
珍珠蚌5个群体不同阶段生长性能及外部形态变异分析
2017-08-07 10:33吴雷明白志毅刘晓军李家乐
淡水渔业 2017年4期
吴雷明,白志毅,2,刘晓军,2,金 参,殷 浩,李家乐,2
(1.上海海洋大学 农业部淡水水产种质资源重点实验室,上海 201306;2.上海海洋大学 上海市水产养殖工程技术研究中心,上海 201306)
珍珠蚌5个群体不同阶段生长性能及外部形态变异分析
吴雷明1,白志毅1,2,刘晓军1,2,金 参1,殷 浩1,李家乐1,2
(1.上海海洋大学 农业部淡水水产种质资源重点实验室,上海 201306;2.上海海洋大学 上海市水产养殖工程技术研究中心,上海 201306)
采用温室大棚早繁技术,获得三角帆蚌(Hyriopsiscumingii)选育群体、武义养殖群体、鄱阳湖、洞庭湖野生群体,池蝶蚌(Hyriopsisschlegelii)养殖群体的子代,对1龄和2龄蚌的生长性能进行分析,并利用三种多元统计分析方法对1龄蚌外部形态变异进行了研究。结果显示:珍珠蚌 5 个群体的养殖性能由高到低为:池蝶蚌群体>选育群体>鄱阳湖群体>洞庭湖群体>养殖群体。生长指标之间存在显著相关性,与体质量相关程度由大到小为:壳长、壳宽和壳高。1龄时,影响珍珠蚌体质量的生长指标主要为壳长,2龄时,为壳长和壳宽。珍珠蚌生长后期壳宽对体重的影响比重加大,壳高与体重的相关性降低。三种多元分析结果显示,1龄时,池蝶蚌群体与鄱阳湖群体、洞庭湖群体以及养殖群体的外部形态存在较大的相似性,综合判别率较低,选育群体外部形态较前四者存在明显差异。
池蝶蚌(Hyriopsisschlegelii);三角帆蚌(Hyriopsiscumingii);生长性能;形态差异
三角帆蚌(Hyriopsiscumingii)是我国特有种。池蝶蚌(Hyriopsisschlegelii)与三角帆蚌同属于帆蚌属,原产于日本的琵琶湖,为外来引进群体[1],是我国目前主要应用于生产淡水无核珍珠的两种优质育珠蚌。通过三角帆蚌与池蝶蚌育珠性能比较发现,两者所产珍珠性能各具有独特优点,三角帆蚌所产珍珠珠质细腻、光滑、色泽鲜艳、形状较圆,池蝶蚌具有珍珠质分泌能力强、珍珠层厚、产量高等优点[2]。
三角帆蚌广泛分布于洞庭湖、太湖、鄱阳湖等大中型湖泊及其流域,丰富的生态多样性造就了不同地理群体明显的生长差异和形态特征[3]。通过形态变异分析物种的遗传特性,具有研究方法直接且经济等优点[4]。国内学者对我国淡水蚌种间及群体间形态差异的研究已有一些报道。魏开建[5]等对褶纹冠蚌(Cristariaplicata)、三角帆蚌、圆背角无齿蚌(Anodontawoodianapacifica)、椭圆背角无齿蚌(Anodontawoodianaelloptica)和球形无齿蚌(Anodontaglobosula)5种蚌的种间以及地理种群间的形态变异进行了研究,发现不同淡水蚌种间存在明显的形态差异。钱荣华等[6]使用多元分析方法,对三角帆蚌不同地理种群间形态差异进行了分析,表明三角帆蚌不同地理种群的形态差异明显;罗会等[7]运用多变量形态度量学方法,对合浦珠母贝的4个不同地理群体的9个形态特征进行研究,也得出类似的结论。
