
番茄果实的芳香物质组成及其影响因素研究进展
2017-09-09 05:37王利斌李雪晖石珍源白晋和金昌海罗海波郁志芳
食品科学 2017年17期
王利斌,李雪晖,石珍源,白晋和,金昌海,罗海波*,郁志芳*
(1.扬州大学食品科学与工程学院,江苏 扬州 225127;2. 美国农业部园艺研究实验室,美国 佛罗里达 皮尔斯堡 34945;3.南京农业大学食品科技学院,江苏 南京 210095;4.南阳师范学院生命科学与技术学院,河南 南阳 473061;5.扬州大学 教育部植物功能基因组学重点实验室,江苏 扬州 225009;6.浙江医药高等专科学校食品学院,浙江 宁波 315100)
番茄果实的芳香物质组成及其影响因素研究进展
王利斌1,2,3,李雪晖4,石珍源5,白晋和2,金昌海1,罗海波6,*,郁志芳3,*
(1.扬州大学食品科学与工程学院,江苏 扬州 225127;2. 美国农业部园艺研究实验室,美国 佛罗里达 皮尔斯堡 34945;3.南京农业大学食品科技学院,江苏 南京 210095;4.南阳师范学院生命科学与技术学院,河南 南阳 473061;5.扬州大学 教育部植物功能基因组学重点实验室,江苏 扬州 225009;6.浙江医药高等专科学校食品学院,浙江 宁波 315100)
香气是番茄果实重要的感官品质之一,由果实中400多种具有挥发性的芳香物质协同产生,在很大程度上决定了商品等级和消费者的接受度。本文简述了番茄果实中的芳香物质及其在成熟过程中的变化趋势,总结了其合成途径,并重点介绍了影响番茄果实中芳香物质形成的因素,以期为研究者、生产者、销售者和消费者提供理论和生产实践方面的指导。
番茄香气;芳香物质;变化趋势;合成途径;影响因素
王利斌, 李雪晖, 石珍源, 等. 番茄果实的芳香物质组成及其影响因素研究进展[J]. 食品科学, 2017, 38(17): 291-300. 7DOI:10.7506/spkx1002-6630-201717047. http://www.spkx.net.cn
WANG Libin, LI Xuehui, SHI Zhenyuan, et al. Recent advances in research on volatile aroma compounds in tomatoes and their impacting factors[J]. Food Science, 2017, 38(17): 291-300. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201717047. http://www.spkx.net.cn
番茄(Lycopersiconesculentum Mill.)是茄科(Solanoideae)番茄属(Lycopersicon)的半直立或蔓生草本植物[1],其果实营养丰富且风味独特,在人类饮食中占有不可或缺的地位[2-3]。科研人员对番茄果实的产量、外观及耐贮性等方面进行了大量研究,培育出了高产耐贮的品种,但消费者发现这些品种的风味明显下降[4]。随着社会经济的发展和人们生活水平的提高,消费者愈发重视番茄的风味品质,也愿意为高品质番茄溢价支付[5]。
香气是番茄风味的重要组成部分,由多种具有挥发性的芳香物质组成,在很大程度上决定了消费者对番茄果实的偏爱程度和购买欲[6]。在番茄果实成熟过程中,目前已鉴定出的芳香物质有400多种,其中对番茄香气有重要贡献的约20~30余种。近年来,随着风味化学、基因组学和代谢组学的不断发展,有关芳香物质的研究取得了很大的进展。
本文简述了构成番茄香气的芳香物质及其在果实成熟过程中的变化趋势,进一步总结了其合成路径,并从基因型和环境因素等不同方面入手,重点介绍了影响番茄果实中芳香物质形成的因素,以期为研究者、生产者、销售者和消费者提供理论和生产实践方面的指导。
1 番茄果实中芳香物质及其在成熟过程中的变化趋势
番茄果实的成熟是一个复杂的、受遗传和生长发育等诸多因子调控的过程。在该过程中,会发生各种生理生化反应和结构的变化,影响果实的外观、质地和风味[7]。美国农业部依据番茄果实表面的颜色变化将成熟过程分为6 个时期,包括绿熟期、破色期、转色期、粉红期、微红期和红熟期[8]。
在番茄果实成熟过程中,已有超过400 种芳香物质被鉴定出来,主要包括醇类、醛类、酮类、酯类、萜类及含硫化合物等,其中16 种物质的对数阈值单位大于0且被认为对番茄香气有重要贡献[9](表1)。此外,研究显示,部分对数阈值单位为负的芳香物质如香叶醛、橙花醛、香叶基丙酮和愈创木酚等也会作为背景信息影响番茄香气[4,10-11]。

表 1 番茄果实中重要芳香物质含量、阈值浓度和对数阈值单位[9]Table 1 Important volatile aroma compounds in tomato fruits along with their contents, odor thresholds and log odor units[9]
在已鉴定出的400多种芳香物质中,部分组分(如己醛、顺-3-己烯醛和反-2-己烯醛等)在成熟过程中的变化趋势已有报道[11]。在“Solar Set”和“Sunny”番茄成熟过程中,丁香油酚含量逐渐下降,顺-3-乙烯醇、乙醛、己醛、顺-3-己烯醛、反-2-己烯醛、丙酮、6-甲基-5-庚烯-2-酮、香叶基丙酮和2-异丁基噻唑逐渐增加并在转色期和粉红期达到最大值[12]。课题组在研究过程中,发现“FL 47”番茄果实成熟过程中芳香物质的变化趋势(图1)。50%的组分在成熟早期积累并在红熟期激增,包括2-苯乙醇、3-甲基丁醇、3-甲基戊醇、6-甲基-5-庚烯-2-醇、2-甲基乙醛、丁醛、2-甲基丁醛、3-甲基丁醛、2-甲基-2-丁烯醛、顺-3-己烯醛、己醛、反-2-己烯醛、反,反-2,4-己二烯醛、苯甲醛、β-环柠檬醛、橙花醛、香叶醛、6-甲基-5-庚烯-2-酮、香叶基丙酮、假紫罗酮、丙酮、乙酸丙酯、D-柠檬烯、异戊腈、2-甲基呋喃(Ⅰ型);12%在成熟过程中逐渐上升,包括4-甲基戊醇、庚醛、1-戊烯-3-酮、乙酸异丁酯、2-甲基丁基乙酸酯和二甲基二硫醚(Ⅱ型);2-甲基丙醇、2-甲基丁醇、2-乙基己醇和β-紫罗兰酮在粉红期激增(Ⅲ型);壬醛、辛醛、2,3-丁二酮、2-乙基呋喃(Ⅳ型);戊醇、戊醛、反-2-辛烯醛、5-乙基-2(5H)-呋喃酮、1-辛烯-3-酮和水杨酸甲酯(Ⅴ型)在果实成熟过程中均先上升后下降;其余组分(沉香醇、壬烷、异丙基甲苯和愈创木酚)在成熟过程中保持不变(Ⅵ型)。