
象山港春、夏季仔稚鱼种类组成结构特征
2017-07-12 16:19王宇坛李圣法杨林林袁兴伟姜亚洲
海洋渔业 2017年3期
王宇坛,李圣法,杨林林,袁兴伟,姜亚洲,林 楠
(1.中国水产科学研究院东海水产研究所;农业部东海与远洋渔业资源开发利用重点实验室,上海 200090;2.上海海洋大学海洋科学学院,上海 201306)
象山港春、夏季仔稚鱼种类组成结构特征
王宇坛1,2,李圣法1,杨林林1,袁兴伟1,姜亚洲1,林 楠1
(1.中国水产科学研究院东海水产研究所;农业部东海与远洋渔业资源开发利用重点实验室,上海 200090;2.上海海洋大学海洋科学学院,上海 201306)
为研究象山港仔稚鱼的时空分布特征及其与环境因子的相关关系,在象山港港湾内设置14个站,于2015年4月3日~6月10日期间按周采样,共调查10个航次(Ⅰ~Ⅹ),用方形网(网口为1 m×2 m,网目1.0 mm)采集仔稚鱼,同时采集温度、盐度和浮游动物等数据。调查期间采集仔稚鱼180 254 ind共45种,隶属25科39属,主要优势种为斑鰶(Konosirus punctatus)、矛尾鰕虎鱼(Chaeturichthys stigmatias)、鮻(Chelon haematocheilus)、斑尾刺鰕虎鱼(Acanthogobius ommaturus)、普氏缰鰕虎鱼(Amoya pflaumi)、拟矛尾鰕虎鱼(Parachaeturichthyspolynema)、黑棘鲷(Acanthopagrus schlegelii)、舌鰕虎鱼(Glossogobiusgiuris)等。仔稚鱼密度呈先升后降的趋势,第Ⅲ航次密度最高(平均密度为587.9 ind·100 m-3),第Ⅷ航次最低(8.2 ind·100 m-3)。前弯曲期仔鱼最多,占63.6%,弯曲期仔鱼占26.4%。基于层级聚类(Bray-Curtis相似性指数,Ward法聚类)分析表明,可将10航次仔稚鱼分为4个类型,各类型间存在明显差异:类型1(Ⅰ和Ⅱ航次)主要为矛尾鰕虎鱼和斑尾刺鰕虎鱼,类型2(Ⅲ~Ⅵ航次)为斑鰶、鮻、矛尾鰕虎鱼和黑棘鲷,类型3(Ⅶ和Ⅷ航次)为斑鰶、日本鳀(Engraulis japonicus)、拟矛尾鰕虎鱼,类型4(Ⅸ和Ⅹ航次)为普氏缰鰕虎鱼、拟矛尾鰕虎鱼、美肩鳃鳚(Omobranchus elegans)。Spearman相关分析表明,仔稚鱼分布与温度极显著相关(P<0.01),与浮游动物密度呈显著相关(P<0.05),与盐度无明显相关关系(P>0.05)。
象山港;仔稚鱼;种类组成;环境因子
仔稚鱼作为鱼类早期生活史的重要阶段,对渔业种群资源的补充至关重要[1-3]。仔稚鱼游泳能力差,逃避不利环境条件的能力差,营随波逐流生活,极易受海洋环境条件影响[4]。非生物因子(温度、盐度等)和生物因子(饵料生物)等对仔稚鱼分布有着重要影响[5-6]。同时,仔稚鱼时空分布特征是研究渔业资源可持续利用的重要基础,有利于提高对鱼类早期生命阶段的了解,推测亲体产卵的模式和繁殖策略[7]。因此,仔稚鱼分布以及其栖息环境的调查研究是至关重要的生态学研究。
象山港位于浙江省中部,是沿东北西南走向的狭长型半封闭海湾,是优良的鱼类产卵和育幼场所[8],亦是蓝点马鲛(Scomberomorus niphonius)种质资源保护区(中华人民共和国农业部第1491号公告),港湾内蓝点马鲛早期个体主要出现时间为4月中旬至6月初[9-10]。研究表明蓝点马鲛仔稚鱼主要摄食其它鱼类的早期个体[11],但至今尚未有关其饵料背景研究报告,一定程度上影响了保护区工作的开展。为此,通过2015年4~6月象山港仔稚鱼调查,分析仔稚鱼种类组成和分布特征及其环境因子的影响,为种质资源保护、渔业资源合理利用提供科学依据。