本试验对三角帆蚌选育群体、武义养殖群体、鄱阳湖野生群体、洞庭湖野生群体和池蝶蚌养殖群体子代的生长性能及各生长指标之间的相关性进行了研究,并采用聚类分析、主成分分析、判别分析三种多元统计分析方法,对上述几个群体的子代的外部形态变异进行比较,旨为对不同淡水珍珠蚌群体的生长性能进行初步评价,为选育优良种质提供参考。
1 材料和方法
1.1 材料
选育群体为连续选育4代的紫色三角帆蚌(BP),鄱阳湖(PY)与洞庭湖(DT)野生群体采集于鄱阳湖与洞庭湖,池蝶蚌(JX)来源于江西省池蝶蚌良种场,所有样本于2014年1月收集完成后,养殖于浙江金华武义伟民有限公司;此外,养殖群体(FP)采用浙江金华武义伟民有限公司自留群体。实验采用早繁技术,挑选健康、喷水有力、性腺发育良好的个体作为亲本,吊养于温室大棚中进行培育。按照常规淡水珠蚌繁苗方法进行繁育,苗种经过2个月的培育,至壳长达到1.2~1.5 cm时,将稚蚌转入池塘进行网箱培育。网箱规格为40 cm×40 cm×10 cm,每个网箱中放120个稚蚌,网箱吊养于离水面40 cm深处,各群体保留10个网箱的稚蚌,养殖于上海崇明“坚新基地”(31°35′N,121°31′E)。养殖期间、做好施肥、水质调节等日常养殖管理工作。
1.2 生长指标与外部形态测量
2年池塘网箱养殖期间,1龄(6月龄)蚌各群体随机抽取40个个体,2龄(12月龄)蚌各群体取30个个体。使用游标卡尺和电子天平(PC8000)测量每个个体的生长指标。外部形态测量参数为壳长、壳宽、壳高等11个可量性状,测量部位与参数参考钱荣华等[6]。
1.3 数据统计与分析
利用Excel对各群体生长数据进行整理和录入,并制作长度性状与体质量之间的线性回归关系图。采用SPSS 17.0软件分析各群体生长指标的差异性和相关性,P>0.05代表无显著差异,P<0.05代表差异显著。采用聚类分析、主成分分析、判别分析3种多元分析方法对5个群体的子代进行形态变异分析。为消除蚌体规格大小对参数值的影响,以壳长为标准,将每只蚌的所有参数分别除以壳长进行标准化处理,得出10个形态学比例性状,再分别求出各组样本每个参数校正值的平均值,用于统计分析。
2 结果
2.1 1龄、2龄蚌生长性能及变异分析
由表1可知,BP体质量、壳长显著高于PY、DT以及FP,群体间壳宽差异不显著。BP体质量与JX无显著差异,壳长显著高于JX,壳宽显著低于JX,而BP生长性状的变异系数稍高。壳宽与壳高的比例性状对比中发现,JX显著高于其它几个群体,FP显著低于其它4个群体,BP与2个野生群体差异不显著。如表2所示,DT体质量变异系数最大,FP壳长、壳高变异系数最大,BP壳宽变异系数最大。BP体质量、壳长和壳高变异系数均低于其它群体,比鄱阳湖和洞庭湖2组野生群体分别低30.34%、23.49%、6.99%和54.65%、34.29%、8.37%。JX壳宽与壳高比例性状的变异系数最小,FP变异系数最大。