该变化趋势与Klee等[13]的报道基本一致。尽管各组分在成熟过程中的变化趋势不尽相同,但大多数芳香物质在成熟后期达到最大值,其中就包括了许多对番茄香气有重要贡献的组分,如己醛、顺-3-己烯醛、反-2-己烯醛和6-甲基-5-庚烯-2-醇等(图1)。这一现象除了与番茄果实成熟期间芳香物质合成相关底物含量及酶活性增加有关外[11],也可能是因为在果实成熟过程中细胞结构发生改变,细胞膜通透性增加,破坏了酶和底物的区域化分布[13-14]。Klee[4]研究发现,机械伤可显著促进绿熟果实中C6芳香物质的形成,为上述推测提供了证据。
此外,研究表明芳香物质在成熟过程中的含量和变化趋势与基因型也密切相关。在“Sunny”番茄果实成熟过程中,1-戊烯-3-酮含量逐渐下降而其丰度则随着“Solar Set”果实成熟呈上升趋势;同时,在红熟“Solar Set”果实中顺-3-己烯醇、顺-3-己烯醛、乙醛、丙酮、6-甲基-5-庚烯-2-酮和丁香油酚含量也较前者更高[12]。


图 1 “FL 47”番茄果实成熟过程中50 种芳香物质的变化趋势Fig. 1 Variations in 50 volatile aroma compounds identifi ed in FL 47 tomato fruits during ripening
2 番茄果实中芳香物质的合成途径

图 2 番茄果实中重要芳香物质前体的起源[15]Fig. 2 Metabolic origins of the precursors of important volatile aroma compounds in tomato fruits[15]

表 2 番茄果实中与重要芳香物质合成有关的前体、酶和基因Table 2 Precursors, enzymes and genes related to the biosynthesis of important volatile aroma compounds in tomato fruits
目前,番茄果实成熟过程中已有超过400 种芳香物质被鉴定出来,部分重要组分的合成途径已逐渐清晰,许多关键的酶和基因已被克隆分离(表2)。芳香物质均起源于脂肪酸、类胡萝卜素、苯丙氨酸和支链氨基酸等[13],这些前体物质由CO2经卡尔文循环形成蔗糖后再经糖酵解、莽草酸途径和甲羟戊酸途径等不同代谢途径形成[15](图2)。接下来将依据不同芳香物质合成的前体逐一介绍其合成途径。
2.1 脂肪酸衍生物
C6芳香物质赋予番茄果实青草味和生青味[22],其合成途径如图3所示:亚油酸或亚麻酸依次经脂氧合酶C(lipoxygenase C,TomloxC)和13-氢过氧化物裂解酶(13-hydroperoxide lyase,13-HPL)和催化形成己醛和顺-3-己烯醛,而后在乙醇脱氢酶2(aldehyde dehydrogenase 2,ADH2)的作用下进一还原成相应的醇类[11](图3)。顺-3-己烯醛也可通过酶促或非酶促反应异构为反-2-己烯醛[15]。

图 3 番茄果实中C6芳香物质合成途径[11]Fig. 3 Biosynthesis pathway of C6 volatile aroma compounds in tomato fruits[11]
一般认为,TomloxC、13-HPL和ADH2均定位于有色体[13],其分别由TomloxC、13-HPL和ADH2基因编码[11]。在番茄成熟过程中,TomloxC的表达呈先升后降趋势,并在破色期后3 d达到最大值[23];而13-HPL和ADH2的表达量及相应酶活力逐渐增加并在红熟期达到最大值[24-25]。
C5芳香物质(戊醇、1-戊烯-3-醇、戊醛、反-2-戊烯醛和1-戊烯-3-酮)具有果香味,其含量高低与消费者对番茄的喜好密切相关[11,26]。最新研究发现,TomloxC也参与C5芳香物质的合成代谢[16]。
2.2 类胡萝卜素衍生物
番茄果实中,以类胡萝卜素为前体合成的芳香物质有香叶醛、橙花醛、6-甲基-5-庚烯-2-酮、香叶基丙酮、假紫罗酮、β-紫罗兰酮和β-大马酮(表2)。它们均具有果香味和芳香味,对番茄香气有重要贡献[4,11]。其中,香叶醛、橙花醛和假紫罗酮起源于番茄红素及其前体,而番茄红素、β-胡萝卜素和ζ-胡萝卜素分别是6-甲基-5-庚烯-2-酮、β-紫罗兰酮和香叶基丙酮的前体[22,27]。
LeCCD1A和LeCCD1B基因编码的类胡萝卜素裂解双加氧酶(carotenoid cleavage dioxygenases,LeCCDs)催化类胡萝卜素衍生物的形成[4,17],二者均具有底物广谱性和裂解位点专一性的特点。不仅可水解线型类胡萝卜素中5~6、7~8、9~10或13~14位点的双键,还参与环型类胡萝卜素中9~10位点双键的裂解[4,17]。研究发现,LeCCD1位于番茄叶绿体的外膜上[28]。在果实成熟过程中,LeCCD1A的表达量先降后升,在粉色期最低,而LeCCD1B mRNA含量逐渐上升,并在粉色期达到最大值[28]。
2.3 酚类物质(苯丙氨酸衍生物)
在番茄果实中,起源于苯丙氨酸的芳香物质包括2-苯乙醇、2-苯乙醛、水杨酸甲酯、愈创木酚、儿茶酚、丁香油酚、2-苯乙腈和1-硝基-2-乙基苯(表2)。
