
家榆胚胎学研究
2017-11-10 01:28郑婧媛李芸增周萃萃卢耘申王秀华
植物研究 2017年5期
郑婧媛 李芸增 周萃萃 卢耘申 王秀华
(东北林业大学生命科学学院,哈尔滨 150040)
家榆胚胎学研究
郑婧媛 李芸增 周萃萃 卢耘申 王秀华*
(东北林业大学生命科学学院,哈尔滨 150040)
采用常规石蜡切片法,借助光学显微镜对家榆胚胎发育进行研究,研究结果表明:(1)家榆的花药具4个药室;小孢子母细胞减数分裂过程中的胞质分裂为同时型;小孢子四分体排列为四面体型和左右对称型;成熟花粉为2-细胞型;(2)花药壁发育类型为双子叶型,腺质绒毡层;(3)家榆子房两心皮1室,侧膜胎座,1枚倒生胚珠,双珠被,厚珠心;四分体呈直线型和T型排列;(4)家榆胚囊类型为单孢子蓼型胚囊;胚的发育类型为茄型;核型胚乳;无胚乳型种子。
家榆;胚胎学;大、小孢子;雌、雄配子体;胚;胚乳
家榆(UlmuspumilaL.)隶属榆科(Ulmaceae)榆属(UlmusL.)植物。别名榆、白榆、榆树。分布于东北、华北、西北及西南各省区。生于海拔1 000~2 500 m以下的山坡、山谷、川地、丘陵及沙岗等处。长江下游各省有栽培,也为华北及淮北平原农村的习见树木。朝鲜、前苏联、蒙古也有分布[1]。家榆具有较高的经济价值,以果实(榆钱)、树皮、叶、根入药,具有安神健脾的功效,用于神经衰弱,失眠,食欲不振,外伤出血,同时又是良好的防护林,四旁绿化和用材树种。
近年来,有关家榆的研究工作,在生理学,遗传学,生态学等多方面均有所进展[2~6],但有关家榆的胚胎学研究未见报道。本文采用石蜡切片技术对家榆的胚胎发育进行研究,以积累有关生殖生物学的基础资料,填补家榆胚胎学研究方面的空白,为家榆的育种与栽培以及家榆在榆属中的系统演化提供理论依据。
1 材料与方法
实验材料为东北林业大学林场内家榆,采样时间为2015年3月~2016年5月中旬。取材后用改良FAA对不同时期的材料进行固定保存,爱氏苏木精整体染色,采用常规石蜡切片法制片,切片厚度为8 μm[7],OLYMPUS-DP72显微照相系统进行观察拍摄。
2 结果与分析
2.1 小孢子的发生
家榆花两性,簇状聚伞花序[8],每朵小花有4枚雄蕊。由家榆雄蕊原基最初形成的幼小花药横切面来看,在花粉囊发育初期,由于花药的四个角隅处细胞分裂较快,使花药分裂成四个药室(图版Ⅰ:1),每个药室内分化出孢原细胞(图版Ⅰ:2)。孢原细胞体积较其他细胞大。进行平周分裂后,一个孢原细胞分裂成两个细胞,向外分裂成初生壁细胞,向内分裂成初生造孢细胞(图版Ⅰ:3)。初生造孢细胞进一步发育为排列紧密的次生造孢细胞(图版Ⅰ:4),次生造孢细胞继续分裂,形成多个小孢子母细胞。小孢子母细胞的形态与周围的药壁细胞具有显著的不同,体积大,细胞核较大,细胞质浓厚并且没有明显的液泡,具有不明显的较薄的纤维素壁,细胞排列紧密。小孢子母细胞发育到一定程度时进行减数分裂。(图版Ⅰ:5~12)为小孢子母细胞减数分裂时期的形态。随着减数分裂的结束,胼胝质的积累,细胞壁愈发明显,小孢子逐渐变成圆形。减数分裂后形成的四分体具有正四面体和左右对称两种结构(图版Ⅰ:13),孢质分裂时未出现分隔壁,孢质分裂为同时型。
2.2 雄配子体的发育