1 材料与方法
1.1 调查方法与样品采集
2015年4月3日~6月10日,用方形网(网口为1 m×2 m,网目1.0 mm)[9]按周对布设于象山港湾内的14个站点进行10个航次(Ⅰ~Ⅹ)的仔稚鱼调查(图1)。采样时,网口系有浮球和流量计(Hydro-Bios 438115型),拖绳长30~50 m,船速为1~2节,水平拖拽10 min。分别记录放网和起网时流量计数字以计算滤水体积,用无水乙醇溶液固定样品,带回实验室进行分析。为研究仔稚鱼与浮游动物密度的相关性,在仔稚鱼采样同时,以浮游生物Ⅱ网[12]同步采集浮游动物,样品以5%福尔马林溶液固定。各站点温度和盐度采用SEB-37 CTD测量。
在室内对样品进行分拣、鉴定,并按KENDALL等[13]的仔稚鱼发育分期标准划分仔稚鱼发育阶段,划分为:卵黄囊期仔鱼(yolk-sac larva)、前弯曲期仔鱼(preflexion larva)、弯曲期仔鱼(flexion larva)、后弯曲期仔鱼(postflexion larva)和稚鱼(juvenile)。其种名按照伍汉霖等[14]进行排列。浮游动物样品按《海洋调查规范》(GB 12763.6—1991)方法进行计数。
1.2 数据分析
采用Margalef丰富度指数(D),Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J′)进行多样性研究[15]。使用斯皮尔曼秩相关系数(Spearman’s rank correlation)分析仔稚鱼密度与环境因子之间的关系[16],环境因子主要包括海水温度、盐度以及浮游动物密度。
利用多元分析方法分析象山港仔稚鱼种类组成的时间变化,计算各航次间的Bray-Curtis相似性系数,应用未加权的组平均方法(UPGMA)进行级聚类分析和非度量多维标度(NMDS)的排序方法分析仔稚鱼组成特征[17-18]。采用单因子相似性分析(ANOSIM)对不同类型的仔稚鱼结构差异进行显著性检验,检验各类型分类的合理性[18]。以分类结果为依据,计算各类型相似性百分比(SIMPER)[19],分析造成各类型组成结构相似的典型种和各类型间组成差异的主要种类,以及鱼种对各类型的相似性和类型间的相异性贡献率。
2 结果与分析
2.1 仔稚鱼种类组成
2015年4~6月象山港水域仔稚鱼调查,共出现仔、稚鱼45种。其中鉴定到种34种,鉴定到属8种,鉴定到科2种,未鉴定种1种。在已鉴定的34种仔稚鱼中,鲈形目18种,鲱形目5种,鲽形目4种,鲉形目3种,鲻形目和刺鱼目各2种,其中鰕虎鱼科所占比例最高(共11种),占总种数的24.4%。优势种类为斑鰶(Konosirus punctatus),占总尾数的25.8%,其后依次为矛尾鰕虎鱼(Chaeturichthys stigmatias)(23.9%)、鮻(Chelon haematocheilus)(19.2%)、斑尾刺鰕虎鱼(Acanthogobius ommaturus)(9.9%)、普氏缰鰕虎鱼(Amoya pflaumi)(8.5%)、拟矛尾鰕虎鱼(Parachaeturichthys polynema)(4.5%)、黑棘鲷(Acanthopagrus schlegelii)(2.5%)、舌鰕虎鱼(Glossogobius giuris)(1.2%)。此外还出现了宽体舌鳎(Cynoglossus robustus)、褐菖鲉(Sebastiscus marmoratus)、蓝点马鲛、日本鳀(Engraulis japonicus)、焦氏舌鳎(Cynoglossus joyneri)、鲻(Mugil cephalus)、黄鲫(Setipinna tenuifilis)、黄姑鱼(Nibea albiflora)、中国花鲈(Lateolabrax maculatus)、小黄鱼(Larimichthys polyactis)等众多经济种类。从生态类型上说,主要为近岸定居种鰕虎鱼科、斑鰶、鮻等;岩礁性种类黑棘鲷、褐菖鲉等;海洋洄游性种类蓝点马鲛等;近海性种类日本鳀、中国花鲈等;偶入种小黄鱼、鲀科鱼类等(表1)。