2-苯乙醇和2-苯乙醛赋予番茄果实花香味,而1-硝基-2-乙基苯具有土腥味,其合成过程如图4所示。苯丙氨酸经LeAADC1A、 LeAADC1B、LeAADC2基因编码的氨基酸脱羧酶(amino acid decarboxylases,LeAADCs)催化形成苯乙胺,随后在单胺氧化酶(monoamine oxidase,MAO)的作用下形成2-苯乙醛,抑或经过一系列反应最终生成2-苯乙腈和1-硝基-2-乙基苯[18]。2-苯乙醛可继续被苯乙醛还原酶(phenylacetaldehyde reductases,LePARs)还原形成2-苯乙醇[19],该酶由LePAR1和LePAR2基因编码[19]。在果实成熟过程中,LeAADC2、 LePAR1和LePAR2的转录效率逐渐降低而LeAADC1A和LeAADC1B的表达量不断变化并在红熟期最低[18-19]。
此外,苯丙氨酸也可先经苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)催化形成反肉桂酸,后者再经过不同的代谢支路形成水杨酸甲酯、愈创木酚、儿茶酚或丁香油酚[15](图4)。目前,参与该途径的部分基因及其产物也已被分离和鉴定,主要包括SlSAMT1基因编码的水杨酸甲基转移酶1(salicylic acid methyl transferase 1,SlSAMT1)[20]和CTOMT1基因编码的儿茶酚氧位甲基转移酶1(catechol-O-methyl transferase 1,CTOMT1)[29]。在果实成熟过程中,二者表达量均呈下降趋势[20,29]。水杨酸甲酯的薄荷味,儿茶酚和丁香油酚的丁香味,及愈创木酚的药味共同丰富了番茄果实的风味品质[30]。

图 4 番茄果实中酚类芳香物质(苯丙氨酸衍生物)合成途径[15]Fig. 4 Biosynthesis pathway of phenolic volatile aroma compounds (phenylalanine derivatives) in tomato fruits[15]
2.4 支链氨基酸衍生物
支链氨基酸衍生物包括2-甲基丁醇、3-甲基丁醇、2-甲基丁醛、3-甲基丁醛、乙酸异丁酯、异戊腈和2-异丁基噻唑(表2)。2-甲基丁醇、3-甲基丁醇、2-甲基丁醛和3-甲基丁醛均赋予果实焦糖味[11],乙酸异丁酯和异戊腈分别具有辛辣味和果香味[11];作为番茄独有的芳香物质,2-异丁基噻唑赋予番茄果香味和生青味[11]。在番茄果实中,它们以支链氨基酸(异亮氨酸和亮氨酸)为前体在支链氨基酸氨基转移酶(branched-chain amino acid aminotransferases,SlBCATs)参与下,经过脱氨、脱羧和还原等一系列反应形成[21]。
目前,在番茄果实中已鉴定出6 个支链氨基酸转氨酶基因,包括SlBCAT1、SlBCAT2、SlBCAT3、SlBCAT4、SlBCAT5和SlBCAT6[31]。其中,SlBCAT1和SlBCAT2位于线粒体中,参与支链氨基酸的分解代谢;SlBCAT3和SlBCAT4位于叶绿体中,参与支链氨基酸的合成代谢;SlBCAT5和SlBCAT6分别位于细胞溶质和液泡中[31]。在果实成熟过程中,SlBCAT1的表达量逐渐增加并在破色期达到最大值,而SlBCAT2、SlBCAT3和SlBCAT4的转录产物含量逐渐下降[31]。
3 影响番茄果实中芳香物质的因素
过去半个世纪,育种学家们对番茄果实产量、外观及耐贮性的过度重视,而忽视了其风味品质[4]。此外,不适宜的采前和采后处理也会影响果实中芳香物质组成[11](图5)。
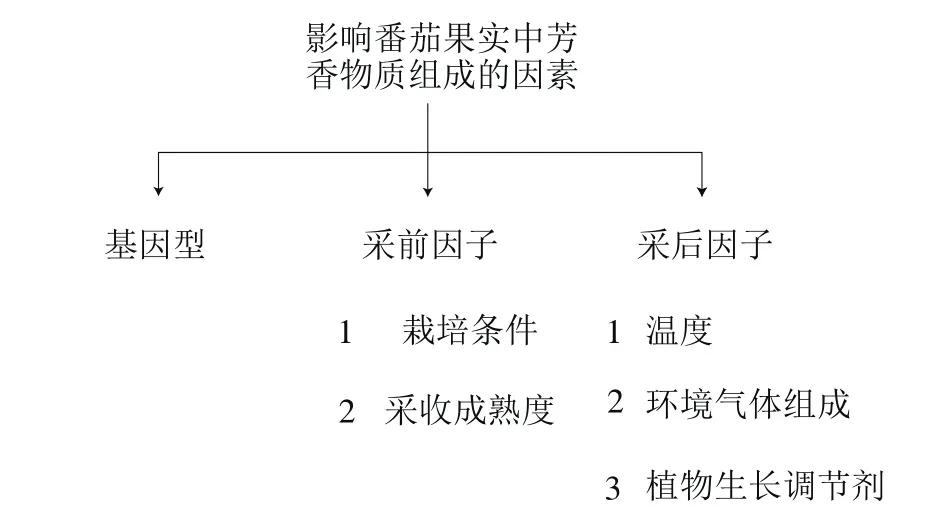
图 5 影响番茄果实中芳香物质组成的因素Fig. 5 Factors affecting volatile aroma composition in tomato fruits
3.1 基因型
基因型对番茄果实中芳香物质组成起决定性作用[11]。常培培等[32]对“红樱桃”、“粉南”、“紫香玉”、“金珠1号”和“绿宝石”5 个不同基因型樱桃番茄果实中芳香物质组成进行了研究。在5 个品种中共检测出81 种芳香物质,包括11 种烃类、19 种醇类、20 种醛类、10 种酮类、13 种酯类及8 种其他类别芳香物质;其中,乙醇、己醇、顺-3-己烯醇、己醛、反-2-己烯醛、反-2-辛烯醛、反,反-2,4-癸二烯醛、顺-2-庚烯醛、香叶基丙酮、法尼基丙酮、棕榈酸甲酯、棕榈酸乙酯、2-正戊基呋喃和2-异丁基噻唑为共有组分,构成“樱桃”番茄的主体风味。尽管均由醇类、醛类、酮类、酯类和烃类组成,但品种间芳香物质组成存在着明显差异:“红樱桃”、“粉南”、“紫香玉”、“金珠1号”、“绿宝石”果实中分别有46、45、44、38 种和34 种组分。“红樱桃”、“粉南”和“紫香玉”果实中酮类物质含量最多,醛类、醇类、酯类和烃类渐次递减;“金珠1号”果实中醛类物质含量最高,其次为醇类和酯类;“绿宝石”果实中醇类物质含量最高,其次为酯类[32]。就共有组分而言,乙醇和棕榈酸乙酯在“金珠1号”果实中含量最高,2-异丁基噻唑在“绿宝石”果实中含量最高,其余组分在“红樱桃”和“粉南”果实中含量最高[32]。Baldwin等[33]研究了佛罗里达州38 个不同基因型番茄果实中芳香物质组成,也发现了类似结果。
作为植物体内重要的内源激素和信号分子,乙烯参与调控果蔬的成熟衰老及抗逆过程[34]。研究显示,乙烯对番茄果实中芳香物质合成有重要的调控作用[11]。对“Ailsa Craig”番茄果实中1-氨基环丙烷-1-羧酸合酶基因进行干涉抑制表达,可显著抑制乙烯的合成,导致果实中甲醇、乙醇、2-甲基丁醇、3-甲基丁醇、顺-3-己烯醇、己醛、顺-3-己烯醛、反-2-己烯醛、顺-2-庚烯醛、丙酮、香叶基丙酮、1-戊烯-3-酮、6-甲基-5-庚烯-2-酮、2-异丁基噻唑和1-硝基-2-乙基苯含量降低[10]。干扰果实中乙烯信号传导也出现类似现象[11]。Klee等[13]研究发现,抑制乙烯不敏感突变体Nr,可使果实中部分芳香物质(如己醛、顺-3-己烯醛、反-2-己烯醛和6-甲基-5-庚烯-2-酮等)含量不再随贮藏时间的延长而增加。乙烯影响芳香物质合成可能是通过干扰合成途径中底物含量或关键酶活力来实现的[11]。
