
高温和干旱胁迫对杨树幼苗光合性能和抗氧化酶系统的影响1)
2017-11-28 07:40张燕红吴永波刘璇朱嘉馨
东北林业大学学报 2017年11期
张燕红 吴永波 刘璇 朱嘉馨
(江苏省南方现代林业协同创新中心(南京林业大学),南京,210037)
高温和干旱胁迫对杨树幼苗光合性能和抗氧化酶系统的影响1)
张燕红 吴永波 刘璇 朱嘉馨
(江苏省南方现代林业协同创新中心(南京林业大学),南京,210037)
采用室内模拟试验,研究了高温、干旱单一或复合胁迫对南林895杨(PopulusבNanlin-895’)幼苗光合特性、叶绿素荧光、抗氧化酶活性的影响。结果表明:随着胁迫时间的延长,高温、干旱单因子和复合胁迫使杨树幼苗净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、最大荧光(Fm)、PSII最大原初光化学量子效率(Fv/Fm)下降、初始荧光(Fo)升高,中度干旱胁迫使胞间CO2摩尔分数(Ci)、水分利用效率(WUE)先降后升,超氧化物歧化酶(SOD)、过氧化物酶(POD)活性先升后降,Pn主要受气孔和非气孔因素共同限制;重度高温、重度干旱和复合胁迫末期SOD、POD活性下降显著,Pn主要受非气孔因素限制。复合胁迫对Pn、Tr具有叠加效应。复合胁迫对Pn、Gs、Ci、Tr、Fv/Fm、SOD的胁迫效应大于单一胁迫;杨树幼苗在高温和干旱复合胁迫条件下受害最严重。
南林895杨;高温胁迫;干旱胁迫;光合特性;抗氧化酶;叶绿素荧光
植物的光合作用对环境变化非常敏感。在高温或干旱胁迫下植物体内会产生大量有害自由基,超氧化物歧化酶和过氧化物酶等抗氧化酶会通过提高自身活性来清除体内自由基以维持植物机体平衡[1]。当自由基积累过量时则表现出酶活性降低、光合速率下降的现象[2]。叶绿素荧光是光合作用研究的探针,在植物逆境胁迫方面得到了广泛地应用[3]。植物在受到逆境胁迫时,主要表现为PII反应中心活性下降。高温、干旱单因子胁迫已经成为目前的研究热点,主要集中在叶片形态结构、光合特性、水分利用效率等方面[4-5]。关于高温和干旱复合胁迫对植物的影响方面的研究较少,且大多集中于农作物籽粒和草本植物叶肉细胞形态结构等[6],对木本植物生理生化性能特别是光合性能、抗氧化酶系统影响的研究鲜有报道[7-8]。植物在复合胁迫下,各生化性能的变化机制非常复杂,并不能简单地通过单一胁迫的结果叠加或抵消获得[9]。笔者研究了高温与干旱复合胁迫对杨树幼苗叶片光合性能、抗氧化酶系统的影响,揭示杨树幼苗对高温与干旱复合胁迫的抗性机理和响应机制,为探讨高温和干旱气候条件下,杨树应对逆境胁迫的适应机理提供科学依据。
1 材料与方法
1.1 材料
控制试验在FYS-8智能人工气候室进行(3.5 m×2.5 m×3.0 m,高压钠灯光源,最大光合有效辐射为1 800 μmol·m-2·s-1,最大光照度为80 000 lx,空气相对湿度为60%),供试土壤为黄棕壤,pH=5.96,有机质质量分数15.14 g·kg-1,全氮质量分数0.83 g·kg-1,碱解氮质量分数15.79 mg·kg-1,全磷质量分数0.34 g·kg-1,有效磷质量分数1.34 mg·kg-1。杨树品种为南林895杨(PopulusבNanlin-895’),穗长20~23 cm,径粗2.0 cm。花盆内径30.0 cm,高20.7 cm。于2015年6月开始盆栽试验。
1.2 试验设计
