
贵州省东方蜜蜂微卫星DNA遗传分化与遗传多样性分析
2017-06-15 18:15于瀛龙周姝婧徐新建朱翔杰杨凯杰陈道印周冰峰
福建农林大学学报(自然科学版) 2017年3期
于瀛龙, 周姝婧, 徐新建, 朱翔杰, 杨凯杰, 陈道印, 周冰峰
(福建农林大学蜂学学院,福建 福州 350002)
贵州省东方蜜蜂微卫星DNA遗传分化与遗传多样性分析
于瀛龙, 周姝婧, 徐新建, 朱翔杰, 杨凯杰, 陈道印, 周冰峰
(福建农林大学蜂学学院,福建 福州 350002)
利用31个微卫星位点对贵州全省的东方蜜蜂进行分析.结果表明:贵州省东方蜜蜂遗传分化系数(FST)为0~0.03,没有发现贵州省东方蜜蜂发生遗传分化.贵州省东方蜜蜂微卫星平均期望杂合度为0.434 5±0.055 0,平均观察杂合度为0.419 6±0.011 4,平均等位基因数为5.56±4.34,平均有效等位基因数为2.693 0±2.113 5,平均香农指数为0.873 5±0.797 1.贵州省东方蜜蜂微卫星遗传多样性高于全国东方蜜蜂的平均水平.本文研究结果对了解我国东方蜜蜂遗传结构、遗传分化和遗传多样性水平具有重要的价值,对指导贵州省东方蜜蜂遗传资源保护与利用提供理论依据.
东方蜜蜂; 遗传分化; 遗传多样性; 微卫星DNA
遗传多样性是衡量物种适应外界环境变化能力的重要指标,能够反映物种的进化潜力,同时也是遗传资源领域开展遗传育种的重要基础[1].遗传分化是种群遗传研究的重要部分,其核心内容是种群的划分和种群的分化规律[2].贵州地处中国西南的云贵高原东部,气候温暖湿润,属亚热带湿润季风气候区,自东向西呈现海拔逐渐升高的丘陵地势,森林覆盖面积大,全省东方蜜蜂分布广,种群数量多,是我国重要的东方蜜蜂主产区.
研究者对贵州省东方蜜蜂的形态指标进行了分析.与重庆、广西东方蜜蜂相比,贵州省东方蜜蜂存在特有的形态特征,说明贵州省东方蜜蜂在遗传资源上的特殊性[3-5].不同研究在对贵州省内东方蜜蜂的遗传分化结果上存在差异.杨冠煌[3]通过12个形态标记比较平均值,将贵州东部东方蜜蜂划分为华中生态型,贵州西部划分为云贵高原生态型;徐祖荫等[4,5]利用7个形态标记的平均值将贵州省东方蜜蜂划分为山地生态型与云贵高原生态型,以织金—册亨、赫章—盘县为界限分别分布在东西两侧;《中国畜禽遗传资源志·蜜蜂志》将贵州省东方蜜蜂划分为云贵高原中蜂和华中中蜂,其中华中中蜂分布于贵州省东部和东北部,云贵高原中蜂分布于贵州省西部[6];骆群等[7]利用32个形态标记进行更为客观的多元统计分析,未发现贵州省东方蜜蜂发生形态分化.
微卫星分子标记属于共显性遗传,遵循孟德尔遗传规律,多态信息含量高,检测遗传分化较为灵敏,能够用来解决形态标记等连续型数量标记无法解决的遗传多样性研究[8].已有研究利用3~21个微卫星标记分析了海南[9,10]、浙江大门岛[11]、泰国南部、北部、泰国苏梅岛东方蜜蜂的遗传分化[12],从分子水平描述了长白山[13]、黄土高原[14]、云南东方蜜蜂的遗传多样性水平[15].对于贵州省东方蜜蜂的微卫星标记分析尚无报道.
本研究利用31个微卫星位点对贵州全省的东方蜜蜂进行遗传分化与遗传多样性分析,为该地东方蜜蜂遗传资源提供基础数据,为东方蜜蜂的保护与利用提供依据.