利用基因工程技术,直接对芳香物质合成途径中相关底物或酶活力进行改良或干扰,实现不同芳香物质在组分和含量上的多样性配伍,也可影响果实风味。有研究表明,ADH2基因过表达可显著增加ADH2酶含量,进而提高“AC”番茄果实中C6醇类芳香物质含量;感官评定发现转基因的果实更具有红熟果实的风味[35]。
3.2 采前因素
3.2.1 栽培条件
3.2.1.1 栽培方式
有研究表明,与温室栽培的相比,田间种植的“V. R. Moscow”番茄果实中3-甲基丁醇、2-甲基-3-己醇、3-甲基丁醛、糖醛、苯甲醛、乙酸异戊酯、水杨酸甲酯和α-蒎烯等芳香物质的含量较高[36]。采用混合模型分析,Cebolla-Cornejo等[37]发现田间种植的“CDP8908”、“CDP0916”、“CDP8075”、“CDP9944”、“Bond”和“Cambria”番茄果实中水杨酸甲酯含量较高而温室栽培的果实中己醛含量较高。
除了作为重要的香气组分外,水杨酸甲酯也可作为信号分子参与植物抗逆过程[38]。有研究表明,低温胁迫可诱导番茄果实中水杨酸甲酯合成[22]。因此,田间种植的果实中水杨酸甲酯含量较高可能是户外较温室温度低所致[37]。
3.2.1.2 栽培季节
由于不同栽培季节的温光条件也不尽相同,也会影响果实中芳香物质的合成[39]。杨明惠等[39]以“佳粉15号”番茄为试材,采用营养液栽培,研究了春茬和秋冬茬果实中芳香物质组成的异同。结果表明,不同栽培季节对番茄果实中芳香物质的种类和含量均有很大影响:在春茬果实中,可检测到48 种芳香物质,而在秋冬茬果实中仅检测到37 种,含量不足春茬的一半。其中18 种组分属二者共有,包括1-辛醇、2-辛醇、辛醛、己醛、壬醛、癸醛、反-2-庚烯醛、反-2-辛烯醛、反,反-2,4-癸二烯醛、α-柠檬醛、β-柠檬醛、1-戊烯-3-酮、6-甲基-5-庚烯-2-酮、二氢假紫罗酮、2,3-二氢-3,5-二羟基-6-甲基-4H吡喃-4-酮、2,6,6-三甲基-3-亚甲基-7-(3-氧亚丁基)氧杂环庚烷-2-yl乙酸酯、紫苏烯和2-异丁基噻唑。除2-辛醇、反-2-辛烯醛、壬醛、癸醛和紫苏烯外,其余组分在春茬果实中较高。
3.2.1.3 水肥供应
依据最佳管理措施的要求,在保证水质的前提下,为了提高番茄产量,在种植过程中要保证氮肥供应[40]。有研究显示,随着肥料中氮和钾含量的提高,“Walter”番茄果实中己醛、苯甲醛、2-苯乙醛、2-己酮、β-紫罗兰酮和6-甲基-5-庚烯-2-酮含量也逐渐增加[41]。肥料中硼含量的高低也会影响果实中芳香物质组成。“中杂101号”番茄果实在缺硼条件下,醇类、酮类、萜烯类较少,而在多硼环境中,除上述物质外,烃类、醛类、酸类和呋喃环类均有不同程度的提升[42]。
此外,采前多雨天气会降低番茄果实中芳香物质的含量[10]。增施CO2,改变培养液中锰、锌或铁的含量也会影响果实中芳香物质组成[43-46]。在这些研究中,改变肥料中不同营养素含量对果实中芳香物质组成影响各异,这可能是因为各营养素对细胞不同代谢途径影响不尽相同。
3.2.2 采收成熟度
为了保证风味品质,番茄应在红熟期采收[11],但供应商为了延长物流时间,通常在成熟早期进行采摘,这会对果实的风味品质产生影响[47]。就“FL 47”番茄而言,与绿熟期采收的相比,破色期采收的果实中甲醇、顺-3-己烯醛、反-2-己烯醛、反-2-庚烯醛、6-甲基-5-庚烯-2-酮、β-大马酮和2-异丁基噻唑的含量较高,而2-甲基丁醇、3-甲基丁醇、己醛、丙酮和水杨酸甲酯较低[48]。同时,红熟期采收的果实(品种:“Cal Ace”、“Cherry”、“Calmart”和“Early pak 7”)比成熟早期采收的果实具有更浓郁的番茄香气[49]。
尽管未成熟绿期采收的果实也能后熟,但与绿熟期采收的相比,果实中芳香物质组成发生了改变。以“Solimar”番茄为例,未成熟绿期采收的果实中顺-3-己烯醛、1-戊烯-3-酮、6-甲基-5-庚烯-2-酮、香叶基丙酮和2-异丁基噻唑含量较低,电子鼻可将它们区分开[50]。
3.3 采后因素
3.3.1 温度
低温冷藏是维持果蔬采后品质的有效手段,但不适宜的低温会对果蔬造成伤害[51]。将绿熟期番茄果实于10 ℃贮藏14 d或于5 ℃贮藏6~8 d,影响果实后熟过程中的一系列的生理生化反应,影响果实的品质[52]。常见的冷害症状包括不能正常成熟,风味损失、提前软化、表面颜色不均、果肉呈水浸状、种子褐变和对链格孢属真菌敏感等[52]。其中,风味损失发生在果实产生可见冷害症状之前[53]。研究发现,5 ℃冷藏破色期“FL 47”番茄9 d后,虽然并未导致在后熟过程中可见冷害症状的出现,但红熟期果实中芳香物质总含量降低;同时,低温抑制了2-苯乙醇、3-甲基丁醇、2-苯乙醛、2-甲基丁醛、3-甲基丁醛、1-戊烯-3-酮、6-甲基-5-庚烯-2-酮和2-异丁基噻唑在内的芳香物质[22]。冷藏绿熟期或红熟期番茄果实也会发现类似结果[54-55]。研究发现不同处理时期对红熟果实中芳香物质组成影响各异[54-55](表3),这可能是由于不同时期果实对低温敏感程度各异[53]。此外,品种、处理温度和时间也会影响果实中芳香物质组成(表3)。Maul等[53]研究发现“BHN-189”番茄果实比“Solimar”果实对低温更加敏感,这可能是由于其基因型决定的。
对采后番茄果实进行热处理可延缓果实后熟进程、控制病虫害、延长贮藏期,具有简单、高效、无毒、无污染、环保健康等优点[56-57],但也会影响果实中芳香物质组成。50 ℃热水处理红熟期“FL 47”番茄果实5 min,抑制了2-苯乙醇、2-甲基丁醛、顺-3-己烯醛、反-2-己烯醛和2-苯乙醛等芳香物质形成[55](表3)。与低温处理相一致,热处理效果也受处理时期、温度和时间的影响[54-55,58]。

表 3 温度处理对番茄果实中芳香物质组成的影响Table 3 Impact of temperature treatment on volatile aroma composition in tomato fruits