随着小孢子的发育,胼胝体逐渐溶解,小孢子从四分体中分离出来。刚释放出来的单核小孢子椭圆形,核清晰可见,位于细胞中央,细胞质浓厚,具小液泡,细胞壁比较薄,此时称为单核居中期小孢子(图版Ⅰ:14)。随后小孢子细胞局部会存在一定的收缩(图版Ⅰ:15),细胞壁开始加厚,细胞体积开始增大,并形成大液泡,细胞核被挤压到细胞的一边,称为小孢子单核靠边期(图版Ⅰ:16)。小孢子的核在靠近细胞壁的位置上开始进行有丝分裂,形成两个核,在两个核之间出现弧形的细胞板将两个核隔离开,大的为营养细胞,小的为生殖细胞并紧贴细胞壁(图版Ⅰ:17~18)。随后弧形细胞板消失,生殖细胞逐渐向中央移动,并且变大成球形,成为一个裸细胞游离在细胞质中。生殖细胞的细胞核大,占据细胞体积的大部分,细胞质稀少(图版Ⅰ:19)。在发育过程中,营养细胞的内容物大量积累,使花粉粒的体积迅速增大,比二细胞花粉初期增大许多。至传粉前家榆花粉粒的生殖细胞一直不分裂,即家榆的成熟花粉粒为2-细胞型。
2.3 花药壁和绒毡层
家榆的花药壁类型为双子叶型,由初生壁细胞经过平周分裂和垂周分裂形成。初生壁细胞经过一次平周分裂和多次垂周分裂后产生了两层次生壁细胞(图版Ⅰ:3)。在初生造孢细胞分裂形成次生造孢细胞的过程中,花药壁发育成四层细胞结构,从外到内依次是表皮,药室内壁,中层,绒毡层,此时绒毡层细胞尚未分化完全(图版Ⅰ:4)。花药壁结构发育完成时,药室内正处于小孢子母细胞时期,此时的绒毡层多为单核(图版Ⅰ:5)。小孢子母细胞减数分裂过程中绒毡层发生核分裂,体积增大,形成多核绒毡层(图版Ⅰ:6~7)。在小孢子母细胞减数第一次分裂结束时可观察到绒毡层的原位溶解现象,为小孢子母细胞的分裂提供营养物质,在释放单核小孢子时期仅能看到部分绒毡层溶解后留下的残迹(图版Ⅰ:14),故家榆的绒毡层为腺质绒毡层。在2-细胞花粉晚期已看不到中层,仅由一层残留的表皮和纤维加厚的药室内壁包围着花粉粒(图版Ⅰ:20)。
2.4 大孢子的发生
家榆的子房两心皮1室,侧膜胎座,一枚倒生胚珠,双珠被,厚珠心。大孢子的形成开始于珠心表皮下的一个孢原细胞。孢原细胞进行平周分裂,产生一个周缘细胞和一个造孢细胞(图版Ⅱ:1)。珠心表皮和周缘细胞共同分裂产生周缘珠心细胞。随后,造孢细胞的体积增加、细胞核扩大、细胞质变浓,染色质凝聚于核仁周围,直接发育成大孢子母细胞(图版Ⅱ:2)。大孢子母细胞的体积较周围的珠心细胞大,细胞质较浓,核与核仁位于细胞中央,体积较大。大孢子母细胞形成不久后开始进行减数分裂,逐渐变为椭圆型。大孢子第一次减数分裂(图版Ⅱ:3~5)后形成了大孢子二分体(图版Ⅱ:6)。然后进行减数第二次分裂,形成直线型的四分体(图版Ⅱ:7),并出现少许的T型四分体(图版Ⅱ:8)。四分体形成后,只有合点端的大孢子起作用,称为功能大孢子(图版Ⅱ:9),其余三个退化。功能大孢子形成后,细胞体积迅速增大,是雌配子体的第一个细胞。
2.5 雌配子体的发育
随着功能大孢子体积的增大,发育为单核胚囊(图版Ⅱ:10)。单核胚囊经过一次核分裂,发育成二核胚囊(图版Ⅱ:11)。二核胚囊又在珠孔端和合点端分别进行有丝分裂,在胚囊的两端分别形成一对姐妹核,即四核胚囊(图版Ⅱ:12)。由四核胚囊的每个核各自进行第三次有丝分裂,形成八核胚囊(图版Ⅱ:13~15)。八核胚囊经过短暂的停留期便开始细胞化,形成的成熟胚囊包括珠孔端的一个洋梨形卵细胞(图版Ⅱ:16),两个楔形助细胞(图版Ⅱ:17),中央的两个开始融合的极核(图版Ⅱ:18),合点端的三个品字形反足细胞(图版Ⅱ:19~20),其中卵细胞和助细胞组成卵器。由此可见,家榆的胚囊类型为为蓼型。
2.6 胚和胚乳的发育
2.6.1 胚的发育