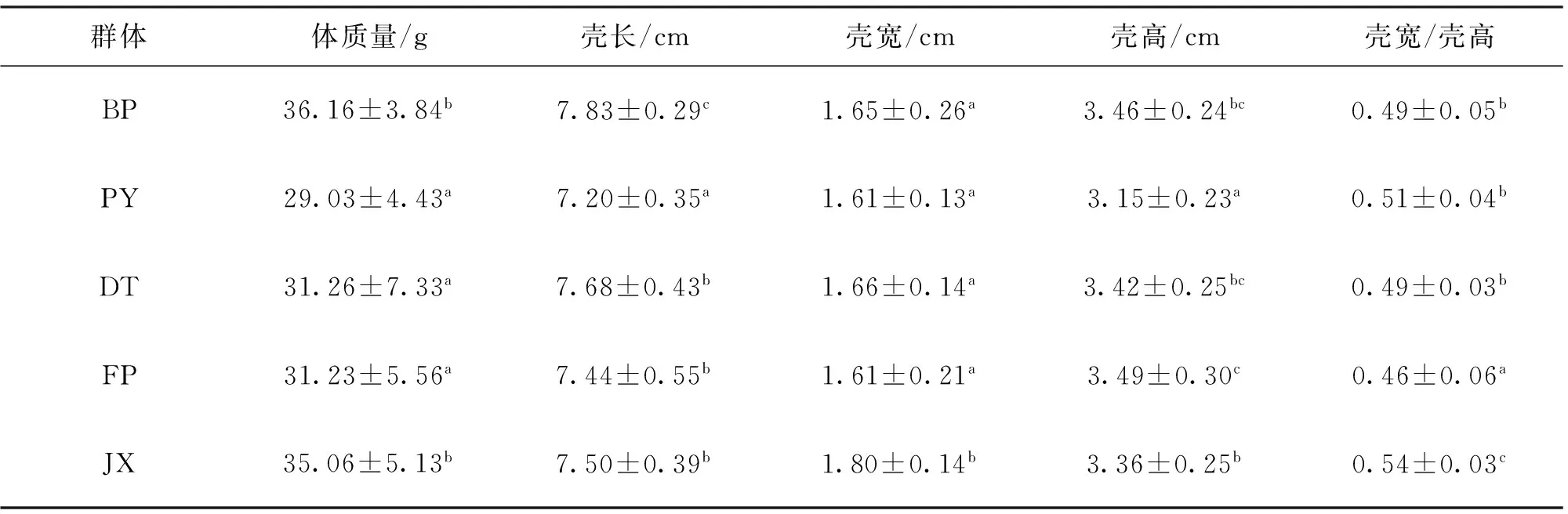
表1 5群体1龄蚌生长性能差异分析比较Tab.1 Comparison of growth traits among five populations
注:同列数不同小写字母代表差异显著(P<0.05)。表3同。
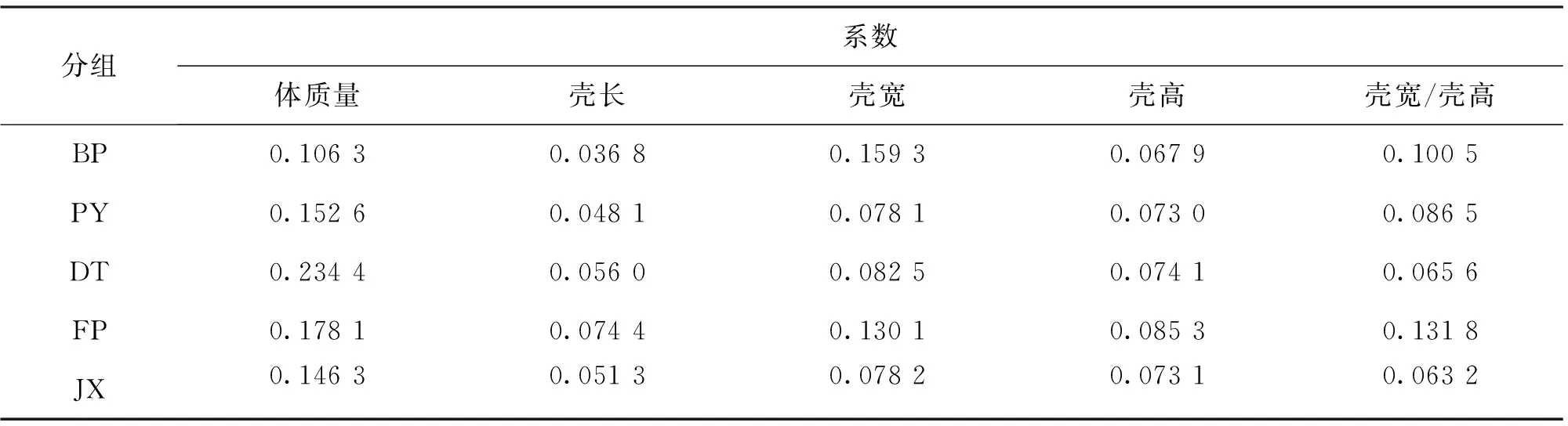
表2 5群体1龄蚌生长性状变异系数比较Tab.2 Comparison of variation coefficient of growth traits among five populations
由表3可知,BP 2龄阶段各生长指标显著高于PY、DT以及FP,其中体质量、壳宽和壳高3个生长指标与JX差异不显著。2个野生群体的体质量、壳长显著高于FP。JX各生长指标显著高于PY、DT以及FP。JX壳宽与壳高比例显著高于4个三角帆蚌群体。如表4所示,对2龄各群体生长性状变异系数进行了比较。BP除壳宽外,各生长指标的变异系数最小,其体质量、壳长比PY和DT分别低38.56%、8.75%和40.56%、19.20%。2个三角帆蚌野生群体之间各生长指标的变异系数相差不大,JX体质量、壳长、壳高变异系数最大,壳宽与壳高的比例变异系数仅高于BP,明显低于其它3个群体。
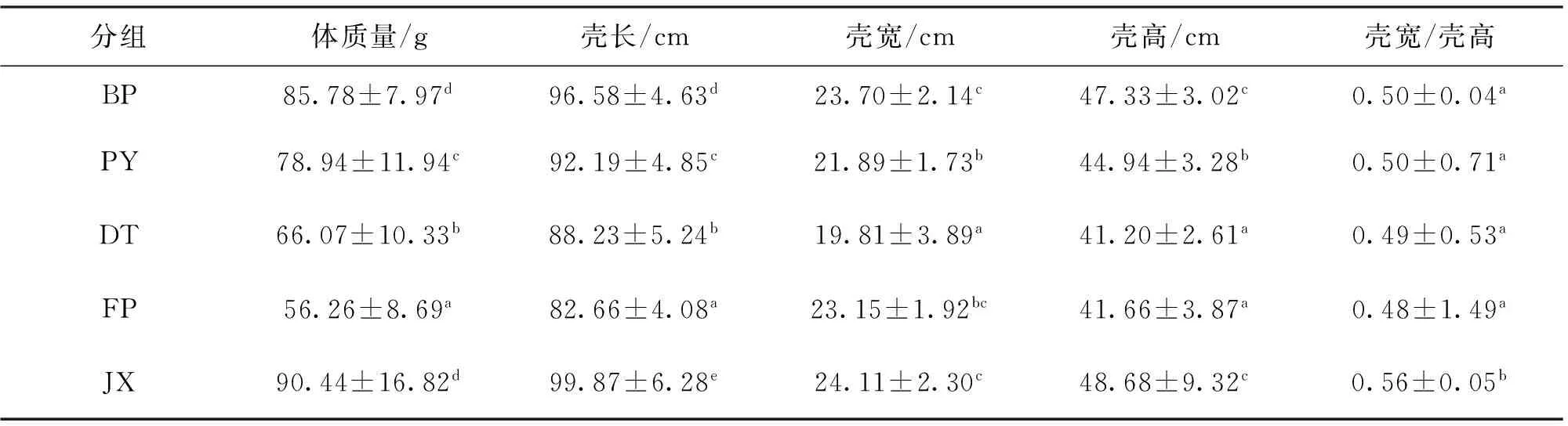
表3 5群体2龄蚌生长性能差异比较分析Tab.3 Comparison of growth traits among five populations
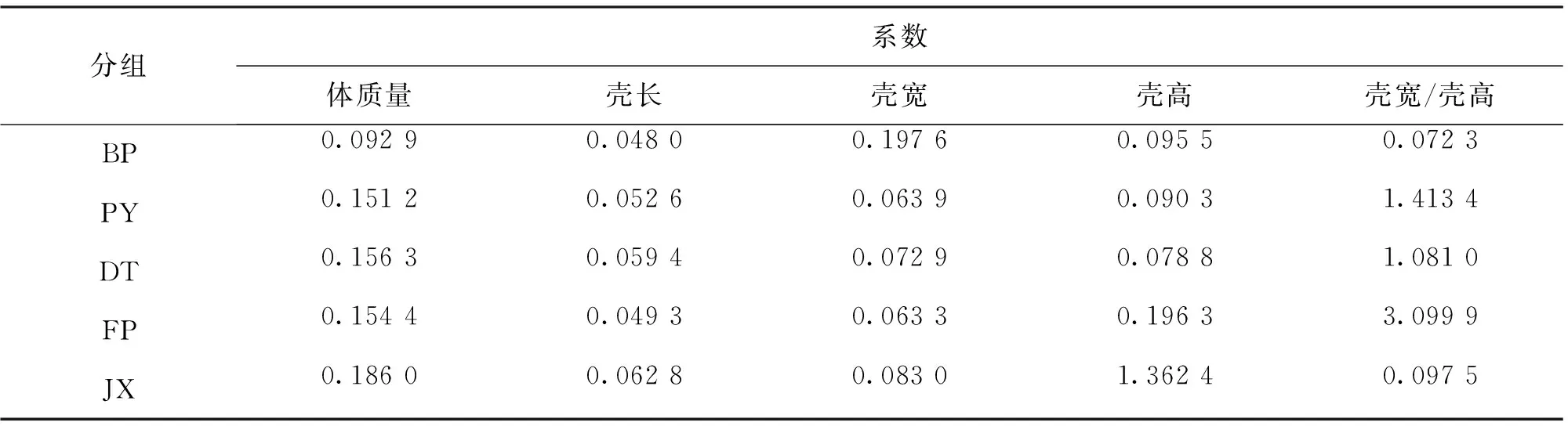
表4 5群体2龄蚌生长性状变异系数比较Tab.4 Comparison of variation coefficient of growth traits among five populations
2.2 生长性状之间相关性
如表5所示,通过对1龄、2龄蚌生长性状之间的相关性分析发现,各性状间均存在显著相关性,各长度指标与体质量相关程度由大到小为壳长、壳宽和壳高,1龄蚌壳宽与壳高的比例与体质量和壳长无显著相关性。与1龄蚌相比,2龄蚌壳宽与体质量的相关性越大。
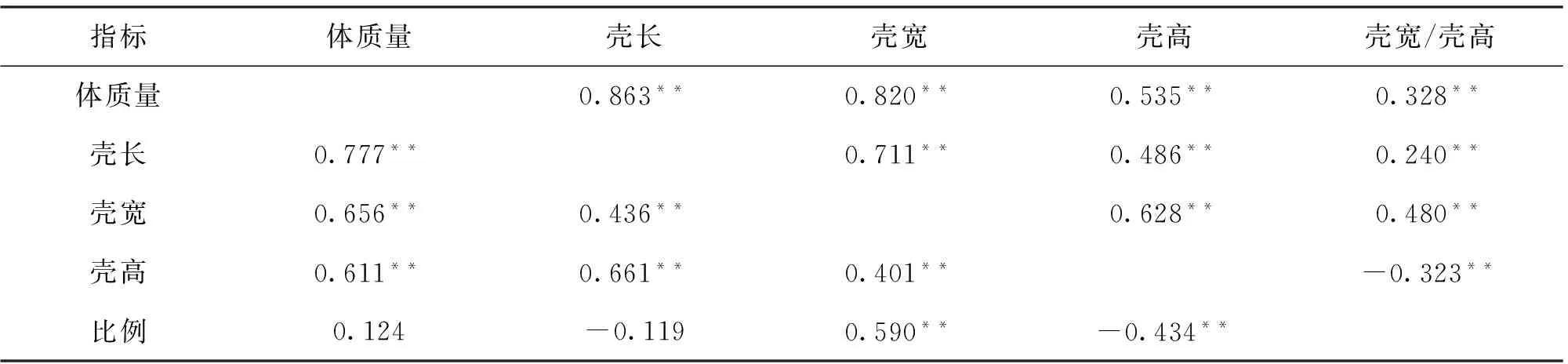
表5 5群体各生长指标Spearman相关性分析Tab.5 Spearman correlation coefficient of growth traits among five populations