表1 象山港水域春、夏季仔稚鱼种类与生态类型Tab.1 Species composition and ecological type in the Xiangshan Bay in spring and summer
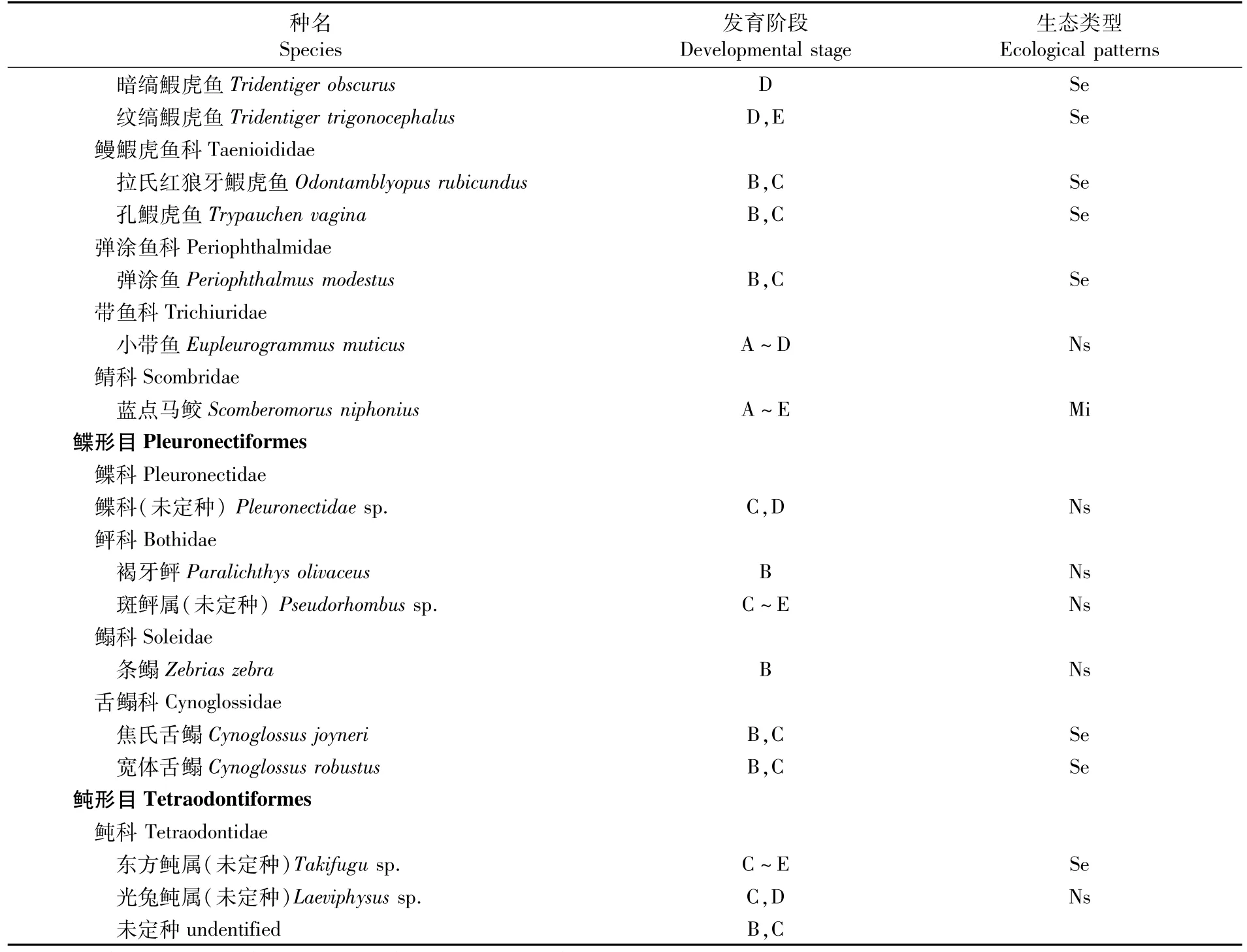
·续上表·
2.2 仔稚鱼多样性
象山港仔稚鱼调查,航次Ⅰ和航次Ⅱ仔稚鱼种类数(S)相对较少,随后各航次种类数有所增加。Shannon-Wiener多样性指数(H′)呈缓慢上升的趋势,最大值出现在航次Ⅶ(H′=2.14)。Margalef种类丰富度指数(D)和Pielou均匀度指数(J′)变化幅度较小,其中二者在航次Ⅲ至航次Ⅵ均较低(图2),这可能与少数种类的大量出现有关,如斑鰶、鮻、矛尾鰕虎鱼等。
2.3 仔稚鱼密度变化
仔稚鱼密度呈先增后减的趋势,其中平均密度最大和最小值分别出现在第Ⅲ和第Ⅷ航次,分别为587.9 ind·100 m-3和8.2 ind·100 m-3。
前弯曲期仔鱼个体数占总渔获量的63.6%,远高于其它发育阶段仔鱼所占比例,其次是弯曲期仔鱼(26.4%),而卵黄囊期、后弯曲期、稚鱼期个体相对较少,分别占总个体数的0.8%、8.0%、1.2%。前弯曲期仔鱼主要集中在Ⅰ~Ⅵ航次,其中航次Ⅳ前弯曲期仔鱼所占比例最大(96.3%)。每个航次都有一定量弯曲期仔鱼分布,其中航次Ⅹ占比最大(66.1%),航次Ⅲ最小(3.5%)(图3)。

图2 象山港仔稚鱼多样性指数的航次间变化Fig.2 Variations of larvae and juveniles fish biodiversity induces in different cruises in the Xiangshan Bay

图3 象山港仔稚鱼各发育阶段密度Fig.3 Densities of individuals in different development stages in Xiangshan Bay
2.4 仔稚鱼种类组成时间变化
象山港各航次仔稚鱼组成聚类分析和NMDS分析显示,象山港仔稚鱼种类群聚可分为4个类型(图4)。通过单因子相似性分析(ANOSIM),显示这4个类型呈极显著性差异(R=0.951、P<0.01),随着时间的变化,各类型仔稚鱼的种类组成和密度大小都有所不同。4月上旬(航次Ⅰ、Ⅱ)为类型1,其组内相似性达75%的仔稚鱼2种,分别是矛尾鰕虎鱼(61.1%)和斑尾刺鰕虎鱼(21.4%);4月中旬~5月中旬(航次Ⅲ~Ⅵ)为类型2,其相似性达75%的仔稚鱼共4种,分别是斑鰶(36.6%)、鮻(23.1%)、矛尾鰕虎鱼(12.0%)和黑棘鲷(5.3%);5月下旬(航次Ⅶ、Ⅷ)为类型3,使其相似性达75%的仔稚鱼共5种,分别是斑鰶(35.3%)、日本鳀(15.2%)、拟矛尾鰕虎鱼(10.7%)、黑棘鲷(9.4%)和侧带小公鱼属(未定种)(6.7%);6月上旬(航次Ⅸ、Ⅹ)为类型4,使其相似性达75%的仔稚鱼共4种,分别是普氏缰鰕虎鱼(44.7%)、拟矛尾鰕虎鱼(19.5%)、美肩鳃鳚(O.elegans)(7.4%)和尖海龙(Syngnathus acus)(7.0%)。