1 材料与方法
1.1 材料
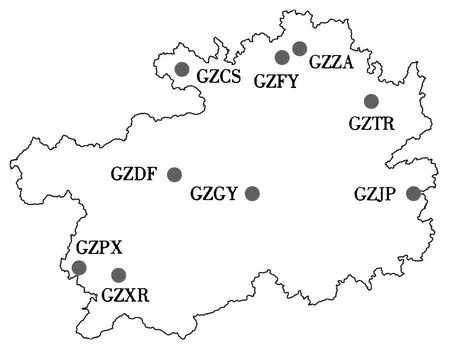
图1 贵州省东方蜜蜂样点Fig.1 Distribution of sample locations in Guizhou
根据贵州省的地形、地貌、以及东方蜜蜂的分布特点,本研究共采集9个样点的东方蜜蜂样本(图1),每个样点样本量在31~65群(表1),每个样点选取当地半野生状态的蜂群,以避免外来引种、商业买卖对样本的影响.
1.2 方法
采用Ezup柱式基因组DNA抽提试剂盒[生工生物工程(上海)股份有限公司]提取基因组.
采用ExTaq Hot Start Version多重扩增试剂盒[宝生物工程(大连)有限公司]和Toptaq多重扩增试剂盒[北京全式金生物技术有限公司]对31个微卫星位点进行多重PCR扩增[16,17](表2).ExTaq Hot Start Version多重扩增试剂盒的位点扩增程序为:95 ℃预变性15 min、95 ℃变性30 s、58 ℃退火40 s、72 ℃延伸40 s、循环次数30;72 ℃延伸10 min.Toptaq扩增试剂盒的扩增程序为:94 ℃预变性15 min、94 ℃变性30 s、58 ℃退火30 s、72 ℃延伸40 s、循环次数30;72 ℃延伸10 min.微卫星扩增产物送至苏州金唯智生物技术公司,利用ABI 3730xl自动测序仪(美国ABI)进行毛细管电泳,电泳内标选择GeneScanTMLIZ®[500],利用GeneMapper 4.0软件分析获得毛细管电泳图谱.
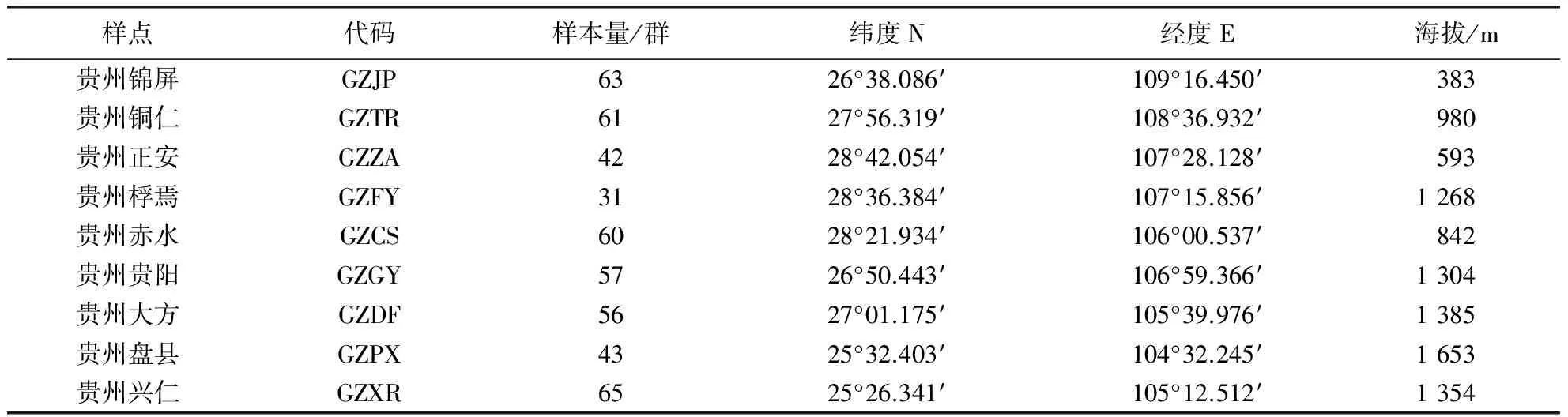
表1 贵州省东方蜜蜂样本信息Table 1 Information of sample locations from Guizhou
1.3 统计方法
利用Mstools软件计算期望杂合度(He)、观察杂合度(Ho)、等位基因数(Na),利用Popgene 1.31软件计算有效等位基因数(Ne)与香农指数(I)总计5个微卫星遗传多样性参数[18].