注: 对角线以下是1龄阶段生长性状的相关性;对角线以上是2龄阶段生长性状的相关性;**表示在 0.01水平上差异显著.
2.3 聚类分析
如图1所示,对所有样本的10个比例性状进行聚类分析,得出4个三角帆蚌群体以及池蝶蚌群体形态聚类图。结果表明,PY与FP之间遗传距离最近,外形相似度最高,而DT、JX、BP与上述2个群体的趋异程度逐渐增加,其中BP与其它4个群体的遗传距离最远,表型差异最大。
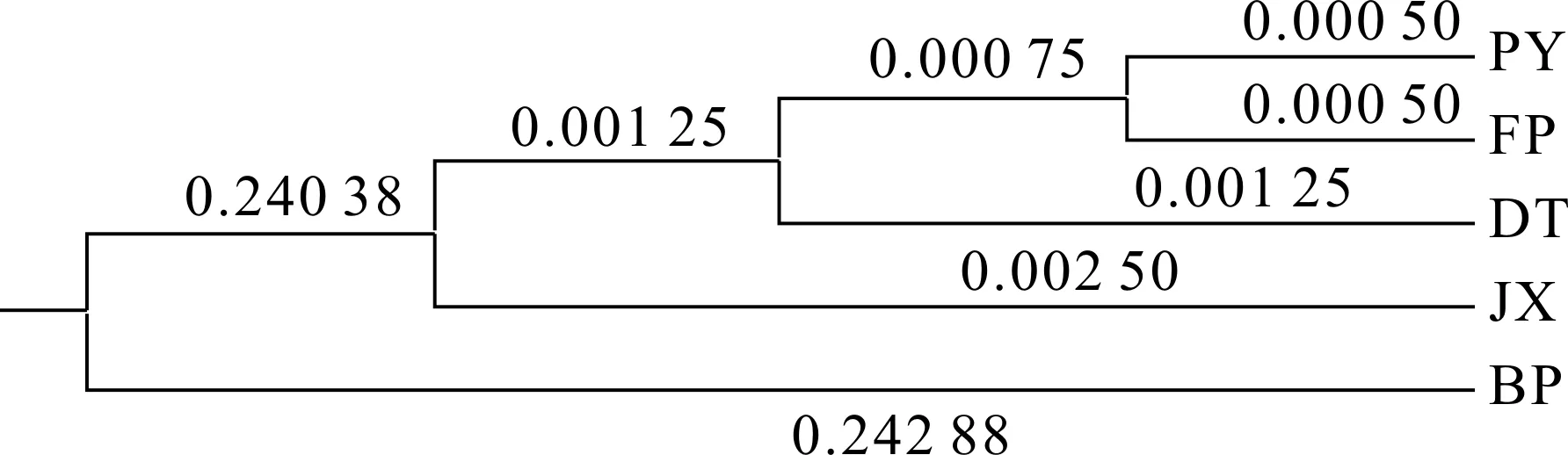
图1 基于外部形态变异聚类树Fig.3 Clustering tree based on the morphological variation
2.4 主成分分析
主成分分析结果见表6。4个主成分的累积贡献率为69.67%,数值较低,说明用几个相互独立的因子难以概括不同种群之间的形态差异。5个群体第1主成分中,全高/AB、FB/AB以及FA/AB的影响最大,从符号可以看出各个原始性状对该复合性状都是正作用,从绝对值大小可以看出第1主成分主要反映的是纵向生长。第2主成分中,OD/AB的影响最大,主要反映的是横向生长。
图2所示为5个群体第1、2主成分的散点图。图中“主成分1”和“主成分2”为相对值(无量纲)。由图2可以看出,BP与其它4个地理群体重叠的程度较小,而DT、PY、FP以及JX存在较多的重叠区域,难以区分,说明4个地理群体在外部形态上很相似,主成分分析的结果与聚类分析的结果相一致。
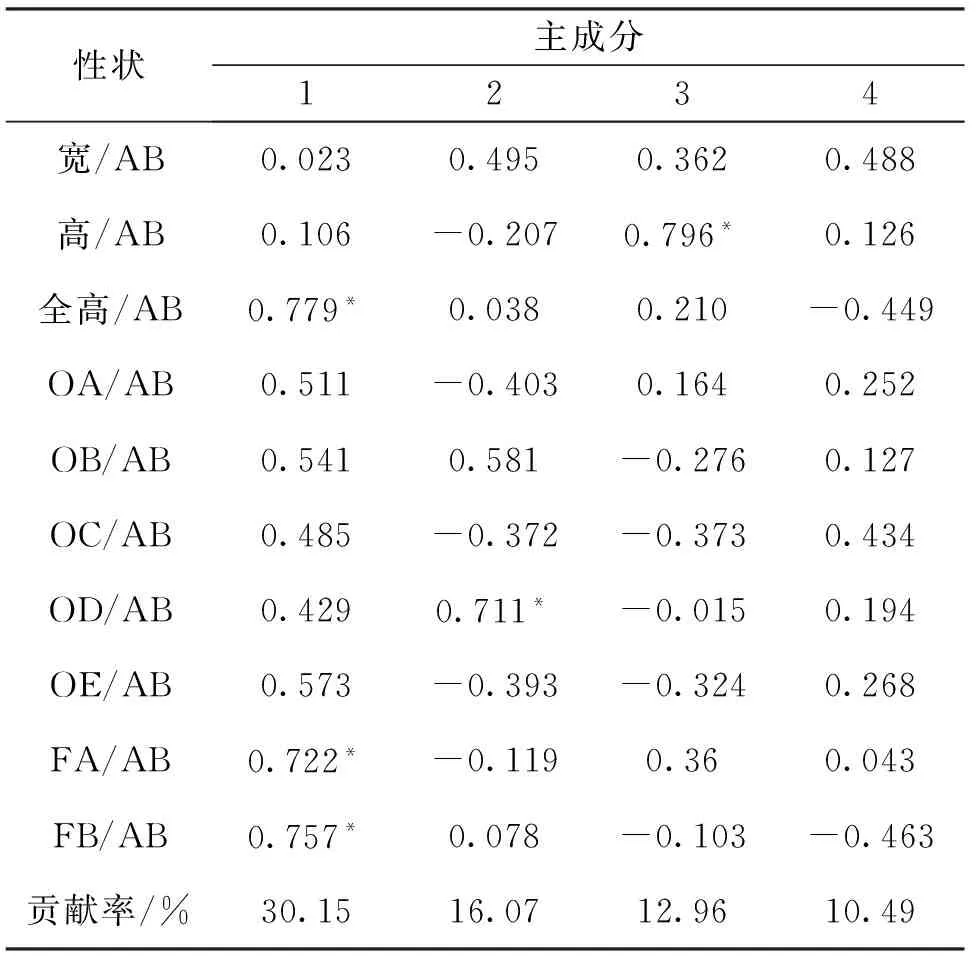
表6 5群体l0个性状的4个主成分的贡献率及负荷值Tab.6 Loading of four principal components for ten characters of five populations
*表示负荷值>0.700
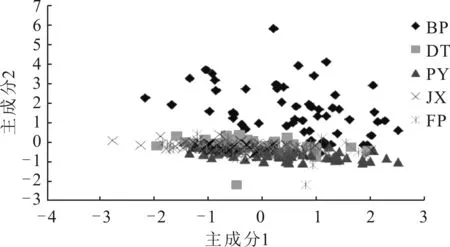
图2 5群体第1、2主成分的散点图分布Fig.2 Scatter diagram for the first and the secondprincipal components of five populations
2.5 判别分析
采用逐步判别的方法对5群体的10组标准化的特征值进行判别分析。通过逐步判别分析,使用9个性状的特征值(去除OA特征值)建立了5种蚌的判别函数,公式中的X1、X2、X3、X4、X5、X6、X7、X8、X9分别代表宽/AB、高/AB、全高/AB、OB/AB、OC/AB、OD/AB、OE/AB、FA/AB、FB/AB共9个比例性状,各函数的自变量系数和常数项见表7。
YBP= 97.571X1+883.670X2+870.637X3
-466.950X9-1 575.590
YPY=137.293X1+879.748X2+878.637X3
-506.648X9-1 598.334