将长势良好且基本一致的杨树幼苗放入3个不同温度的人工气候室内,夜间温度均为25 ℃,白天温度分别为25 ℃(适宜温度T0)、35 ℃(中度高温T1)、40 ℃(重度高温T2)。分别设置3个水分梯度,土壤含水率分别为田间持水量的70%(适宜水分D0)、55%(中度干旱D1)、40%(重度干旱D2)。即:T0D0(对照)、T0D1、T0D2、T1D0、T1D1、T1D2、T2D0、T2D1、T2D2共9个处理。每个处理设置4个重复,共36盆。每日06:00,采用称质量与TDR200土壤水分测定仪相结合的方法对每盆植物进行称质量补充水分,确保土壤含水率在控制处理下。同时模拟自然光照,调节光照量,于每日06:00打开光照、18:00关闭光照。胁迫试验维持7周,于第4周开始每周选取一天测定各项生理生化指标。
1.3 光合参数和荧光参数测定
于测定日10:00用Li-6400XT便携式光合仪连体测定。每个处理随机选取3株,在每株苗木中上部标记3片健康叶片作为待测叶片。测定指标包括净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)和胞间CO2摩尔分数(Ci,μmol·mol-1)。水分利用效率采用光合速率和蒸腾速率的比值计算。光合参数测定结束后,采用OS5-FL调制解调荧光仪(美国Opti-Science)测定。随机抽取3株幼苗,选择不同部位3片叶片,用叶夹暗适应且关闭气候室灯光,20 min后,测量初始荧光Fo、最大荧光Fm,以及PSII最大潜在光化学效率Fv/Fm。
1.4 抗氧化酶活性测定
取鲜叶0.2 g于预冷研钵中,加入2 mL预冷的50 mmol·L-1、pH=7.8的磷酸缓冲液在冰浴中研磨成匀浆,加入提取介质冲洗研钵2~3次,使终体积为10 mL。于4 ℃下10 000g离心15 min,上清液即为酶提取液,在冰浴中保存备用。酶提取液用于超氧化物歧化酶(SOD)、过氧化物酶(POD)的测定。SOD活性采用氮蓝四唑光还原法[10],POD活性采用愈创木酚显色法[11]。
1.5 数据处理
采用多重比较最小显著差法(LSD)和SNK法进行检验,用Microsoft office软件制图表。
2 结果与分析
2.1 不同处理对杨树幼苗光合性能指标的影响
2.1.1对叶片净光合速率(Pn)的影响
从表1可见,各处理下,Pn总体上呈下降趋势,除T1D0处理外,各时期均与对照差异显著(Plt;0.05)。高温或干旱单因子胁迫的各时期内,T1D0处理的Pn均大于T2D0处理,T0D1处理大于T0D2。单因子胁迫末期Pn在T1D0、T2D0、T0D1、T0D2处理下与对照相比分别下降了27.5%、52.5%、34.4%、50.2%,表现出重度高温胁迫对Pn的影响大于干旱胁迫。复合胁迫末期,Pn与单一胁迫相比下降显著,T1D1、T1D2、T2D1、T2D2处理与对照相比分别下降了52.1%、60%、67.3%、76.7%。高温和干旱双重逆境对Pn胁迫效应由大到小为T2D1、T2D2、T1D2、T2D0、T0D2、T1D1、T0D1、T1D0(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫极显著影响Pn(表3)。
2.1.2对叶片气孔导度(Gs)的影响
从表1可见,高温、干旱单因子胁迫下,Gs变化规律与Pn相同,T0D2、T2D0处理在各时期显著低于对照,并随着胁迫时间的延长差异增大。单因子胁迫下,Gs在各个时期内表现为T1D0处理大于T2D0,T0D1处理大于T0D2,T1D0、T2D0、T0D1、T0D2胁迫末期分别较对照下降28.5%、68.8%、37.2%、53.7%。由此可见,重度高温胁迫对Gs的影响大于干旱胁迫,单因子中度胁迫的影响低于重度胁迫。复合胁迫下,Gs呈先快速下降后缓慢上升的趋势,除第7周T2D1处理外,各时期均小于单一胁迫,T1D1、T2D1、T2D2处理最小值均出现在第6周,分别较对照降低41.2%、83.3%、70.7%。高温和干旱双重逆境对Gs胁迫效应由大到小依次为T2D1、T2D2、T1D2、T2D0、T1D1、T0D2、T0D1、T1D0(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫极显著影响Gs(表3)。