相似性百分比分析(SIMPER)显示,4个类型中相邻阶段之间的相异性分别为68.9%(类型1、2)、70.2%(类型2、3)、71.7%(类型3、4)。类型1、2和类型2、3相异的主要贡献种有斑鰶、鮻、矛尾鰕虎鱼等。引起类型3、4相异的主要贡献种为普氏缰鰕虎鱼、斑鰶(表2)。斑鰶、鮻、矛尾鰕虎鱼等仔稚鱼的密度随时间的变化引起前3个类型的相异,第4类型中普氏缰鰕虎鱼出现,且密度较大,是引起类型3、4的相异性的主要原因。
2.5 环境因子对仔稚鱼分布的影响
调查期间象山港水域的表层温度为14.3~22.7℃,自4月份至6月份水温呈逐渐升高趋势,同一航次中各站位温度差异不大(图5),相关分析表明仔稚鱼密度与温度呈极显著相关(R=-0.346,P<0.01)(表3)。各航次平均盐度变化不大,各站位平均盐度范围为24.7~26.0,从港口至港底呈递减趋势(图5)。相关分析表明仔稚鱼密度与盐度无明显相关关系(P>0.05)(表3)。4月至6月象山港湾内浮游动物密度变化波动较大(图6),其中第Ⅸ航次密度最大(755.2 ind·100 m-3),第Ⅰ航次最小(34.9 ind·100 m-3),相关分析表明浮游动物密度与仔稚鱼密度呈显著相关(R=0.189,P<0.05)(表3)。

图4 象山港仔稚鱼航次组成聚类分析图(左)和MDS排序图(右)Fig.4 Hierarchical cluster dendrogram(left)and MDS ordination diagram(right)from catch in different cruises

表2 象山港仔稚鱼组成各类型相异性主要贡献种类Tab.2 Dissim ilarity percentages of the prim ary species between types in the Xiangshan Bay

表3 仔稚鱼与环境因子间的斯皮尔曼秩相关系数Tab.3 Spearman’s rank correlation coefficients between larvae and juvenile fish density and environmental variables

图5 春、夏季象山港表层水温、表层盐度变化Fig.5 Variation of the temperature and the salinity in the Xiangshan Bay in spring and summer

图6 象山港浮游动物各航次平均密度分布Fig.6 Average zoop lankton densities in different cruises in the Xiangshan Bay
3 讨论
3.1 仔稚鱼种类组成
港湾作为鱼类重要的产卵和育幼场所,通常是以小型鱼类利用为主,如在莱州湾[20]、罗源湾[21]和东山湾[22]仔稚鱼的主要优势种为鳀科种类,泉州湾[23]的为鱚科、三沙湾[24]的鰕虎鱼类等,国外的一些港湾也有类似情况,如在加利福尼亚湾[25]以鳀科和灯笼鱼科的仔稚鱼为优势种、墨西哥湾[26]的为鲱科和鳀科以及波斯湾[27]的鲱科、沙鮻科和鳚科等。象山港的仔稚鱼的调查研究结果亦呈现出以鰕虎鱼科和鲱科等小型鱼类为优势种的这一特征,这可能与港湾内成鱼的种类组成有关。有研究[28]表明在象山港的小型桁杆拖网中渔获物的主要优势种为鰕虎鱼类,而流网中以斑鰶为第一优势种。