YDT=151.543X1+854.535X2+928.112X3
-504.178X9-1 605.958
YFP=172.883X1+872.671X2+888.548X3
-514.767X9-1 607.165
YJX=128.294X1+906.133X2+904.891X3
-513.986X9-1 605.006
将9个参数的校正值分别代入上述判别函数中,比较函数值的大小,被判个体归属于函数值最大的那个函数所对应的种群或群体。为验证判别公式的实用性,对测量样本按上述判别公式进行预测分类,结果见表8。判别准确率P1为58.33%~73.33%,判别准确率P2为48.61%~100%,综合判别率为66.00%。
3 讨论
3.1 5群体生长性能差异和生长指标相关性
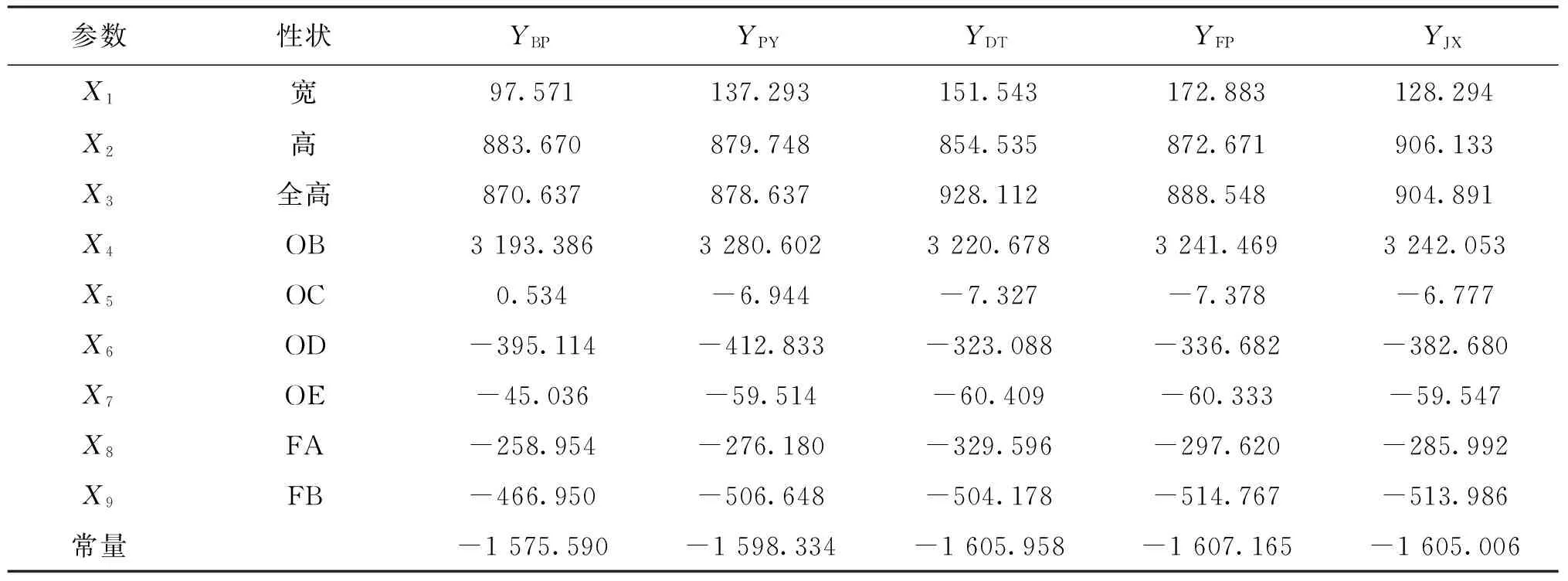
表7 5种群蚌壳外部形态判别公式及常数项Tab.7 The coefficients and constants of discriminant functions of the five populations of Hyriopsis
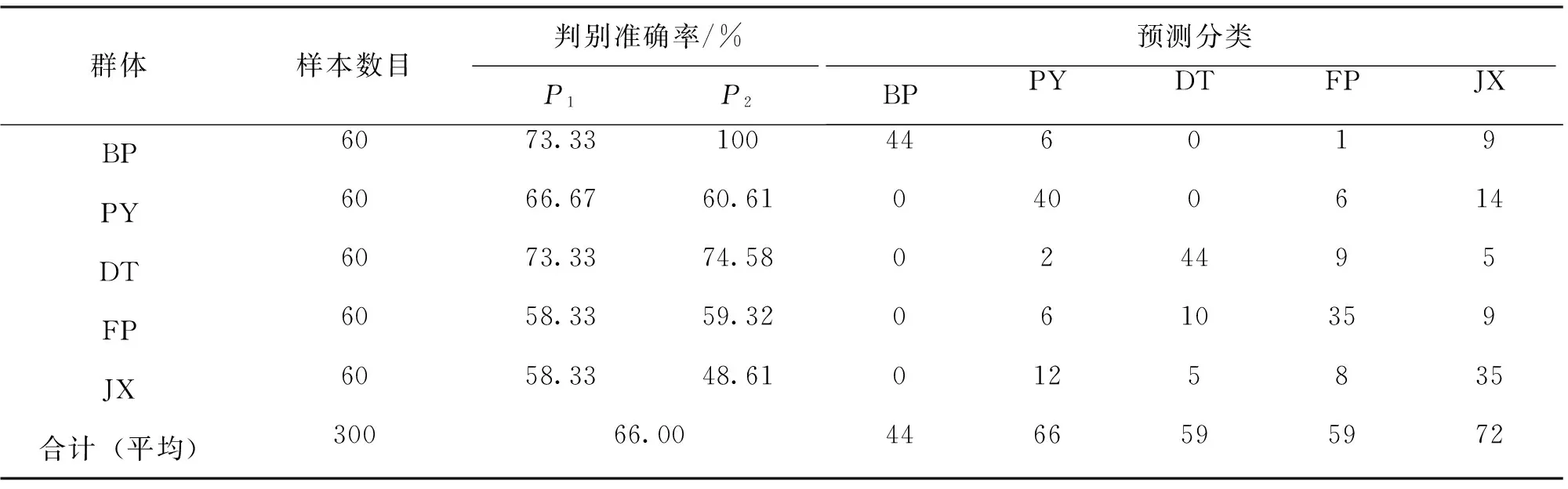
表8 5群体判别分析测试结果Tab.8 The discriminant results of five populations of Hyriopsis
对于育珠蚌养殖业而言,提高育珠蚌生长速度,尽快达到插片规格,是节省养殖成本、缩短资金流通周期及降低养殖风险的有效方法。Wada[8]认为育珠蚌亲本的选择应主要以生长性能、贝壳外部形态、珍珠质层颜色等指标为主。本研究通过对5个淡水珠蚌群体1至2龄的生长性能比较分析,发现5群体整体生长性能由大到小为JX、BP、PY、DT、FP。JX生长性能具有明显的优势,在壳宽与壳高比例性状方面尤为突出,显著高于其它几个群体。周春花等[9]通过对三角帆蚌与池蝶蚌几个生物学性状比较后获得类似的研究结果,认为从2龄开始,池蝶蚌的壳宽增长明显快于三角帆蚌。综上可见,池蝶蚌是优良的淡水育珠蚌资源,而经遗传改良的三角帆蚌选育群体在生长性能上也获得了明显的提高,两者具有较高的生产应用潜力。
贝类选育工作中,需要选择简便、准确、相关性强的性状指标作为判断标准,方能在生产实践过程中起到良好的效果[10]。郑怀平等[11]研究发现海湾扇贝的壳色与生长性状间有密切的联系,认为可以通过质量性状和长度性状的协同选择,培育出既美观又生长快的海湾扇贝特色品种。水产动物长度性状相对质量性状更稳定且易于测量,体质量更受性别和发育阶段(尤其是繁殖季节性腺组织)的影响[12]。本研究发现,1龄及2龄蚌的体质量与壳长、壳宽、壳高之间呈显著正相关,因此可通过测量壳长性状来反映个体生长情况,此结果与Jin等[13]研究结果一致。与1龄蚌相比,2龄蚌壳宽与体质量的相关性由显著相关转为高度相关,影响1龄蚌体质量的主要长度性状为壳长,影响2龄蚌体质量的长度性状主要为壳长和壳宽。王照棋等[14]研究认为三角帆蚌生长性状具有高等遗传力,通过选育能够获得良好的遗传效果。因此,通过长度性状对质量性状的间接选择,预计在淡水珠蚌的遗传改良中能取得较好的育种进展。