卵细胞和精细胞结合后形成的合子(图版Ⅲ:1)是胚胎发育的开始。合子核大质浓,核仁明显,经过一次有丝分裂后产生两个细胞,即二细胞原胚(图版Ⅲ:2),靠近珠孔端的为基细胞,靠近合点端的为顶细胞。基细胞、顶细胞分裂,形成直线排列多细胞原胚(图版Ⅲ:3)。进而发育成早期球形胚(图版Ⅲ:4),后发育成球形胚(图版Ⅲ:5)。球形胚后期,顶部两侧细胞分裂较快,胚胎进一步发育成心形胚(图版Ⅲ:5)。可观察到球形胚和心形胚时期胚柄结构发达,直到发育到鱼雷胚的晚期(图版Ⅲ:7),胚柄才退化消失,此时鱼雷胚的子叶明显增长,发育成子叶型胚(图版Ⅲ:13)。因此,家榆胚胎发育类型为茄型。
2.6.2 胚乳的发育
中央细胞受精后形成初生胚乳核(图版Ⅲ:8)。家榆的胚乳发育早于胚胎发育,在合子休眠时期,胚乳开始快速进行有丝分裂,发育为许多游离的胚乳核,并且产生中央大液泡(图版Ⅲ:9)。可见,家榆的胚乳发育类型为核型。胚乳游离核的增殖时间较长,不断积累可达数百个(图版Ⅲ:10)。在球形胚晚期,胚乳开始细胞化(图版Ⅲ:11)。细胞化的胚乳继续发育,在细胞内积累淀粉等营养物质(图版Ⅲ:12)。当胚胎发育到子叶胚时期,未观察到胚乳(图版Ⅲ:13),故家榆的种子为无胚乳型种子。
3 结论
从胚胎学角度分析,家榆具有以下特性:花药具有四个药室,小孢子母细胞减数分裂为同时型,小孢子四分体为正四面体型和左右对称型,成熟花粉为2-细胞型。花药壁发育为双子叶型,腺质绒毡层。子房一室,单枚倒生胚珠,双珠被,厚珠心,具有直线型和T型大孢子四分体,胚囊的发育类型为单孢子发生型,是典型的蓼型胚囊。胚的发育类型为茄型,先后经历了原胚期、早期球形胚、球形胚、心形胚、鱼雷胚和子叶胚。核型胚乳,无胚乳型种子。
从本研究结果可见:家榆的小孢子发生与已报道的其他榆科的物种发育特性不尽相同。均是孢质分裂同时型,腺质绒毡层;不同的是朴树的花药壁为基本型,家榆的花药壁为双子叶型[9~10]。朴属(CeltisL.),青檀属(PteroceltisMaxim.)均为2-细胞花粉,榆属(UlmusL.)为3-细胞花粉[9~10],但家榆却是2-细胞花粉。从已研究过榆科其他属的植物[9~10]和Walker等对榆属植物的胚胎研究来看[11~14],青檀属(PteroceltisMaxim.)的胚囊类型为蓼型,榆属(UlmusL.)和朴属(CeltisL.)的胚囊类型多为德鲁撒型或五福花型[10],但根据本文的研究,家榆具有蓼型胚囊。榆科植物胚的发育只在榆属(Ulmus),Homoptelea(我国无此属)和朴属(CeltisL.)中研究过,榆属(UlmusL.)和朴属(CeltisL.)为茄型,Homoptelea为十字花型,胚乳发育均为核型胚乳[15~16],家榆的胚和胚乳则符合上述情况。朴树(CeltissinensisPers.)的胚乳在胚发育过程中逐渐消失,家榆种子亦是无胚乳型种子。
榆属是由林奈于1754年创立的。根据不同的性状,如开花季节、花序演化及翅果特征等的不同,分类学家对榆属植物的系统演化位置,其属下等级的划分均存在不少争论[1,17~18]。榆属植物开花可分春季开花和秋季开花两类,Dippel以开花季节和花序着生的部位可分为两个亚属(Subgen.1.EuulmusK.Kock和Subgen.1.Microptelea(Spacb)Planchon)。秋季开花的榆树,其花序均生于当年生枝的叶腋(如榔榆UlmusparvifoliaJacq.),春季开花的榆树则按其花序着生的部位分为四类,其中包括花全部自花芽抽出而生于去年生枝的叶痕上部、花开于新生枝叶发出之前的家榆。就系统位置而言,Schneider认为秋季开花的在前,而Rehder、傅立国等认为春季开花的在前[17,19]。其中傅立国就榆属植物的开花季节,花序的演化及其相关性状作为划分和排列系统位置的依据,将榆属的系统位置重新排列分类为组1:长序榆组;组2:秋榆组;组3:睫毛榆组;组4:榆组;组5:榔榆组[18]。