根据生态类型,可将象山港鱼类分为3个类型:1)将港湾作为其产卵场和育幼场,如定居性种类鰕虎鱼类、斑鰶、鮻等,仔稚鱼的各发育阶段个体均有出现,亦有大量的成鱼长期生活在港湾内,这一类型还包括了主要增殖品种黑棘鲷和褐菖鲉等岩礁性鱼类。此外,长距离洄游种类蓝点马鲛亦将该水域作为产卵场和育幼场,其产卵群体于每年清明节前后进入港湾内产卵,但发育至稚鱼期之后离开象山港。2)将港湾仅作为其育幼场,如近海种日本鳀,其主要在高盐水域产卵,早期个体随流迁徙,进入港湾等较浅水域生长[29];中国花鲈亦为如此,其主要在较深水域产卵,发育至后期个体进入港湾育幼[30]。3)为偶入种,邻近水域产卵场的种类,其早期个体在海流作用下,零星进入港湾,本研究中很大一部分种类为这一类型,如小黄鱼、颌针鱼属、红娘鱼属、蛇鳗科和鲀科鱼类等。
3.2 仔稚鱼与环境之间的关系
海洋环境因子的变化对仔稚鱼的密度分布影响较为明显,如,水温、盐度、潮汐[31]、饵料生物丰富度等。其中水温和盐度的变动会直接影响仔稚鱼的形态发育、生长及数量分布等[32]。本研究表明温度影响仔稚鱼密度分布,相关性分析结果为仔稚鱼密度与温度的变化极显著负相关(P<0.01)。水温影响卵的孵化和仔稚鱼生长发育,孵化率的高低决定仔稚鱼的密度大小,4月下旬至5月上旬港内温度适宜,亲鱼集中产卵,卵孵化率高,仔稚鱼密度较大。相关性分析结果表明仔稚鱼密度与海水盐度无显著相关关系,这可能与调查时间、范围有关,此次调查区域相对较小,且象山港为半封闭港湾,与外界海水交换较慢,海水盐度变化不大,这些原因可能对分析结果产生一定的影响。
一般研究认为鱼类早期发育阶段主要摄食浮游动物[33],仔稚鱼分布受饵料生物的影响,通常表现为两者变化趋势较为一致,如王建茹等[34]通过对福建三沙湾中浮游动物与鱼卵和仔稚鱼分布的研究,发现两者密度的变化趋势一致。阙江龙等[35]对北部湾西北部浮游动物季节变化及其与鱼卵、仔稚鱼的关系的研究表明两者具有较强的正相关关系。象山港仔稚鱼和浮游动物两者间亦呈现出显著相关,与上述结果一致。丰富的饵料生物是鱼类选择产卵场和育幼场的条件之一[36],仔稚鱼出现时间与饵料生物出现时间的匹配性直接影响仔稚鱼的存活率[37],本研究中两者的变化趋势一致,从侧面反映了该水域之所以成为众多鱼类产卵场和育幼场的原因,而丰富的仔稚鱼资源也为蓝点马鲛在该水域产卵和育幼提供了有利条件。
[1] HJORT J.Fluctuations in the great fisheries of northern Europe viewed in the lightof biological research[J].Rapports et Proies-Verbaux des Réunions,1914(4):1-228.
[2] 万瑞景,孙 珊.黄、东海生态系统中鱼卵、仔稚幼鱼种类组成与数量分布[J].动物学报,2006,52(1):28-45.WAN R J,SUN S.The category composition and abundance of ichyoplankton in the ecosysterm of Yellow Sea and the East China Sea[J].Acta Zoologica Sinica,2006,52(1):28-45.
[3] 黄凤鹏,黄景洲,杨玉玲,等.胶州湾鱼卵、仔鱼和稚鱼的分布[J].海洋科学进展,2007,25(4):468-473.HUANG F P,HUANG J Z,YANG Y L,et al.Distributions of fish eggs and larval fish in the Jiaozhou Bay[J].Advances in Marine Science,2007,25(4):468-473.
[4] ROBERTS C M.Connectivity and management of Caribbean coral reefs[J].Science,1997,278(5342):1454-1457.