3.2 基于3种多元分析方法的群体蚌壳外部形态变异分析
本研究通过测定11个形态性状,通过校正,获得10个比例性状,在此基础上运用3种多元分析方法,对5个不同生态淡水珍珠蚌群体的形态性状进行分析,结果表明,5群体间形态差异很难用几个相互独立的因子来概括。
聚类分析可以对不同品系进行初步归类,量化家系间的差异程度,形成一个亲疏关系谱系图,直观地显示分类对象的差异和联系[15-17]。聚类分析结果表明,养殖群体与鄱阳湖群体首先聚类,关系紧密,推测可能与养殖户采用的亲本多来源于鄱阳湖有关。两者与洞庭湖群体较为接近,因为鄱阳湖、洞庭湖地理种群均隶属于长江水系,两群体间可能存在遗传交流,使其外部形态差异程度较低。在鱼类及贝类的研究中发现,形态学表型的差异建立在一定的遗传基础之上,同时与生存环境也密切相关[18-20]。Laudien等[21]认为大西洋浪蛤(Spisulasolidissima)形态学上的差异是由地理环境特性决定的[22-23],而群体间的遗传相似性则是基因交流、选择压力和进化习惯造成的。选育群体与其它几个群体的趋异程度最大,并最后聚类,这种差异可能主要源于人为选择压力以及特定养殖环境导致的。
主成分分析可将多个形态比例性状综合成少数几个因子,并可根据不同群体的主成分值,找出在形态上差异较大的参数。三角帆蚌与池蝶蚌均是我国主要的淡水育珠蚌,相关研究通过对其形态[24]以及同工酶[25]进行分析,结果表明两者之间的亲缘关系紧密,也有研究认为两者可能是同一个种的不同地理种群[26]。周春花等[9]通过若干生物学性状比较研究发现,池蝶蚌与三角帆蚌早期外部形态较为相近。本文主成分分析结果显示,JX与三角帆蚌群体(PY、DT、FP)之间存在较大的重叠区域,说明他们在外部形态上有相当程度的相似性,与以往研究结果一致。值得注意的是,BP主成分散点图分布区域与其它几个群体存在明显的区分,重叠区域较少,可能是由于继代选育,对优良表型的不断巩固稳定造成的。
判别分析是种群鉴别的最常用的方法,判别分析可以建立简单的公式对群体进行鉴别。判别分析结果表明,判别准确率P1为58.33%~73.33%,P2为48.61%~100%,综合判别准确率仅为66.00%。类似的情况在其他双壳类也有相关报道,么宗利等[27]在青蛤(Cyclinasinensis)的形态差异分析中综合判别率为67.50%,刘建勇等[28]对我国南海区4个真曲巴非蛤种群(Paphiaeuglypta)形态差异进行了分析,综合判别率为64.70%。研究获得较低判别准确率,一方面可能是由于这种形态判别为种内判别[29-30],另一方面可能是各群体间亲缘关系较近造成的。