本研究得出榆树的花粉类型是较原始的2-细胞花粉;具有双珠被、厚珠心,蓼型胚囊发育均属于比较原始的胚胎学发育特征,Hall认为倒生胚珠比弯生胚珠及直生胚珠原始[20],故家榆具有较原始的胚胎学发育特性。因此,本研究赞同傅立国在榆属之下的分组的演化关系,即春季开花的家榆在系统位置上较秋季开花的榔榆(UlmusparvifoliaJacq.)相对靠前。
1.黄成就.中国植物志:第22卷[M].北京:科学出版社,1998:334-413.
Huang C J.Flora republicae popularis sinicae:vol.22[M].Beijing:Science Press,1998:334-413.
2.李方祯.锡盟沙地榆(Ulmuspumilavar.sabulosa)遗传多样性及其与家榆(U.pumila)的比较研究[D].呼和浩特:内蒙古农业大学,2008.
Li F Z.The genetic diversity study ofUlmuspumilavar.sabulosaand with notes on its relationship toU.pumilaL.[D].Hohhot:Inner Mongolia Agricultural University,2008.
3.王亚秋.榆树个体生态场行为的研究[D].长春:东北师范大学,2005:4.
Wang Y Q.The research on the ecological field behavior ofUlmuspumilaindividual[D].Changchun:Northeast Normal University,2005:4.
4.Wang Q.The programmed cell death nuclear characteristics and preliminary study on signal transduction pathway during control deterioration ofUlmuspumilaL.seeds[D].Beijing:Beijing Forestry University,2012:4.
5.李茜茜.家榆种子人工老化过程中细胞程序性死亡机理的研究[D].北京:北京林业大学,2009:5.
Li Q Q.Studies on the mechanism of PCD of elm seeds during accelerated aging[D].Beijing:Beijing Forestry University,2009:5.
6.曹书敏,杨晴,杨俊明,等.家榆和金叶榆光合·蒸腾及荧光参数对水分胁迫的响应[J].安徽农业科学,2011,39(22):13477-13480.
Cao S M,Yang Q,Yang J M,et al.Research on the response of the photosynthesis,transpiration and fluorescence index ofUlmuspumilaL.andUlmuspumilacv.jinyeto Water Stress[J].Journal of Anhui Agricultural Sciences,2011,39(22):13477-13480.
7.李和平.植物显微技术:2版[M].北京:科学出版社,2009.