一些学者对温度处理抑制C6芳香物质形成的调控机制进行了研究[57]。研究发现,低温冷藏和热处理以不同方式调控“Tasti-Lee”和“Sanibel”番茄果实中C6芳香物质合成:低温下调了HPL分支中13-HPL和ADH2的表达量,同时抑制了相应的酶活力;而热处理是通过抑制13-HPL和ADH2酶活力来实现[57]。
3.3.2 环境气体组成
气调贮藏是目前保鲜新鲜农产品最有效、最安全的方法之一。研究表明,采用自发气调贮藏“Solairo”番茄果实35 h抑制了己醛、甲基丁醛和2-异丁基噻唑的合成而乙醇含量显著提高[58]。与此类似,人工气调可提高“Bermuda”果实中乙醇和乙醛含量,减少“Tasti-Lee”果实中己醛、柠檬醛、β-紫罗兰酮、香叶基丙酮和6-甲基-5-庚烯-2-酮含量[59-60]。
3.3.3 植物生长调节剂的使用
植物生长调节剂是人们在了解天然植物激素的结构和作用机制后,人工合成的与天然植物激素具有类似生理和生物学效应的物质,具有稳产增产、改善品质、增强作物抗逆性等作用[61]。
水杨酸甲酯和茉莉酸甲酯均可作为信号分子参与番茄果实的成熟和抗逆过程[22,38]。研究发现,用0.05 mmol/L水杨酸甲酯或茉莉酸甲酯熏蒸破色期“FL 47”番茄果实24 h抑制了红熟果实中3-甲基丁醇、2-甲基丁醛、1-戊烯-3-酮、6-甲基-5-庚烯基-2-酮和香叶基丙酮等芳香物质合成[22](表4)。此外,水杨酸甲酯处理可提高果实中水杨酸甲酯和壬烷含量,而茉莉酸甲酯熏蒸则可促进2-苯乙醛合成[22](表4)。
利用外源乙烯处理番茄果实,除了可促进成熟外[62],也会影响果实中芳香物质组成。将绿熟期“Agriset”番茄用100 mg/L乙烯熏蒸24 h减少了红熟果实中甲醇、己醛、6-甲基-5-庚烯-2-酮、香叶基丙酮、2-异丁基噻唑和1-硝基-2-乙基苯等芳香物质含量[63](表4)。1-甲基环丙烯(1-methylcyclopropene,1-MCP)是一种乙烯受体抑制剂,能不可逆地作用于乙烯受体蛋白,阻断其与乙烯的正常结合,抑制其诱导的与果实后熟相关的一系列生理生化反应,延缓果实成熟和衰老,具有无毒、低量、高效、作用效果持久等优点[64]。有研究发现,500 nL/L 1-MCP熏蒸破色期“FL 47”番茄果实12 h促进了红熟果实中3-甲基丁醇、2-甲基丁醇形成[48](表4)。

表 4 外源植物生长调节剂的使用对番茄果实中芳香物质组成的影响Table 4 Impact of exogenous plant growth regulators on volatile aroma composition in tomato fruits
4 存在问题及展望
番茄香气是由果实内具有挥发性的芳香物质组成的复杂混合物,在消费者对番茄果实的接受和购买过程中起着重要作用。在已鉴定出来的400多种芳香物质中,约20~30余种组分对果实香气有重要贡献。然而,育种学家对于目标组分的选择标准依然存在疑问[15]。就植物生长调节剂对芳香物质的影响而言,尽管采后水杨酸甲酯或茉莉酸甲酯熏蒸影响了红熟果实中芳香物质组成,然而并未影响人类通过嗅觉系统主观上对整体香气的鉴定[22]。这种由嗅觉系统的复杂性、芳香物质间的互作等诸多因子共同作用的现象,为科研人员寻找目标组分提出了更为严苛的要求[15,22]。
随着基因组学和代谢组学的不断发展,番茄果实中部分芳香物质的合成途径已被阐明,催化其合成相关的酶和基因也已被挖掘和验证(表2),但对关键调控因子的了解还不深入。通过数量性状基因(quantitative trait loci,QTLs)分析发现ADH2酶活力是合成C6醇类芳香物质的限制性因子[21],这一点已通过转基因实验验证[35]。随着QTLs分析技术的持续发展,关联分析技术的广泛应用,通过基因芯片和转录组测序技术(RNA sequence,RNA seq)等高新技术去追踪番茄芳香物质代谢过程中的关键酶及调控因子,将有助于我们进一步克隆和分离相关基因,了解多种基因互作下芳香物质的代谢途径,从而培育出风味更好的番茄果实[11]。
虽然许多采前和采后因素均会影响番茄果实中芳香物质组成,但多数研究集中在组分含量变化方面,尚未深入挖掘其调控机理,这可能是与许多芳香物质的合成途径及其调控因子尚未被完全阐明有关。C6芳香物质合成途径是目前了解最为透彻的,有研究发现,采收成熟度、栽培季节及环境气体组成等均会影响其含量[11]。Bai Jinhe等[57]从基因和酶活角度探讨了温度处理调控C6芳香物质合成的机制。在低温胁迫下,植物会启动丙二烯氧化物合成酶(allene oxide synthase,AOS)途径合成茉莉酸[57],后者在植物抵御低温胁迫过程中起重要作用[65-67]。亚油酸和亚麻酸也是AOS支路的底物,但在低温胁迫下,番茄果实中HPL支路和AOS支路之间的互作还鲜有研究。就类胡萝卜素衍生物而言,其含量与在果实中类胡萝素组成密切相关[4],栽培季节、氮肥供应、温度处理及植物生长调节剂的使用等均会影响该类衍生物含量,目前,已有相关研究证实,低温会抑制类胡萝卜素的合成[68],然而其他因素对类胡萝卜素组成的影响还鲜有报道。此外有研究表明,LeAADCs是合成2-苯乙醛的关键酶,该酶的转录活性直接决定了植物中2-苯乙醛的含量[69-71]。尽管氮肥供应、温度处理及植物生长调节剂的使用等均会影响2-苯乙醛含量,但这些处理是否会影响LeAADC1ALeAADC1B和LeAADC2表达仍不清楚。另一方面,在植物体中PAL支路和AADC支路之间对底物存在竞争关系[72],但这两个代谢支路在采前和采后因素下的互作还鲜有研究。随着科学技术的不断发展以及基因组学、代谢组学以及QTLs分析技术等分子生物学技术的在采后生物学上的应用,这些问题将逐渐被解决,从而将有利于深入了解各种采前和采后因素调控芳香物质合成的机制。
随着风味化学、基因组学和代谢组学等研究技术的进一步发展,番茄果实中芳香物质的合成途径与调控机制将被进一步揭示,为筛选提高番茄香气的育种技术和采前/采后处理方式奠定扎实的理论基础。
[1] 密森. 番茄果实主要芳香物质含量和营养品质组成变化的研究[D].扬州: 扬州大学, 2012: 8.