以往人们主要利用壳宽相对于壳长的比值以及壳顶在铰合部中的前后位置两处作为蚌类鉴定的数量性状,辅助贝壳颜色、壳饰等属性性状进行描述统计,但对形态相似的种类和种群的鉴别难以实现[31]。众所周知,表型值是遗传和环境共同作用的结果,尽管选育群体与鄱阳湖、洞庭湖野生基础群体之间在外部形态方面发生了明显的变异,但基于外部形态差异进行区分其余4个群体,对于不同个体的形态判别准确率较低。因此,尽管形态差异分析适用于研究群体间遗传变异,但基于外部形态判别对不同种群间的个体种质鉴定仅可作为参考,在实践过程中仍建议借助生化、分子标记进行研究,获得更准确的鉴别。
[1]袁晓泉.池蝶蚌与三角帆蚌育珠性能比较研究[D].武汉:华中农业大学,2008.
[2]金 梅.三角帆蚌与池蝶蚌育珠性能比较[J].当代水产,2006,31(9):29-29.
[3]赵亚辉.中国特有洞穴金线鲃属(鲤形目:鲤科)鱼类的研究—物种多样性、系统演化和动物地理[D].中国科学院动物研究所,2005.
[4]Pinheiro A,Teixeira C M,Rego A L,et al.Genetic and morphological variation of Solea lascaris (Risso,1810) along the Portuguese coast[J].Fish Res,2005,73(2):67-78.
[5]魏开建,熊邦喜,赵小红,等.五种蚌的形态变异与判别分析[J].水产学报,2003,27(1):13-18.
[6]钱荣华,李家乐,董志国.中国五大湖三角帆蚌形态差异分析[J].海洋与湖沼,2003,34(4):436-443.
[7]罗 会,刘宝锁,黎火金,等.合浦珠母贝不同地理种群的形态差异和判别分析[J].广东农业科学,2013,40(12):171-174.
[8]Wada K T.Bivalve broodstock development in Japan[J].World Aquacult,1993,24:54-57
[9]周春花,徐毛喜,欧阳珊,等.池蝶蚌(HyriopsisSchlegeli)与三角帆蚌(H.Cumingii)若干生物学性状比较研究[J].江西科学,2003,21(2):122-124.
[10]王爱民,石耀华,周志刚.马氏珠母贝不同地理种群内自繁和种群间杂交子一代形态性状参数及相关性分析[J].海洋水产研究,2004,25(3):39-45.
[11]郑怀平,许 飞,张国范,等.海湾扇贝壳色与数量性状之间的关系[J].海洋与湖沼,2008, 39(4):328-333.
[12]斯烈钢,富 裕,李鸿鹏,等.四明山脉溪流区水产经济动物棘胸蛙(Paaspinosa)成体形态性状对体质量和净体质量影响效应的性别差异[J].海洋与湖沼,2015,46(3):679-686.