Li H P.Microscopic technique of plant:2nd ed[M].Beijing:Science Press,2009.
8.中国科学院植物研究所.中国高等植物图鉴(补编第一册)[M].北京:科学出版社,1982.
Institute of botany.The Chinese academy of sciences.Iconographia cormophytorum sinicorum[M].Beijing:Science Press,1982.
9.管亦农.青檀的生殖过程[C].//中国植物学会.中国植物学会五十五周年年会论文摘要汇编.北京:高等教育出版社,1988:376-377.
Guan Y N.The reproductive process ofPteroceltistatarinowiiMaxim[C].//Botanical Society of China.Summary of papers for the fifty-fifth anniversary of botanical society of China.Beijing:Higher Education Press,1988:376-377.
10.任群,郑亦津,张昭洁.朴树大小孢子发生及雌雄配子体形成[J].山东大学学报:自然科学版,1991,26(4):467-475.
Ren Q,Zheng Y J,Zhang Z J.Studies on megasporogenesis microsporogenesis,megagametogenesis microgametogenesis ofCeltissinensisPers[J].Journal of Shandong University,1991,26(4):467-475.
11.Bhojwani S S ,Bhatnagar S P.The embryology of angiosperms[M].New Delhi Bombay Bangalore Calcutta Kanpur :Vikas Publishing House PVT Ltd.,1978:74,78.
12.Walker R I.Megasporogenesis and development of megagametophyte in Ulmus[J].American Journal of Botany,1950,37(1):47-52.
13.胡适宜.被子植物生殖生物学[M].北京:高等教育出版社,2005.
Hu S Y.Reproductive biology of angiosperms[M].Beijing:Higher Education Press,2005.
14.樊汝汶,乔士义,李文钿.树木显微解剖图谱[M].北京:中国林业出版社,1989:106-116.
Fan R W,Qiao T Y,Li W D.Microscopic anatomical atlas of trees[M].Beijing:China Forestry Publishing House,1989:106-116.
15.Davis G L.Systematic embryology of the Angiosperms[J].New York:Wiley,1966:266-267.
16.任群,张昭洁,陈永喆,等.朴树的受精作用及胚、胚乳的发育[J].山东大学学报:自然科学版,1993,28(1):98-105.
Ren Q,Zhang Z J,Chen Y Z,et al.Fertilization and development of embryo and endosperm inCeltissinensisPers[J].Journal of Shandong University,1993,28(1) :98-105.
17.Planchon M J E.Sur les ulmacées[J].New York:Ann Sci Nat Bot,1848,3(10):244-341.
18.傅立国.中国榆属的研究[J].东北林学院学报,1980(3):1-40.
Fu L G.Notulae de Ulmus sinensibus[J].Journal of NorthEast Forestry University,1980(3):1-40.
19.李法曾,张学杰.中国榆科植物系统分类研究综述[J].武汉植物学研究,2000,18(5):412-416.
Li F Z,Zhang X J.A summary on phytogenetic classification of Ulmaceae from China[J].Journal of Wuhan Botanical Research,2000,18(5):412-416.
20.Hall M A.Plant structure,function and adaptation[M].Yao B J,Trans.Beijing:Science Press,1987:100-120.