利用GenAlEx 6.5软件进行主坐标分析(PCoA),检测遗传分化情况[19];Genepop on Web在线软件分析样点间的FST值;通过FST与基因流Nm之间的公式FST=1/(1+4Nm),计算基因流Nm[20];利用Structure 2.3.4软件检测遗传分化情况[21],Markov chain Monte Carlo iterations设置为100000,burn-in设置为10000,利用clumpp 1.1.2和distruct 1.1对结果进行输出.为验证贵州省内部样本是否存在遗传分化,在各个分析中加入明显存在地理隔离的辽宁宽甸、海南那大样点[9,22]作为外群分析.

表2 微卫星位点信息1)Table 2 Microsatellite loci used in this study

续表2
1)/表示没有报道染色体的位置.
2 结果与分析
2.1 贵州省东方蜜蜂遗传分化
通过Structure分析、主坐标分析和遗传分化系数、基因流的分析等4种方法,没有发现贵州省东方蜜蜂发生遗传分化,全省9个样点的东方蜜蜂属于同一种群.
(1)Structure分析:Structure分析中,贵州样点与两个外群明显分化为3个不同的种群.贵州、辽宁、海南的样点分别显示为3种不同的颜色,其中贵州9个样点均为紫红色,辽宁样点为蓝色,海南样点为绿色(图2).并且不同色块颜色完整,说明分别为独立的3个种群.
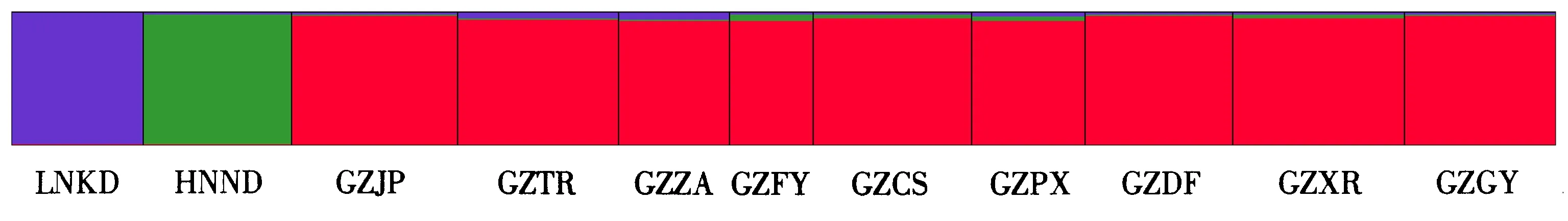
LNKD为辽宁宽甸样点、HNND为海南那大样点、贵州9个样点均为紫红色.图2 贵州各样点Structure结果图Fig.2 Structure analyses of sample locations from Guizhou

LNKD为辽宁宽甸样点,HNND为海南那大样点,GZ为贵州各样点.图3 贵州各样点主坐标分析图(PCoA)Fig.3 Principal coordinate analyses of sample locations from Guizhou
(2)主坐标分析:主坐标分析中,贵州样点在散点图中分布集中,为同一种群.两个外群分别在主坐标1和主坐标2上,与外群明显分离(图3).
(3)遗传分化系和基因流分析:贵州样点间的遗传分化系数小,基因流数值较大.贵州省9个样点间的遗传分化系数FST为0~0.03,与两个外群的FST值均大于0.08.
贵州省内部的基因流Nm范围在8.43~57.89之间,与外群的基因流数值均小于2.97(表3).
2.2 贵州省东方蜜蜂遗传多样性
贵州各个样点的遗传多样性水平较一致,平均期望杂合度为0.434 5±0.055 0,平均观察杂合度为0.419 6±0.011 4,平均等位基因数为5.56±4.34,平均有效等位基因数为2.693 0±2.113 5,平均香农指数为0.873 5±0.797 1(表4).遗传多样性最低的样点为贵州正安,其在期望杂合度、观察杂合度、有效等位基因数、香农指数上均表现最低.