[2] 刘明池, 郝静, 唐晓伟. 番茄果实芳香物质的研究进展[J]. 中国农业科学, 2008, 41(5): 1444-1451. DOI:10.3864/j.issn.0578-1752.2008.05.024.
[3] 谭石勇. 解淀粉芽孢杆菌T-5防控番茄土传青枯病的效果及机理研究[D]. 南京: 南京农业大学, 2013: 1.
[4] KLEE H J. Improving the flavor of fresh fruits: genomics, biochemistry, and biotechnology[J]. New Phytologist, 2010, 187(1): 44-56. DOI:10.1111/j.1469-8137.2010.03281.x.
[5] 陈素艳. 不同番茄品种糖酸代谢及 浙粉702 种子纯度的SSR鉴定[D].杭州: 浙江农林大学, 2014: 1.
[6] EL HADI M A, ZHANG F J, WU F F, et al. Advances in fruit aroma volatile research[J]. Molecules, 2013, 18(7): 8200-8229. DOI:10.3390/ molecules18078200.
[7] GIOVANNONI J J. Genetic regulation of fruit development and ripening[J]. Plant Cell, 2004, 16(Suppl 1): 170-180. DOI:10.1105/tpc.019158.
[8] United States Department of Agriculture. United States standards for grades of fresh tomatoes[EB/OL]. [2015-02-14]. http://www.ams.usda. gov/AMSv1.0/getfi le.
[9] ACREE T, TERANISHI R. Flavor science: sensible principles and techniques[M]. Washington: American Chemical Society, 1993: 259-286.
[10] BALDWIN E A, SCOTT J W, SHEWMAKER C K, et al. Flavor trivia and tomato aroma: biochemistry and possible mechanisms for control of important aroma components[J]. Hortscience, 2000, 35(6): 1013-1022.
[11] WANG L B, BALDWIN E A, BAI J H. Recent advance in aromatic volatile research in tomato fruit: the metabolisms and regulations[J]. Food and Bioprocess Technology, 2016, 9(2): 203-216. DOI:10.1007/ s11947-015-1638-1.
[12] BALDWIN E A, NISPEROSCARRIEDO M O, MOSHONAS M G. Quantitative analysis of flavor and other volatiles and for certain constituents of two tomato cultivars during ripening[J]. Journal of the American Society for Horticultural Science, 1990, 116(2): 265-269.
[13] KLEE H J, GIOVANNONI J J. Genetics and control of tomato fruit ripening and quality attributes[J]. Annual Review of Genetics, 2011, 45(1): 41-59. DOI:10.1146/annurev-genet-110410-132507.
[14] 罗云波. 脂氧合酶与番茄采后成熟的关系[J]. 园艺学报, 1994, 21(4): 357-360. DOI:10.3321/j.issn:0513-353X.1994.04.015.
[15] RAMBLA J L, TIKUNOV Y M, MONFORTE A J, et al. The expanded tomato fruit volatile landscape[J]. Journal of Experimental Botany, 2014, 65(16): 4613-4623. DOI:10.1093/jxb/eru128.
[16] SHEN J, TIEMAN D, JONES J B, et al. A 13-lipoxygenase, TomloxC, is essential for synthesis of C5 fi avour volatiles in tomato[J]. Journal of Experimental Botany, 2014, 65(2): 419-428. DOI:10.1093/jxb/ert382.
[17] ILG A, BRUNO M, BEYER P, et al. Tomato carotenoid cleavage dioxygenases 1A and 1B: relaxed double bond specificity leads to a plenitude of dialdehydes, mono-apocarotenoids and isoprenoid volatiles[J]. FEBS Open Bio, 2014, 4(1): 584-593. DOI:10.1016/ j.fob.2014.06.005.
[18] TIEMAN D, TAYLOR M, SCHAUER N, et al. Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde[J]. Proceedings of the National Academy of Sciences, 2006, 103(21): 8287-8292. DOI:10.1073/pnas.0602469103.
[19] TIEMAN D M, LOUCAS H M, KIM J Y, et al. Tomato phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma volatile 2-phenylethanol[J]. Phytochemistry, 2007, 68(21): 2660-2669. DOI:10.1016/j.phytochem.2007.06.005.
[20] TIEMAN D, ZEIGLER M, SCHMELZ E, et al. Functional analysis of a tomato salicylic acid methyl transferase and its role in synthesis of the fi avor volatile methyl salicylate[J]. Plant Journal, 2010, 62(1): 113-123. DOI:10.1111/j.1365-313X.2010.04128.x.
[21] MATHIEU S, DAL CIN V, FEI Z, et al. Flavour compounds in tomato fruits: identifi cation of loci and potential pathways affecting volatile composition[J]. Journal of Experimental Botany, 2009, 60(1): 325-337. DOI:10.1093/jxb/ern294.
[22] WANG L, BALDWIN E A, PLOTTO A, et al. Effect of methyl salicylate and methyl jasmonate pre-treatment on the volatile profi le in tomato fruit subjected to chilling temperature[J]. Postharvest Biology and Technology, 2015, 108: 28-38. DOI:10.1016/ j.postharvbio.2015.05.005.
[23] GRIFFITHS A, PRESTAGE S, LINFORTH R, et al. Fruit-specific lipoxygenase suppression in antisense-transgenic tomatoes[J]. Postharvest Biology and Technology, 1999, 17(3): 163-173. DOI:10.1016/s0925-5214(99)00051-4.
[24] CHEN A R S, CHASE T J R. Alcohol dehydrogenase 2 and pyruvate decarboxylase induction in ripening and hypoxic tomato fruit[J]. Plant Physiology and Biochemistry, 1993, 31(6): 875-885.
[25] RILEY J C M, WILLEMOT C, THOMPSON J E. Lipoxygenase and hydroperoxide lyase activities in ripening tomato fruit[J]. Postharvest Biology and Technology, 1996, 7(1/2): 97-107. DOI:10.1016/0925-5214(95)00032-1.
[26] TIEMAN D, BLISS P, MCINTYRE L, et al. The chemical interactions underlying tomato flavor preferences[J]. Current Biology, 2012, 22(11): 1035-1039. DOI:10.1016/j.cub.2012.04.016.
[27] LEWINSOHN E, SITRIT Y, BAR E, et al. Carotenoid pigmentation affects the volatile composition of tomato and watermelon fruits, as revealed by comparative genetic analyses[J]. Journal of Agricultural and Food Chemistry, 2005, 53(8): 3142-3148. DOI:10.1021/jf047927t.
[28] SIMKIN A J, SCHWARTZ S H, AULDRIDGE M, et al. The tomato carotenoid cleavage dioxygenase 1, genes contribute to the formation of the fi avor volatiles β- ionone, pseudoionone, and geranylacetone[J]. Plant Journal, 2004, 40(6): 882-892. DOI:10.1111/j.1365-313x.2004.02263.x.