2.1.3对叶片胞间CO2摩尔分数(Ci)的影响
从表1可见,单因子胁迫T1D0、T0D1处理下Ci呈先下降后上升的趋势,分别在胁迫第5、第6周时达到最小值,T2D0、T0D2处理下Ci呈上升趋势,处理间差异显著。胁迫末期T1D0、T2D0、T0D1、T0D2分别较对照上升16.4%、31.6%、12.1%、37.5%。复合胁迫下,各处理Ci变化趋势不一致,T1D2、T2D1、T2D2处理下呈上升趋势,在整个胁迫过程中均显著高于对照,T1D1呈先降后升趋势,最小值出现在第5周。胁迫末期,各复合胁迫处理均高于单一胁迫,T1D1、T1D2、T2D1、T2D2处理下分别较对照上升28.4%、54%、69%、62.8%,表现出温度越高、干旱越严重,Ci上升幅度越大的趋势。高温和干旱双重逆境对Ci胁迫效应由大到小依次为T2D2、T2D1、T1D2、T2D0、T0D2、T1D1、T1D0、T0D1(表2)。双因素方差分析结果表明高温、干旱以及复合胁迫极显著影响Ci(表3)。

表1 高温和干旱胁迫对杨树幼苗光合性能指标的影响
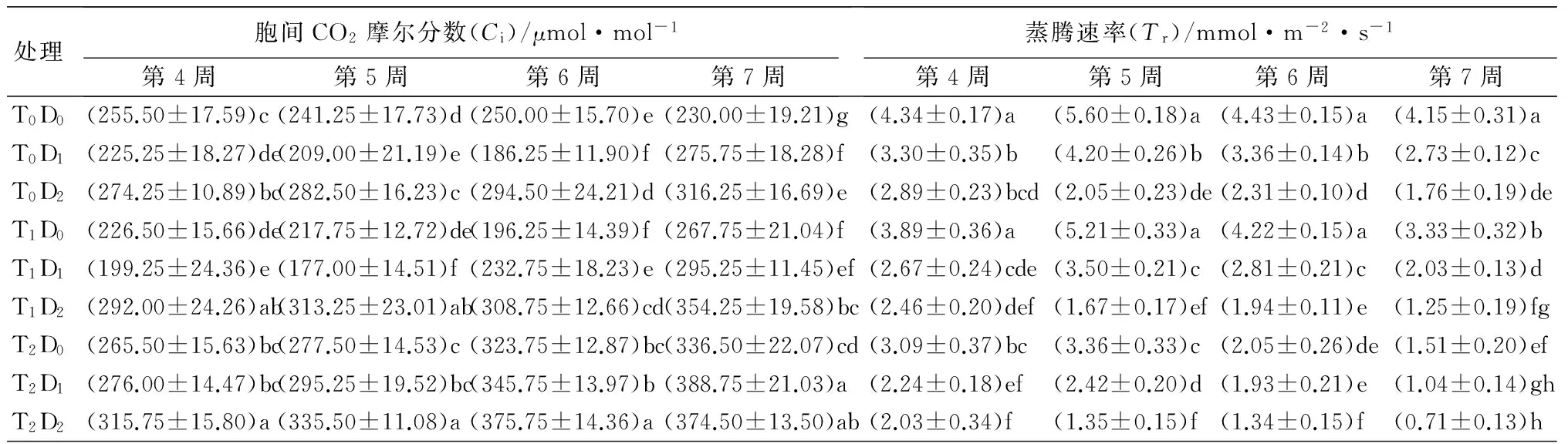
处理胞间CO2摩尔分数(Ci)/μmol·mol-1第4周第5周第6周第7周蒸腾速率(Tr)/mmol·m-2·s-1第4周第5周第6周第7周T0D0(255.50±17.59)c(241.25±17.73)d(250.00±15.70)e(230.00±19.21)g(4.34±0.17)a (5.60±0.18)a(4.43±0.15)a(4.15±0.31)aT0D1(225.25±18.27)de(209.00±21.19)e(186.25±11.90)f(275.75±18.28)f(3.30±0.35)b(4.20±0.26)b(3.36±0.14)b(2.73±0.12)cT0D2(274.25±10.89)bc(282.50±16.23)c(294.50±24.21)d(316.25±16.69)e(2.89±0.23)bcd(2.05±0.23)de(2.31±0.10)d(1.76±0.19)deT1D0(226.50±15.66)de(217.75±12.72)de(196.25±14.39)f(267.75±21.04)f(3.89±0.36)a(5.21±0.33)a(4.22±0.15)a(3.33±0.32)bT1D1(199.25±24.36)e(177.00±14.51)f(232.75±18.23)e(295.25±11.45)ef(2.67±0.24)cde(3.50±0.21)c(2.81±0.21)c(2.03±0.13)dT1D2(292.00±24.26)ab(313.25±23.01)ab(308.75±12.66)cd(354.25±19.58)bc(2.46±0.20)def(1.67±0.17)ef(1.94±0.11)e(1.25±0.19)fgT2D0(265.50±15.63)bc(277.50±14.53)c(323.75±12.87)bc(336.50±22.07)cd(3.09±0.37)bc(3.36±0.33)c(2.05±0.26)de(1.51±0.20)efT2D1(276.00±14.47)bc(295.25±19.52)bc(345.75±13.97)b(388.75±21.03)a(2.24±0.18)ef(2.42±0.20)d(1.93±0.21)e(1.04±0.14)ghT2D2(315.75±15.80)a(335.50±11.08)a(375.75±14.36)a(374.50±13.50)ab(2.03±0.34)f(1.35±0.15)f(1.34±0.15)f(0.71±0.13)h