[5] AOKIA I,MIYASHITAB K.Dispersal of larvae and juveniles of Japanese anchovy Engraulis japonicus in the Kuroshio Extension and Kuroshio-Oyashio transition regions,western North Pacific Ocean[J].Fisheries Research,2000(2):155-164.
[6] KIM J Y,KANG Y S,OH H J,et al.Spatial distribution of early life stages of anchovy(Engraulis japonicus)and hairtail(Trichiurus lepturus)and their relationship with oceanographic features of the East China Sea during the 1997-1998 El Niño Event[J].Estuarine Coastal and Shelf Science,2005,63(1-2):13-21.
[7] NONAKA RH,YASUNOBUM,SUZUKIK.Seasonal variation in larval fish assemblages in relation to oceanographic conditions in the Abrolhos Bank region off eastern Brazil[J].Fishery Bulletin,2000,98(4):767-784.
[8] 唐峰华,李 磊,廖 勇,等.象山港海洋牧场示范区渔业资源的时空分布[J].浙江大学学报(理学版),2012,39(6):696-702.TANG F H,LI L,LIAO Y,et al.Spatial and temporal distribution on fishery resources of marine pasture demonstration area in Xiangshan Harbor[J].Journal of Zhejiang University(Science Edition),2012,39(6):696-702.
[9] 宋 超,王宇坛,刘尊雷,等.象山港蓝点马鲛鱼卵、仔稚鱼的时空分布特征及其与环境因子关系[J].中国水产科学,2016,23(5):1197-1204.SONG C,WANG Y T,LIU Z L,et al.Relationship between environmental factors and distribution of Scomberomorus niphonius eggs,larvae,and juveniles in Xiangshan Bay[J].Journal of Fishery Sciences of China,2016,23(5):1197-1204.
[10] 楼 丹,施慧雄,焦海峰,等.象山港蓝点马鲛渔获情况初步研究[J].河北渔业,2010(10):38-39.LOU D,SHIH X,JIAO H F,et al.Fishing output for spawningstock of Scomberomorus niphonius in Xiangshan Bay[J].Hebei Fishery,2010(10):38 -39.
[11] SHOJI J,KISHIDA T,TANAKA M.Piscivorous habits of Spanish mackerel larvae in the Seto Inland Sea[J].FISHERIESSCIENCE,1997,63(3):388 -392.
[12] 朱艺峰,王 银,林 霞,等.象山港两种网目网采浮游动物群落比较[J].应用生态学报,2012,23(8):2277-2286.ZHU Y F,WANG Y,LIN X,et al.A comparison of zooplankton communities collected by two types of nets with different mesh sizes in Xiangshan Bay of Zhejiang,East China[J].Chinese Journal of Applied Ecology,2012,23(8):2277-2286.
[13] KENDALL A W J,AHLSTROM E H,MOSER H G.Early life history stages of fishes and their characters[M]//Ontogeny and systematics of fishes.Lawrence:The American Society of Ichthyologists Herpetologists,1984.
[14] 伍汉霖,邵广昭,赖春福,等.拉汉世界鱼类系统名典[M].基隆:水产出版社,2012.WU H L,SHAO G Z,LAI C F,et al.Latin-Chinese dictionary of fishes names by classification[M].Keelung:The Sueichan Press,2012.
[15] 马克平,刘玉明.生物群落多样性测度方法Ⅰα多样性的测度方法(下)[J].生物多样性杂志,1994,2(4):231-239.MA K P,LIU Y M.Measurement of species diversity,Ⅰαdiversity(second part)[J].Biodiversity Science,1994,2(4):231-239.
[16] RODRIGUEZ JM,GONZALEZN G,GONZALEZP C,et al.The ichthyoplankton assemblage and the environmental variables off the NW and N Iberian Peninsula coasts,in early spring[J].Continental Shelf Research,2009,29(8):1145-1156.
[17] FIELD JG,CLARKE K R,WARWICK R M.A practical strategy for analyzing multispecies distribution patterns[J].Marine Ecology Progress Series,1982,8(1):37-52.
[18] CLARKE K R,WARWICK R M.Change in marine communities:An approach to statistical analysis and Interpretation[M].Plymouth:Natural Environment Research Council,1994.
[19] CLARKE K R,AINSWORTH M.A method of linking multivariate community structure to environmental variables[J].Marine Ecology Progress Series,1993,92(3):205-219.
[20] 高彦洁,吕振波,杨艳艳,等.莱州湾春季鱼卵仔稚鱼群落结构和物种多样性[J].生态学报,2016,36(20):6565-6573.GAO Y J,LV ZB,YANG Y Y,etal.Structure and species diversity of ichthyoplankton in spring in Laizhou Bay[J].Acta Ecologica Sinica,2016,36(20):6565-6573.
