996) Neuroprotection by aspirin and sodium salicylate through blockade of NF-κB activation. Science 274: 1383-1385.
Kaltschmidt B, Kaltschmidt C (2009) NF-kappaB in the nervous system. Cold Spring Harb Perspect Biol 1:a001271.
Kempermann G, Song H, Gage FH (2015) Neurogenesis in the hippocampus. Cold Spring Harb Perspect Biol 7:a01812.
Kohda A, Yamazaki S, Sumimoto H (2014) DNA element downstream of the κB site in the Lcn2 promoter is required for transcriptional activation by IκBζ and NF-κB p50. Genes Cells 19:620-628.
Lu Z, Kipnis J (2010) Trombospondin 1-A key astrocyte-derived neurogenic factor. FASEB J 24:1925-1934.
Meneghini V, Francese MT, Carraro L, Grilli M (2010) A novel role for the receptor for advanced glycation end-products in neural progenitor cells derived from adult subventricular zone. Mol Cell Neurosci 45:139-150.
Meneghini V, Bortolotto V, Francese MT, Dellarole A, Carraro L, Terzieva S, Grilli M (2013) High-mobility group box-1 protein and β-amyloid oligomers promote neuronal differentiation of adult hippocampal neural progenitors via receptor for advanced glycation end products/nuclear factor-κB axis: relevance for Alzheimer’s disease. J Neurosci 33:6047-6059.
Meneghini V, Cuccurazzu B, Bortolotto V, Ramazzotti V, Ubezio F, Tzschentke TM, Canonico PL, Grilli M (2014) The noradrenergic component in tapentadol action counteracts μ-opioid receptor-mediated adverse effects on adult neurogenesis. Mol Pharmacol 85:658-670.
Nafez S, Oikawa K, Odero GL, Sproule M, Ge N, Schapansky J, Abrenica B, Hatherell A, Cadonic C, Zhang S, Song X, Kauppinen T, Glazner GW, Grilli M, Czubryt MP, Eisenstat DD, Albensi BC (2015) Early growth response 2 (Egr-2) expression is triggered by NF-κB activation. Mol Cell Neurosci 64:95-103.
Rolova T, Puli L, Magga J, Dhungana H, Kanninen K, Wojciehowski S, Salminen A, Tanila H, Koistinaho J, Malm T (2014) Complex regulation of acute and chronic neuroinflammatory responses in mouse models deficient for nuclear factor kappa B p50 subunit. Neurobiol Dis 64:16-29.
Song H, Stevens CF, Gage FH (2002) Astroglia induce neurogenesis from adult neural stem cells. Nature 417:39-44.
Ueda A, Okuda K, Ohno S, Shirai A, Igarashi T, Matsunaga K, Fukushima J, Kawamoto S, Ishigatsubo Y, Okubo T (1994) NF-kappaB and Sp1 regulate transcription of human monocyte chemoattractant protein-1 gene. J Immunol 153:2025-2063.
Valente MM, Allen M, Bortolotto V, Lim ST, Conant K, Grilli M (2015) The MMP-1/PAR-1 axis enhances proliferation and neuronal differentiation of adult hippocampal neural progenitor cells. Neural Plast 2015:646595.
Valente MM, Bortolotto V, Cuccurazzu B, Ubezio F, Meneghini V, Francese MT, Canonico PL, Grilli M (2012) α2δ Ligands act as positive modulators of adult hippocampal neurogenesis and prevent depressive-like behavior induced by chronic restraint stress. Mol Pharmacol 82:271-280.
Yun S, Reynolds RP, Masiulis I, Eisch AJ (2016) Re-evaluating the link between neuropsychiatric disorders and dysregulated adult neurogenesis. Nat Med 22:1239-1247.
*Correspondence to: Mariagrazia Grilli, M.D., mariagrazia.grilli@uniupo.it.
orcid: 0000-0001-9165-5827 (Mariagrazia Grilli)
10.4103/1673-5374.202919
Accepted: 2017-02-14

- 中国神经再生研究(英文版)的其它文章
- Tissue-type plasminogen activator is a homeostatic regulator of synaptic function in the central nervous system
- The emerging role of autophagic-lysosomal dysfunction in Gaucher disease and Parkinson’s disease
- Neuromelanin, one of the most overlooked molecules in modern medicine, is not a spectator
- Telomerase and mTOR in the brain: the mitochondria connection
- Anesthetic considerations for patients with acute cervical spinal cord injury
- Impacts of the retinal environment and photoreceptor type on functional regeneration