处理水分利用效率(WUE)/μmol·mol-1第4周第5周第6周第7周T0D0(3.21±0.11)a(2.39±0.15)b(2.96±0.28)bc(3.12±0.51)aT0D1(3.34±0.83)a(2.24±0.27)b(2.92±0.24)bc(3.11±0.53)aT0D2(3.41±0.55)a(4.09±0.59)a(3.31±0.35)bc(3.68±0.74)aT1D0(3.33±0.53)a(2.31±0.24)b(2.74±0.26)c(2.84±0.50)aT1D1(3.64±0.57)a(2.29±0.12)b(3.04±0.67)bc(3.05±0.57)aT1D2(3.05±0.43)a(3.97±0.79)a(3.28±0.54)bc(4.22±1.15)aT2D0(2.80±0.44)a(2.25±0.34)b(3.68±0.74)ab(4.05±0.70)aT2D1(3.13±0.67)a(1.82±0.39)b(2.91±0.39)bc(4.00±0.97)aT2D2(3.68±0.60)a(4.75±0.80)a(4.37±0.58)a(4.32±1.34)a
注:表中数据为平均值±标准差。同列数据后不同小写字母表示处理间差异显著(Plt;0.05)。

表2 高温和干旱胁迫对杨树幼苗生理生化指标的胁迫效应
2.1.4对叶片蒸腾速率(Tr)的影响
从表1可见,高温胁迫T1D0、T2D0处理下Tr呈先上升后下降的趋势,从第7周开始T1D0处理与对照差异显著,各时期T2D0处理均显著低于对照,处理间差异显著,表现为T1D0大于T2D0。干旱胁迫T0D1处理Tr变化趋势与高温胁迫相同,T0D2总体上呈下降趋势,干旱胁迫各时期显著低于对照,表现为T0D1大于T0D2。T1D0、T2D0、T0D1、T0D2胁迫末期较对照分别下降39%、63.6%、34.3%和57.7%,说明胁迫对Tr有抑制作用。复合胁迫T1D1、T2D1处理下Tr呈先上升后下降的趋势,T1D2、T2D2处理下Tr总体上呈下降趋势,各时期Tr均低于单一胁迫处理,T1D1、T1D2、T2D1、T2D2在胁迫末期分别下降51.21%、69.80%、74.90%和83.00%。高温和干旱双重逆境对Tr胁迫效应由大到小依次为T2D2、T1D2、T2D1、T0D2、T2D0、T1D1、T0D1、T1D0(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫极显著影响Tr(表3)。
2.1.5 对叶片水分利用效率(WUE)的影响
从表1可见,高温和干旱单因子胁迫下,除T0D2处理外,WUE呈先降后升趋势,表明前期蒸腾速率降低幅度大于净光合速率,后期蒸腾速率降低幅度小于净光合速率,最小值出现在第5周。复合胁迫T1D1、T2D1处理下,WUE呈先降后升的趋势,T2D2处理下,WUE呈先升后降的趋势,最大值出现在第5周。复合胁迫末期,除T1D1外,T1D2、T2D1、T2D2处理分别较对照增加35.7%、28.6%和38.9%(表2)。双因素方差分析结果表明,高温以及复合胁迫对WUE影响不显著,干旱胁迫极显著影响WUE(表3)。