3 讨论
(1)本研究通过微卫星中性分子遗传标记没有发现贵州省东方蜜蜂发生种群遗传分化.主要原因可能是蜜蜂分布连续,基因交流没有阻碍.贵州各样点间遗传分化系数为0~0.03,按照Wright标准[23]属于无遗传分化的水平.基因流为8.43~57.89,参照西方蜜蜂和东方蜜蜂上的研究,基因交流频繁[24].贵州省东方蜜蜂在全省范围的69个市县均有分布[25],蜂群总数量约为14万群[26],东方蜜蜂在贵州省境内能够连续并且以大种群的形式分布,境内没有明显的隔离屏障,样点之间不存在基因流的阻碍,所以没有发现种群遗传分化.

表3 贵州各样点间遗传分化系数FST和基因流Nm1)Table 3 FST and Nm between sample locations from Guizhou
1)左下角为遗传分化系数FST,右上角为基因流Nm.
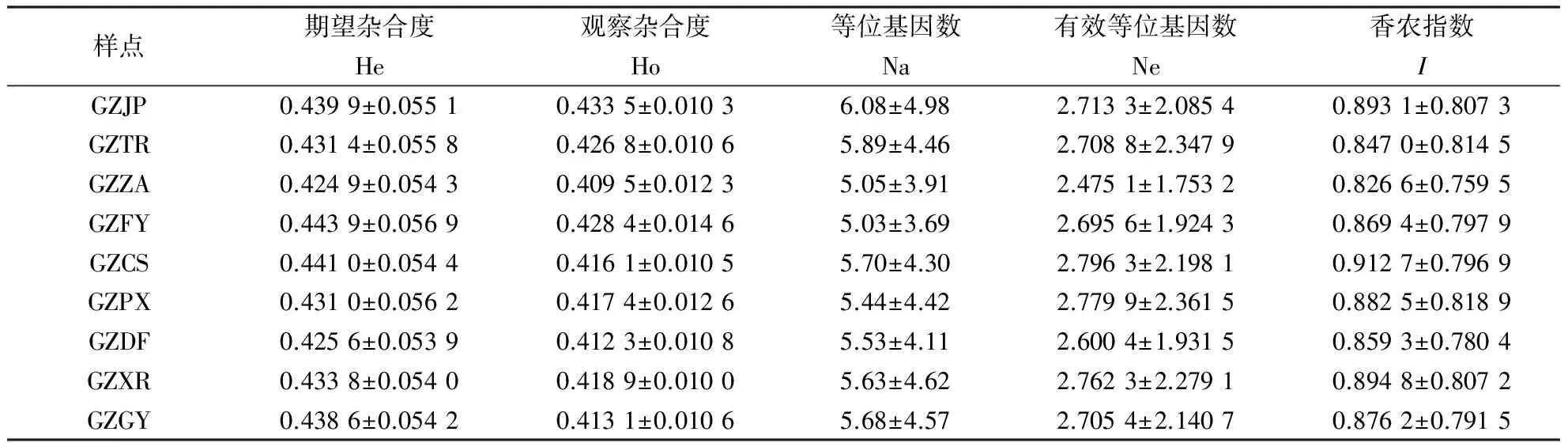
表4 贵州省各样点遗传多样性参数Table 4 Genetic diversity parameters of sample locations from Guizhou
(2)根据现有的文献分析,贵州省东方蜜蜂遗传多样性高于全国平均水平.主要原因可能是东方蜜蜂在云贵高原东部分布时间长,环境资源丰富,东方蜜蜂种群数量大.与全国其它地区相同微卫星位点的遗传多样性相比,贵州省东方蜜蜂的遗传多样性水平处于较高水平,仅略低于华东地区.与海南[9]同类研究中相同的7个微卫星比较,贵州省样本的期望杂合度为0.648 3,高于海南的0.614 6,与云南[15]、华东[27]同类研究中相同的4个微卫星比较,贵州省样本期望杂合度为0.813 9,高于云南的0.785 6,略低于华东的0.8570,与长白山[13]同类研究中相同的1个微卫星比较,贵州省样本期望杂合度为0.8280,高于长白山的0.142 7.另外,通过与我们在全国31个微卫星数据比较(未发表),贵州省东方蜜蜂的遗传多样性水平在期望杂合度、观察杂合度和等位基因数上高于全国的平均水平,显示了丰富的遗传多样性水平.分布贵州省各个地区的样点基因交流顺畅,基因流Nm为8.43~57.89,并且贵州省东方蜜蜂蜂群数量较大,占全国东方蜜蜂群数10%左右,整个贵州省的东方蜜蜂属于同一个大种群.