[21] 刘 勇,沈长春.罗源湾夏季鱼卵和仔稚鱼种类组成与数量分布[J].福建水产,2012,34(4):309-315.LIU Y,SHEN C C.Species composition and quantity distribution of ichthyoplankton in Luoyuan bay in summer[J].Journal of Fujian Fisheries,2012,34(4):309-315.
[22] 郑惠东.福建东山湾春、夏季鱼卵和仔稚鱼丰度分布特征及其与环境因子的关系[J].应用海洋学学报,2016,35(1):87-97.ZHENG H D.Abundance distribution of fish eggs and larvae and its relation with environmental factors in spring and summer in Dongshan Bay,Fujian[J].Journal of Applied Oceanography,2016,35(1):87 -97.
[23] 戴燕玉.泉州湾浮性鱼卵和仔、稚鱼的种类和数量分布[J].福建水产,2005,6(2):15-19.DAIY Y.Distribution of ichthyoplankton larva and juvenile of fish in Quanzhou Bay[J].Journal of Fujian Fisheries,2005,6(2):15-19.
[24] 沈长春.福建三沙湾鱼卵、仔稚鱼种类组成及其丰度时空分布[J].海洋渔业,2011,33(4):361 -367.SHEN C C.Species composition and abundance trmporal-spatial distribution of fish egg,larvae and juveniles in Sansha Bay of Fujian[J].Marine Fisheries,2011,33(4):361-367.
[25] PEGUERO IM,SANCHEZ V L,LAVIN M F,et al.Larval fish assemblages,environment and circulation in a semienclosed sea(Gulf of California,Mexico)[J].Estuarine,Coastal and Shelf Science,2008,79(2):277-288.
[26] ROSS S T,MCMICHAEL R H JR,RUPLE D L.Seasonal and diel variation in the standing crop of fishes and macroinvertebrates from a Gulf of Mexico surf zone[J].Estuarine,Coastal and Shelf Science,1987,25(4):391-412.
[27] RABBANIHA M,MOLINERO JC,LOPEZ L L,et al.Habitat association of larval fish assemblages in the northern Persian Gulf[J].Marine Pollution Bulletin,2015,97(1-2):105-110.
[28] YUAN X W,JIANG Y Z,LIN N,et al.Using bioacoustics and conventional netting methods to assess the initial effectiveness of a newly deployed artificial reef on fish assemblages in Xiangshan Bay near Ningbo,Zhejiang Province,China[J].Journal of Applied Ichthyology,2013,29(6):1430 -1435.
[29] 陈渊戈,钟俊生,徐兆礼,等.春季东海有害藻华高发区仔稚鱼时空分布[J].海洋学报,2015,37(2):44-54.CHEN Y G,ZHONG J S,XU Z L,et al.Spatiotemporal distribution of larval and juvenile fishes at frequent harm ful algal blooms(HABs)areas of the East china sea in spring[J].Haiyang Xuebao,2015,37(2):44-54.
[30] HIBINO M,OHTA T,ISODA T,et al.Distribution of Japanese temperate bass,Lateolabrax japonicus,eggs and pelagic larvae in Ariake Bay[J].Ichthyology Research,2007,54(4):367-373.
[31] 刘媛媛,宋 超,赵 峰,等.长江口及其邻近水域仔稚鱼种类组成及分布特征[J].海洋渔业,2016,38(6):597-608.LIU Y Y,SONG C,ZHAO F,et al.Category composition and distributional patterns of fish larvae and juveniles in Yangtze Estuary and adjacent waters[J].Marine Fisheries,2016,38(6):597-608.
[32] ABOOKIRE A A,PIATT J F,ROBARDS M D.Nearshore fish distribution in an Alaskan Estuary in relation to stratification,temperature and salinity[J].Estuarine,Costal and Shelf Science,2000,51(1):45-59.
[33] NUNN A D,TEWSON L H,COWX I G.The foraging ecology of larval and juvenile fishes[J].Reviews in Fish Biology and Fisheries,2012,22(2):377-408.
[34] 王建茹,林元烧,周美玉,等.福建三沙湾鱼卵和仔稚鱼的生态分布[J].厦门大学学报(自然科学版),2010,49(1):116-121.WANG J R,LIN Y S,ZHOU M Y,et al.The ecological distribution of fish eggs and larvae in Sansha Bay,Fujian[J].Journal of Xiamen University(Natural Science),2010,49(1):116-121.
[35] 阙江龙,徐兆礼,孙鲁峰.北部湾西北部饵料浮游动物季节变化及其与鱼卵、仔稚鱼的关系[J].中国水产科学,2015,22(5):1027-1035.QUE JL,XU Z L,SUN L F.Seasonal variation in zooplankton characteristics and their relationship with fish eggs and larvae in the Northwest Beibu Gulf[J].Journal of Fishery Sciences of China,2015,22(5):1027-1035.