[29] MAGEROY M H, TIEMAN D M, FLOYSTAD A, et al. A Solanum lycopersicum, catechol-O-methyltransferase involved in synthesis of the fi avor molecule guaiacol[J]. Plant Journal for Cell and Molecular Biology, 2012, 69(6): 1043-1051. DOI:10.1111/j.1365-313x.2011.04854.x.
[30] ACREE T, ARN H. Flavornet and human odor space. Gas chromatography-olfactometry (GCO) of natural products[EB/OL]. [2015-02-14]. http://www.fi avornet.org/fi avornet.html.
[31] MALONEY G S, KOCHEVENKO A, TIEMAN D M, et al. Characterization of the branched-chain amino acid aminotransferase enzyme family in tomato[J]. Plant Physiology, 2010, 153(3): 925-936. DOI:10.1104/pp.110.154922.
[32] 常培培, 梁燕, 张静, 等. 5 种不同果色樱桃番茄品种果实挥发性物质及品质特性分析[J]. 食品科学, 2014, 35(22): 215-221. DOI:10.7506/spkx1002-6630-201422042.
[33] BALDWIN E A, SCOTT J W, BAI J H. Sensory and chemical fi avor analyses of tomato genotypes grown in Florida during three different growing seasons in multiple years[J]. Journal of the American Society for Horticultural Science, 2015, 140(5): 490-503.
[34] 穆师洋, 胡文忠, 姜爱丽, 等. 乙烯的信号分子作用及其在采后果蔬生理代谢调控的研究进展[J]. 食品工业科技, 2015, 36(3): 375-378. DOI:10.13386 /j.issn1002-0306.2015.03.073.
[35] SPEIRS J, LEE E, HOLT K, et al. Genetic manipulation of alcohol dehydrogenase levels in ripening tomato fruit affects the balance of some fi avor aldehydes and alcohols[J]. Plant Physiology, 1998, 117(3): 1047-1058. DOI:10.1104/pp.117.3.1047.
[36] DALAL K B, OLSON L E, YU M H, et al. Gas chromatography of the fi eld-, glass-greenhouse-grown, and artifi cially ripened tomatoes: Lycopersicon esculentum mill.[J]. Phytochemistry, 1967, 6(1): 155-157. DOI:10.1016/0031-9422(67)85025-8.
[37] CEBOLLA-CORNEJO J, ROSELLÓ S, VALCÁRCEL M, et al. Evaluation of genotype and environment effects on taste and aroma flavor components of Spanish fresh tomato varieties[J]. Journal of Agricultural and Food Chemistry, 2011, 59(6): 2440-2450. DOI:10.1021/JF1045427.
[38] REYMOND P, FARMER E E. Jasmonate and salicylate as global signals for defense gene expression[J]. Current Opinion in Plant Biology, 1998, 1(1): 404-411. DOI:10.1016/S1369-5266(98)80264-1.
[39] 杨明惠, 陈海丽, 唐晓伟, 等. 不同栽培季节番茄果实芳香物质的比较[J]. 中国蔬菜, 2009(18): 8-13.
[40] OZORES-HAMPTON M, SIMONNE E H, MCAVOY E, et al. Nitrogen best management practice with tomato production in Florida in the 2005-2006 season[J]. Proceedings of the Florida State Horticultural Society, 119: 284-288.
[41] WRIGHT D H, HARRIS N D. Effect of nitrogen and potassium fertilization on tomato flavor[J]. Journal of Agricultural and Food Chemistry, 1985, 33(3): 355-358. DOI:10.1021/JF00063A009.
[42] 李梅兰, 吴俊华, 李远新, 等. 不同供硼水平对番茄产量及风味品质的影响[J]. 核农学报, 2009, 23(5): 875-878.
[43] 张志明. 二氧化碳施肥对番茄果实品质的影响[D]. 杭州: 浙江大学, 2012: 19-35.
[44] 侯雷平, 吴俊华, 李梅兰, 等. 供锌水平对番茄果实抗氧化性及风味品质的影响[J]. 植物营养与肥料学报, 2010, 16(3): 763-767. DOI:10.11674/zwyf.2010.0336.
[45] 吴俊华, 侯雷平, 李远新, 等. 不同供铁水平对番茄产量及果实风味品质的影响[J]. 土壤通报, 2011, 42(1): 154-157.
[46] 侯雷平, 吴俊华, 李远新. 缺锰和多锰对番茄产量及风味品质的影响[J]. 核农学报, 2010, 24(1): 83-87.
[47] KADER A A. Effects of postharvest handling procedures on tomato quality[J]. Acta Horticulturae, 1986, 190: 209-221. DOI:10.17660/ actahortic.1986.190.21.
[48] BALDWIN E, PLOTTO A, NARCISO J, et al. Effect of 1-methylcyclopropene on tomato flavour components, shelf life and decay as influenced by harvest maturity and storage temperature[J]. Journal of the Science of Food and Agriculture, 2011, 91(6): 969-980. DOI:10.1002/jsfa.4281.
[49] KADER A A, STEVENS M A, ALBRIGHT-HOLTON M, et al. Effect of fruit ripeness when picked on flavor and composition in fresh market tomatoes[J]. Journal of the American Society for Horticultural Science, 1977, 102(6): 724-731.
[50] MAUL F, SARGENT S A, BALABAN M O, et al. Aroma volatile profi les from ripe tomatoes are infi uenced by physiological maturity at harvest: an application for electronic nose technology[J]. Journal of the American Society for Horticultural Science, 1998, 123(6): 1094-1101.
[51] 解静. 1-MCP对番茄冷害及机械损伤的影响[D]. 杭州: 浙江大学, 2010: 10.
[52] SUSLOW T, CANTWELL M. Tomato: recommendations for maintaining postharvest quality[EB/OL]. [2015-02-14]. http:// postharvest.ucdavis.edu/pfvegetable/Tomato/.
[53] MAUL F, SARGENT S A, SIMS C A, et al. Tomato fi avor and aroma quality as affected by storage temperature[J]. Journal of Food Science, 2000, 65(7): 1228-1237. DOI:10.1111/j.1365-2621.2000.tb10270.x.
[54] WANG L, BALDWIN E A, ZHAO W, et al. Suppression of volatile production in tomato fruit exposed to chilling temperature and alleviation of chilling injury by a pre-chilling heat treatment[J]. LWT-Food Science and Technology, 2015, 62(1): 115-121. DOI:10.1016/ j.lwt.2014.12.062.
[55] WANG L, BALDWIN E A, YU Z F, et al. The impact of kitchen and food service preparation practices on the volatile aroma profi le in ripe tomatoes: effects of refrigeration and blanching[J]. HortScience, 2015, 50(9): 1358-1364.
[56] 金鹏, 王静, 朱虹, 等. 果蔬采后冷害控制技术及机制研究进展[J]. 南京农业大学学报, 2012, 35(5): 167-174. DOI:10.7685/ j.issn.1000-2030.2012.05.018.