(3)贵州省正安县东方蜜蜂活框饲养技术推广早,人工育王较普遍.人工育王常因母群选择的片面,在少量蜂群中移虫,使蜜蜂遗传多样性丧失.微卫星数据中贵州正安在SV039、BI314、AC011位点上多样性低于其余8个样点,在这3个位点上,以1~2种等位基因为主,所占比例为0.93、0.68和0.69,极可能是不当的育王活动造成了这3个位点的等位基因受到了人为选择.
致谢:本研究样本的获得过程中得到了贵州省农业厅、贵州省养蜂学会徐祖荫和樊芸的协助.本科生夏凤枝参与本研究样本采集工作,硕士生郭慧萍、郝璐楠、本科生王恒、夏凤枝参与部分实验工作.
[1] 陈灵芝,马克平.生物多样性科学:原理与实践[M].上海:上海科学技术出版社,2001.
[2] PALUMBI S R. Genetic divergence, reproductive isolation, and marine speciation[J]. Annual Review of Ecology & Systematics, 2003,25(25):547-572.
[3] 杨冠煌.中华蜜蜂[M].北京:中国农业科技出版社,2001:8-21.
[4] 徐祖荫.贵州省中蜂资源调查及其开发利用研究——贵州中蜂不同生态类型的划分、评价及开发利用意见[J].贵州畜牧兽医,1986(1):13-19.
[5] 徐祖荫.蜂海求索——徐祖荫养蜂论文集[C].贵阳:贵州科技出版社,2010:32-40.
[6] 国家畜禽遗传资源委员会.中国畜禽遗传资源志:蜜蜂志[M].北京:中国农业出版社,2011.
[7] 骆群,周姝婧,徐新建,等.贵州东方蜜蜂形态遗传分析[J].福建农林大学学报(自然科学版),2015,44(3):298-302.
[8] ALLENDORF F W, HOHENLOHE P G. Genomics and the future of conservation genetics[J]. Nature Reviews Genetics, 2010,11(10):697-709.
[9] 徐新建,周姝婧,朱翔杰,等.海南岛中华蜜蜂遗传多样性的微卫星DNA分析[J].昆虫学报,2013,56(5):554-560.
[10] ZHAO W, WANG M, LIU Y, et al. Phylogeography ofApiscerana, populations on Hainan island and southern mainland China revealed by microsatellite polymorphism and mitochondrial DNA[J]. Apidologie, 2016:1-12.
[11] XU X, ZHU X, ZHOU S, et al. Genetic differentiation betweenApisceranacerana, populations from Damen Island and adjacent mainland in China[J]. Acta Ecologica Sinica, 2013,33(3):122-126.
[12] SITTIPRANEED S, LAOAROON S, KLINBUNGA S, et al. Genetic differentiation of the honey bee (Apiscerana) in Thailand: evidence from microsatellite polymorphism.[J]. Journal of Apicultural Research, 2001,40(1):9-16.
[13] 于瀛龙,周姝婧,徐新建,等.长白山中华蜜蜂(Apisceranacerana)遗传多样性分析[J].福建农林大学学报(自然科学版),2013,42(6):643-647.
[14] 徐新建,周姝婧,朱翔杰,等.黄土高原中华蜜蜂遗传多样性的微卫星DNA分析[J].福建农林大学学报(自然科学版),2013,42(6):638-642.
[15] 殷玲,吉挺,陈国宏.利用微卫星标记分析云南省6个东方蜜蜂(Apiscerena)群体遗传多样性及遗传分化[J].西南农业学报,2011,24(2):772-778.
[16] SOLIGNAC M, VAUTRIN D, LOISEAU A, et al. Five hundred and fifty microsatellite markers for the study of the honeybee (ApismelliferaL.) genome[J]. Molecular Ecology Notes, 2003,3(2):307-311.