[13]Jin W,Bai Z,Fu L,et al.Genetic analysis of early growth traits of the triangle shell mussel,Hyriopsiscumingii,as an insight for potential genetic improvement to pearl quality and yield[J].Aquacult Int,2012,20(5):927-933.
[14]王照旗,韩学凯,白志毅,等.三角帆蚌紫色选育系1龄阶段内壳色及生长性状的遗传参数估计[J].水产学报,2014,38(5):644-650.
[15]唐守正.多元统计分析方法[M].北京:中国林业出版社,1990.
[16]蔡庆华.武汉东湖浮游植物水华的多元分析[J].水生生物学报,1990,14(1):22-31.
[17]李勤生,蔡庆华,华 俐,等.东湖异养细菌群落的分类结构和聚类分析[J].水生生物学报,1991,15 (3):242-253.
[18]Salini J P,Milton D A,Rahman M J,et al.Allozyme and morphological variation throughout the geographic range of the tropical shad,hilsaTenualosailisha[J].Fish Res,2004,66(1):53-69.
[19]Susanna P,Jorma P.Morphological differentiation among local trout (Salmotrutta) populations[J].Biological J Lin Soc,2001,72(2):231-239.
[20]Swain D P,Foote C J .Stocks and chameleons:the use of phenotypic variation in stock identification[J].Fish Res,1999,43(13):113-128.
[21]Laudien J,Flint N S,Bank F H V D,et al.Genetic and morphological variation in four populations of the surf clam Donax serra,(R?ding) from southern African sandy beaches[J].Biochem Syst Ecol,2003,31(7):751-772.
[22]Kong L,Li Q,Qiu Z.Genetic and morphological differentiation in the clam Coelomactra antiquata, (Bivalvia:Veneroida) along the coast of China[J].J Exp Mar Biol Ecol,2007,343(1):110-117.
[23]Plejdrup J K,Simonsen V,Pertoldi C,et al.Genetic and morphological diversity in populations of Nucella lapillus (L.;neogastropoda) in response to tributyltin contamination[J].Ecotoxicol Environ Saf,2006,64(2):146-154.
[24]郑汉丰,张根芳,李家乐,等.三角帆蚌、池蝶蚌及其杂交F1代早期形态差异分析[J].上海水产大学学报,2005,14(3):225-230.
[25]韩 庆,鲁密芳,张建平,等.池蝶蚌与三角帆蚌不同组织SOD及EST同工酶的比较研究[J].湖北农业科学,2013,52(15):3615-3618.
[26]Sakai H,Ujiie M,Mizutani E.Mlozyme comparison between Japanese and Chinese limnetic pearl mussels[J].J Nati Fish Univ,1997,46(2):23-39,101-104.
[27]么宗利,王 慧,周 凯,等.我国5个青蛤地理群体形态差异分析[J].海洋水产研究,2007,28(2):63-70.
[28]刘建勇,孙成波,李 文,等.我国南海4个真曲巴非蛤自然种群的形态差异分析[J].热带生物学报,2010,1(1):72-77.
[29]冯建彬,李家乐,王美珍,等.我国四海区不同群体文蛤形态差异与判别分析[J].浙江海洋学院学报,2005,24(4):318-323.
[30]张永普,林志华,应雪萍,等.不同地理种群泥蚶的形态差异与判别分析[J].水产学报,2004,28 (3):339-342.
[31]魏开建.中国蚌科的遗传多样性与系统发育的研究[D].武汉:华中农业大学,2004.
(责任编辑:邓 薇)
Comparative analysis of growth trait and morphological difference among five freshwater pearl mussel populations
WU Lei-ming1,BAI Zhi-yi1,2,LIU Xiao-jun1,2,JIN Can1,YIN Hao1,LI Jia-le1,2
(1.KeyLaboratoryofFreshwaterFisheryGermplasmResources,MinistryofAgriculture,ShanghaiOceanUniversity,Shanghai201306,China;2.ShanghaiEngineeringResearchCenterofAquaculture,ShanghaiOceanUniversity,Shanghai201306,China)
Hyriopsiscumingiiincluding breeding population F4,Wuyi cultured population,wild populations from Poyang Lake and Dongting Lake,as well asH.schlegeliipopulation were collected,and were bred synchronously.In this paper,1 and 2 year-old mussel growth performance of 5 groups offspring were analyzed,and three multivariation analysis methods (cluster analysis,principal component analysis and diseriminant analysis) were used to comparably investigate the morphological variations among the five populations.The results showed that the growth performances of five strains,from high to low were: the strain ofH.schlegelii,the selected strain,the Poyang Lake strain,Dongting Lake strain,and the cultured strain.The correlations for the growth indices were extremely significant,and the degrees of association with body quality,from large to small,were: the shell length,shell width and shell height.At 1 year-old,the impact of pearl mussel body mass index (BMI) mainly affected the shell length,and at 2-years old,the BMI mainly affected the shell length and shell width.The shell width and body mass correlation increased in the late pearl mussels,and the shell height and body mass correlation was significantly reduced.Three kinds of multivariate analysis showed that the external morphology of 1 year-old strain ofH.schlegelii,the Poyang Lake strain,the Dongting Lake strain and the cultured strain were very similar,whereas the external morphology of the selected strain varied significantly compared with the other four strains.
Hyriopsisschlegelii;Hyriopsiscumingii;growth trait;morphological difference
2016-04-06;
2017-04-11
国家自然科学基金项目(31272657);国家科技支撑计划课题(2012BAD26B04)
吴雷明(1987-),男,博士,专业方向为水产动物种质资源与种苗工程。E-mail: leiming4854@163.com
李家乐。E-mail: jlli2009@126.com
S966.2
A
1000-6907-(2017)04-0075-08
猜你喜欢
四川蚕业(2022年2期)2022-11-19
今日农业(2022年15期)2022-09-20
湖南电力(2021年1期)2021-04-13
世界科学技术-中医药现代化(2020年2期)2020-07-25
科学大众(2020年10期)2020-07-24
四川蚕业(2020年3期)2020-07-16
当代陕西(2019年6期)2019-04-17
红土地(2018年7期)2018-09-26
云南中医学院学报(2012年2期)2012-07-31
中国火炬(2012年8期)2012-07-25