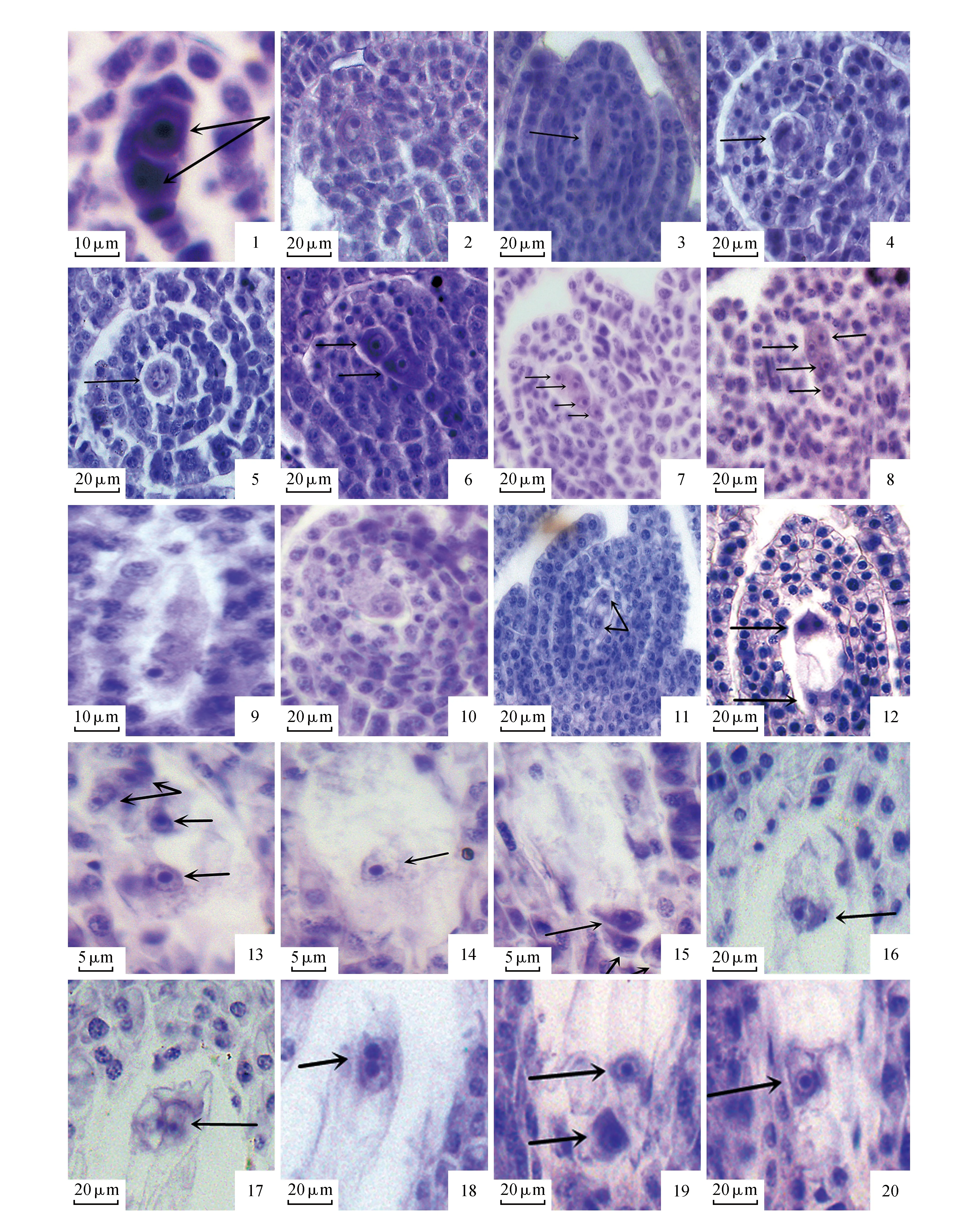
图版Ⅱ 1.孢原细胞分裂;2.大孢子母细胞;3~5.大孢子母细胞减数分裂;6.二分体;7.直线型四分体;8.T型四分体;9.功能大孢子;10.单核胚囊;11.二核胚囊;12.四核胚囊;13~15.八核胚囊;16.卵细胞;17.助细胞;18.融合的极核;19~20.品字形反足细胞Plate Ⅱ 1.Archesporial cell division; 2.Megaspore mother cell; 3-5.Meiosis of megaspore mother cell; 6.Dyad; 7.A linear tetrad; 8.T-shaped megasporocyte tetrad; 9.Functional megaspore; 10.Uninucleate embryo sac; 11.Two-nucleate embryo sac; 12.Four-nucleate embryo sac; 13-15.Eight- nucleate embryo sac; 16.Egg cell; 17.Two synergids of the mature embryo sac; 18.Polar fusion nucleus; 19-20.Three antipodal cells
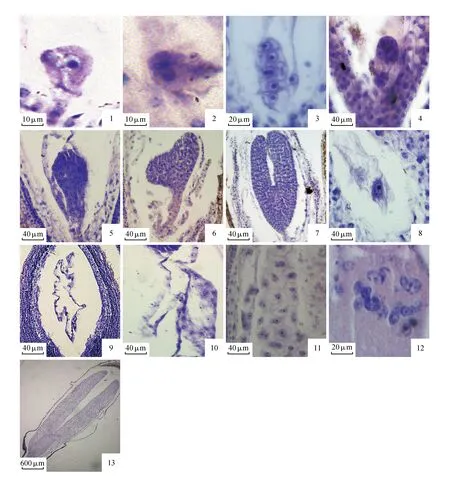
图版Ⅲ 1.合子;2.二细胞原胚;3.六细胞原胚;4.早期球型胚;5.球型胚;6.心形胚;7.鱼雷胚;8.极核受精;9.游离的胚乳核;10.胚乳核增殖期;11.胚乳细胞化;12.细胞化后的胚乳细胞积累营养物质;13.无胚乳型种子Plate Ⅲ 1.Zygote; 2. 3-elled proembryo; 3. 6-celled proembryo; 4.Early globular embryo; 5.Globular embryo; 6.Heart-shaped embryo; 7.Torpedo-shaped embryo; 8.Sperm nucleus is fusing with upper polar nucleus; 9.Free endosperm nucleus; 10.Endosperm increment period; 11.Endosperm cells are forming showing the free growing wall margin continues to grow; 12.Endosperm cells accumulate nutrients; 13.No endosperm seeds
This study was funded by the college student innovation project of the College of Life Science,Northeast Forestry University.
introduction:ZHENG Jing-Yuan(1991—),female,master degree,main research interests focus on Plant reproductive science.
date:2017-04-09
StudyonEmbryologyofUlmuspumilaL.
ZHENG Jing-Yuan LI Yun-Zeng ZHOU Cui-Cui LU Yun-Shen WANG Xiu-Hua*