[57] BAI Jinhe, BALDWIN E A, IMAHORI Y, et al. Chilling and heating may regulate C6 volatile aroma production by different mechanisms in tomato (Solanum lycopersicum) fruit[J]. Postharvest Biology and Technology, 2011, 60(2): 111-120. DOI:10.1016/ j.postharvbio.2010.12.002.
[58] BOUKOBZA F, TAYLOR A J. Effect of postharvest treatment on fi avour volatiles of tomatoes[J]. Postharvest Biology and Technology, 2002, 25(3): 321-331. DOI:10.1016/s0925-5214(02)00037-6.
[59] RATANACHINAKORN B, KLIEBER A, SIMONS D H. Effect of short-term controlled atmospheres and maturity on ripening and eating quality of tomatoes[J]. Postharvest Biology and Technology, 1997, 11(3): 149-154. DOI:10.1016/s0925-5214(97)00021-5.
[60] DELTSIDIS A I, PLIAKONI E D, BALDWIN E A, et al. Tomato fi avor changes at chilling and non-chilling temperatures as infi uenced by controlled atmospheres[J]. Acta Horticulturae, 2015, 1071: 703-710. DOI:10.17660/ActaHortic.2015.1071.93.
[61] 乔汝香. 植物生长调节剂在热带经济作物上的应用[J]. 中国农业信息, 2015(1): 132-133.
[62] 王建军, 周雅涵, 曾凯芳. 乙烯催熟对采后果实贮藏品质的影响[J]. 食品工业科技, 2012, 33(21): 361-364. DOI:10.13386/ j.issn1002-0306.2012.21.061.
[63] MCDONALD R E, MCCOLLUM T G, BALDWIN E A. Prestorage heat treatments infi uence free sterols and fi avor volatiles of tomatoes stored at chilling temperature[J]. Journal of the American Society for Horticultural Science, 1996, 121(3): 531-536.
[64] 王倩倩. 不同处理方法对寒露蜜桃和冬雪蜜桃贮藏效果的影响[D].青岛: 青岛农业大学, 2010: 10.
[65] WASTERNACK C, HAUSE B. Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. an update to the 2007 review in Annals of Botany.[J]. Annals of Botany, 2013, 111(6): 1021-1058. DOI:10.1093/aob/mct067.
[66] KONDO S, JITRATHAM A. Relationships between jasmonates and chilling injury in mangosteens are affected by spermine[J]. HortScience, 2004, 39(6): 1346-1348.
[67] YOSHIKAWA H, HONDA C, KONDO S. Effect of low-temperature stress on abscisic acid, jasmonates, and polyamines in apples[J]. Plant Growth Regulation, 2007, 52(3): 199-206. DOI:10.1007/s10725-007-9190-2.
[68] RUGKONG A, MCQUINN R, GIOVANNONI J J, et al. Expression of ripening-related genes in cold-stored tomato fruit[J]. Postharvest Biology and Technology, 2011, 61(1): 1-14. DOI:10.1016/ j.postharvbio.2011.02.009.
[69] KOCHEVENKO A, ARAÚJO W L, MALONEY G S, et al. Catabolism of branched chain amino acids supports respiration but not volatile synthesis in tomato fruits[J]. Molecular Plant, 2012, 5(2): 366-375. DOI:10.1093/mp/ssr108.
[70] FENG L, MENG W, JIA W, et al. Isolation of 2-phenylethanol biosynthesis related genes and their relationship with 2-phenylethanol accumulation in Rosa rugosa[J]. Acta Physiologiae Plantarum, 2015, 37(12): 1-9. DOI:10.1007/s11738-015-1996-3.
[71] WANG L B, BAI J, YU Z F. Difference in volatile profile between pericarp tissue and locular gel in tomato fruit[J]. Journal of Integrative Agriculture, 2016, 15(12): 2911-2920. DOI:10.1016/S2095-3119(15)61324-7.
[72] BOATRIGHT J, NEGRE F, CHEN X, et al. Understanding in vivo benzenoid metabolism in petunia petal tissue[J]. Plant Physiology, 2004, 135(4): 1993-2011. DOI:10.1104/pp.104.045468.
Recent Advances in Research on Volatile Aroma Compounds in Tomatoes and Their Impacting Factors
WANG Libin1,2,3, LI Xuehui4, SHI Zhenyuan5, BAI Jinhe2, JIN Changhai1, LUO Haibo6,*, YU Zhifang3,*
(1. College of Food Science and Engineering, Yangzhou University, Yangzhou 225127, China; 2. U.S. Horticultural Research Laboratory, U.S. Department of Agriculture, Fort Pierce 34945, USA; 3. College of Food Science and Technology, Nanjing Agricultural University, Nanjing 210095, China; 4. College of Life Science and Technology, Nanyang Normal University, Nanyang
473061, China; 5. Key Laboratory of Plant Functional Genomics, Ministry of Education, Yangzhou University, Yangzhou 225009, China; 6. Faculty of Food Science, Zhejiang Pharmaceutical College, Ningbo 315100, China)
Aroma is an important sensory attribute of tomatoes. Tomato aroma is formed by a complex mixture of more than 400 volatile compounds, and it plays an important role in the classifi cation and consumer acceptability of tomato products. This article provides a brief overview of the volatile aroma compounds of tomato fruits and their variations during ripening and summarizes their biosynthetic pathways. The focus of this review is on the factors affecting the formation of aroma compounds in tomato fruits. We expect this review to provide some theoretical and practical guidelines for researchers, producers, retailers and consumers.
tomato aroma; volatile compound; variation; biosynthetic pathway; impacting factors
10.7506/spkx1002-6630-201717047
TS255.1
A
1002-6630(2017)17-0291-10引文格式:
2016-08-24
公益性行业(农业)科研专项(2014030232);国家自然科学基金青年科学基金项目(31401612);浙江省自然科学基金面上项目(LY14C200005);南阳发展战略研究院项目(NYZL201629);南阳师范学院校级青年项目(QN2016004)
王利斌(1985—),男,讲师,博士,研究方向为新鲜农产品采后生物学。E-mail:wanglibin2013@sina.cn
*通信作者:罗海波(1979—),男,副教授,博士,研究方向为新鲜农产品采后生物学与处理技术。E-mail:luohaibo_1216@126.com
郁志芳(1960—),男,教授,博士,研究方向为新鲜农产品采后生物学。E-mail:yuzhifang@njau.edu.cn
猜你喜欢
分子催化(2022年1期)2022-11-02
化学工业与工程(2022年1期)2022-03-29
化工管理(2020年26期)2020-10-09
少儿美术(2019年8期)2019-12-14
少儿美术(快乐历史地理)(2018年7期)2018-04-02
岭南音乐(2017年3期)2017-07-18
中国塑料(2016年2期)2016-06-15
传奇故事(破茧成蝶)(2015年1期)2015-02-28
山西大同大学学报(自然科学版)(2015年1期)2015-01-22
应用化工(2014年11期)2014-08-16