[36] ALAN K W,PAULA P.Habitat type and nursery function for coastal marine fish species,with emphasis on the Eastern Cape region,South Africa[J].Estuarine,Coastal and Shelf Science,2015,160(2):49-59.
[37] MERTZG,MYERSR A.Match/mismatch predictions of spawning duration versus recruitment variability[J].Fisheries Oceanography,1994,3(4):236-245.
Species com position of larval and juvenile fish in the Xiangshan Bay in spring and summer
WANG Yu-tan1,2,LISheng-fa1,YANG Lin-lin1,YUAN Xing-wei1,JIANG Ya-zhou1,LIN Nan1
(1.Key Laboratory of East China Sea and Oceanic Fishery Resources Exploitation&Utilization,Ministry of Agriculture,East China Sea Fisheries Research Institute,Chinese Academy of Fishery Science,Shanghai 200090,China;2.College of Marine Science and Technology,ShanghaiOcean University,Shanghai 201306,China)
The larvae and juvenile fish assemblage was investigated in Xiangshan Bay to analyze their composition and distribution aswell as the relationships between the fish larva amount and the environmental factors.Fourteen sampling stationswere established,and larval and juvenile fish were weekly collected by the frame trawls(1 m×2 m,1.0 mm mesh size)from April 3 to June 10 in 2015.The water temperature,salinity and zooplankton at each sampling site were also measured at each sampling time.In total,180 254 larval and juvenile fisheswere collected belonging to 25 families and 45 species.The dominant fish species were Konosirus punctatus(25.8%),Chaeturichthys stigmatias(23.9%),Chelon haematocheilus(19.2%),Acanthogobius ommaturus(9.9%),Amoya pflaumi(8.5%),Parachaeturichthys polynema(4.5%),Acanthopagrus schlegelii(2.5%)and Glossogobius giuris(1.2%).The density of larval and juvenile fishes varied significantly among cruises(P<0.05).The highestmean density of larval and juvenile fishes was recorded in the third cruise(587.9 ind·100 m-3),while the lowestmean density(8.2ind·100 m-3)occurred in the eighth cruise.The number of species increased gradually from the first cruise to the last one.Fish larvae in preflexion stage accounted for 63.6%of the total catch.The flexion stage of larvaemade 26.4%contribution to the total catch.There were temporal variation in the dominant species in Xiangshan Bay.According to the Bray-Curtis similarity measure on the relative abundance of each taxonomic type,four principal typeswere identified,including Type 1(cruisesⅠandⅡ),Type 2(cruisesⅢtoⅥ),Type 3(cruisesⅦandⅧ)and Type 4(cruisesⅨandⅩ).Variations of some larval and juvenile fish density,such as Konosirus punctatus,Chelon haematocheilus,Chaeturichthys stigmatias,Acanthogobius ommaturus,Amoya pflaumi and Parachaeturichthys polynema were thought to account for the type variation in the larval and juvenile fish assemblage structure.The environmental factors can have important implications for the distribution,growth,and survival of larval and juvenile fish.Furthermore,wemeasured environmental factors and determined the responses of the larval and juvenile fish communities to the factors,including the temperature,the salinity and the zooplankton density.The results of the Spearman Correlation between the amount of fish and the simultaneous environmental factors showed that the fish larva amount had the distinct correlation with water temperature and zooplankton density,but the salinity had no remarkable relationship with the fish larva density.Zooplankton is the vital food source for most larval and juvenile fish specie.Variations in zooplankton distribution and quantity can directly or indirectly affect the larval and juvenile fish.High quantities of zooplankton provide the sufficient food to support the larval and juvenile fish populations and therefore provide a valuable condition for fish reproduction and growth.
Xiangshan Bay;larvae and juveniles fish;species composition;environment factors
S 931
A
1004-2490(2017)03-0286-11
2016-11-03
国家公益性行业(农业)科研专项(201303047)
王宇坛(1990-),男,硕士研究生,主要从事渔业资源和生态学研究。E-mail:1129529886@qq.com
林 楠,助理研究员。E-mail:linn@eastfishery.ac.cn
猜你喜欢
农村科学实验(2022年16期)2022-11-19
水产科学(2022年1期)2022-01-26
渔业科学进展(2021年6期)2022-01-04
浙江海洋大学学报(自然科学版)(2021年1期)2021-07-06
南方水产科学(2021年3期)2021-06-23
水产科学(2020年4期)2020-07-28
食品与发酵工业(2020年13期)2020-07-22
创新作文(3-4年级)(2019年8期)2019-12-17
创新作文(小学版)(2019年23期)2019-01-11
中国海洋大学学报(自然科学版)(2014年8期)2014-02-28