[17] TAKAHASHI J, SHIMIZU S, KOYAMA S, et al. Variable microsatellite loci isolated from the Asian honeybee,Apiscerana(Hymenoptera; Apidae)[J]. Molecular Ecology Resources, 2009,9(3):819-821.
[18] PARK S. Trypanotolerance in west African cattle and the population genetic effects of selection[D]. Dublin: University of Dublin, 2001.
[19] PEAKALL R, SMOUSE P E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update[J]. Bioinformatics, 2012,28(19):2 537-2 539.
[20] ROUSSET F. genepop′007: a complete re-implementation of the genepop software for Windows and Linux.[J]. Molecular Ecology Resources, 2008,8(1):103.
[21] PRITCHARD J K, WEN X, FALUSH D. Documentation for STRUCTURE software: version 2.3[J]. Journal of Pediatric Surgery, 2010,41(10):55-63.
[22] 郭慧萍,周姝婧,朱翔杰,等.秦巴山区中华蜜蜂种群微卫星DNA遗传分析[J].昆虫学报,2016,59(3):337-345.
[23] WRIGHT S. Variability within and among Natural Populations[M]. Chicago: University of Chicago Press, 1978:439-459.
[24] NIKOLOVA S, BIENKOWSKA M, GERULA D, et al. Microsatellite DNA polymorphism in selectively controlledApismelliferacarnicaandApismelliferacaucasicapopulations from Poland[J]. Archives of Biological Sciences, 2015,67(3):889-894.
[25] 徐祖荫.贵州省蜂业区划[J].贵州畜牧兽医,1987(3):28-32.
[26] 徐祖荫.贵州省中蜂资源调查[C].重庆:第三届中国畜牧科技论坛,2007:436-440.
[27] 吉挺,殷玲,刘敏,等.华东地区中华蜜蜂六地理种群的遗传多样性及遗传分化[J].昆虫学报,2009,52(4):413-419.
(责任编辑:吴显达)
Genetic diversity and genetic differentiation ofApisceranain Guizhou Province of southwest China
YU Yinglong, ZHOU Shujing, XU Xinjian, ZHU Xiangjie, YANG Kaichieh, CHEN Daoyin, ZHOU Bingfeng
(College of Bee Science, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China)
Honeybees were analyzed using 31 microsatellite loci to assess the differentiation and diversity levels throughout Guizhou Province. Our data showed that Guizhou eastern honeybees have little divergence between honeybee sampling localities becauseFSTvalue is from 0 to 0.03. Our data showed that Guizhou eastern honeybees have a higher level of genetic diversity than honeybee elsewhere in China due to high value of expected heterozygosity (0.434 5±0.055 0), observed heterozygosity (0.419 6±0.011 4), number of alleles (5.56±4.34), number of effective alleles (2.693 0±2.113 5) and Shannon index (0.873 5±0.797 1). This study is of great value to understand the genetic structure, genetic differentiation, and genetic diversity, and to provide theoretical basis for the protection and utilization of eastern honeybee in Guizhou.
Apiscerana; genetic differentiation; genetic diversity; microsatellite DNA
2017-02-28
2017-03-21
现代农业蜂产业技术体系建设专项资金(CARS-45-KXJ11).
于瀛龙(1989-),男,博士研究生.研究方向:蜜蜂种群遗传学.Email:yuyinglong2012@163.com.通讯作者周冰峰(1958-),男,教授,博士生导师.研究方向:蜜蜂生态学和蜜蜂种群遗传学.Email:bingfengfz@126.com.
文献标识码: A 文章编号:1671-5470(2017)03-0323-06
10.13323/j.cnki.j.fafu(nat.sci.).2017.03.015
猜你喜欢
湖北植保(2022年4期)2022-08-23
种子(2021年3期)2021-04-12
土壤(2021年1期)2021-03-23
四川动物(2017年6期)2017-12-12
土壤学报(2017年5期)2017-11-01
四川动物(2017年4期)2017-07-31
外语教学理论与实践(2016年1期)2016-06-11
中学生理科应试(2016年7期)2016-05-14
中南林业科技大学学报(2015年6期)2015-12-20
河北遥感(2014年3期)2014-07-10