(College of Life Science,Northeast Forestry University,Haerbin 150040)
The processes of embryology ofUlmuspumilaL. were observed by serial paraffin sections and under optical microscope. The results are as follows: the anther wall ofU.pumilaL. includes 4 layers; the cytoplasmic division of microspore mother cells during meiosis was simultaneous; the tetrad of microspores resulted from the meiosis are simultaneously bilateral and tetrahedral;mature pollen grains are two-cell type. The development of the anther wall belongs to a dicotyledonous type. Tapetum belongs to glandular tapetum. The ovary consists of 2 carpels with a single locule,parietal placenta,anatropous ovule,double integument,and thick nucellus. The tetrad of megaspores is linear or T-type in arrangement.Embryo sac ofU.pumilaL. is of single-spore polygonum type. The embryogenesis ofU.pumilaL. conformed to the So-lanad type; karyotype endosperm; cotyledon embryo without endosperm.
UlmuspumilaL.;embryology;megaspore and microspore;female and male gamete;embryo and endosperm
东北林业大学生命科学学院院级大学生创新项目资助
郑婧媛(1991—) ,女,硕士研究生,主要从事植物生殖生物学研究。
* 通信作者:E-mail:shengming615@163.com
2017-04-09
* Corresponding author:E-mail:shengming615@163.com
S564.1
A
10.7525/j.issn.1673-5102.2017.05.003
猜你喜欢
中国临床医学影像杂志(2022年6期)2022-07-26
西北植物学报(2022年4期)2022-05-24
科学大众·教师版(2022年6期)2022-05-23
北京航空航天大学学报(2021年9期)2021-11-02
辽宁农业科学(2021年1期)2021-03-17
国际放射医学核医学杂志(2021年10期)2021-02-28
中国临床医学影像杂志(2019年1期)2019-04-25
中国生殖健康(2019年9期)2019-01-07
小学阅读指南·高年级版(2016年9期)2016-10-31
中国粮油学报(2016年5期)2016-01-23