表3 高温和干旱胁迫下杨树幼苗生理生化指标的双因素方差分析结果
注:*表示显著性概率为Plt;0.05;** 表示显著性概率为Plt;0.01;*** 表示显著性概率为Plt;0.001。
2.2 不同处理对杨树幼苗荧光参数的影响
2.2.1对叶绿素初始荧光产量(Fo)的影响
从表4可见,高温胁迫下,Fo呈上升趋势,T1D0处理下各时期与对照差异不显著,T2D0处理下各时期显著高于对照,处理间差异显著。干旱胁迫下,Fo呈先降后升趋势,T0D1处理下Fo从第7周开始显著高于对照,T0D2处理下各时期均显著高于对照。胁迫末期T1D0、T2D0、T0D1、T0D2处理分别较对照上升5.4%、41.9%、11.1%和23.0%。复合胁迫下,除T2D2处理,Fo变化趋势与高温胁迫相似。T1D1、T1D2处理下Fo各时期显著高于单一胁迫处理。胁迫末期T1D1、T1D2、T2D1、T2D2较对照上升19.1%、32.9%、30.8%和37.7%(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫极显著影响Fo(表3)。
2.2.2对叶绿素最大荧光产量(Fm)的影响
从表4可见,高温或干旱单因子胁迫下,Fm总体上呈下降趋势,且随着胁迫程度的增加,下降幅度越大。干旱胁迫各处理间在第5、第6周差异显著,高温胁迫各处理间各个时期差异显著。胁迫末期T1D0、T2D0、T0D1、T0D2处理下,Fm分别下降了20.7%、40.3%、26.1%和26.2%。复合胁迫下,除T1D1处理,Fm变化趋势与单因子胁迫相同。胁迫末期T1D1、T1D2、T2D1、T2D2分别下降9.6%、29.7%、38.1%、40.2%(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫极显著影响Fm(表3)。
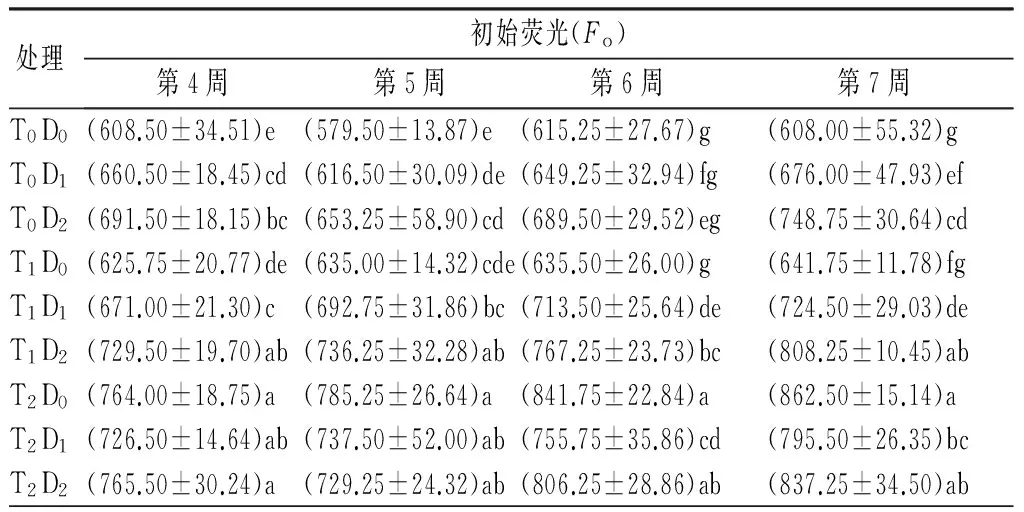
表4 高温和干旱胁迫对杨树幼苗荧光参数的影响

处理最大荧光产量(Fm)第4周第5周第6周第7周T0D0(3325.00±73.09)a(3261.00±132.32)a(3280.75±87.86)a(2944.00±45.47)aT0D1(2849.50±91.76)c(2653.25±94.95)cde(2500.50±123.84)b(2177.50±64.95)dT0D2(3060.50±112.49)bc(2447.50±114.45)f(2300.00±143.14)c(2141.00±126.93)dT1D0(3167.50±95.88)ab(3043.25±64.75)b(2608.00±102.30)b(2333.00±147.44)cT1D1(3196.00±111.35)ab(2739.75±63.72)c(2618.75±72.76)b(2661.25±73.33)bT1D2(2921.00±40.91)c(2513.50±119.76)ef(2155.00±114.55)cd(2069.00±104.09)dT2D0(2848.75±104.54)c(2686.75±60.10)cd(1873.75±161.23)e(1756.75±87.58)eT2D1(2998.00±56.53)bc(2677.00±73.15)cd(2286.25±100.89)c(1821.75±61.78)eT2D2(2917.50±298.53)c(2541.75±32.12)def(2082.25±42.69)d(1759.75±67.43)e
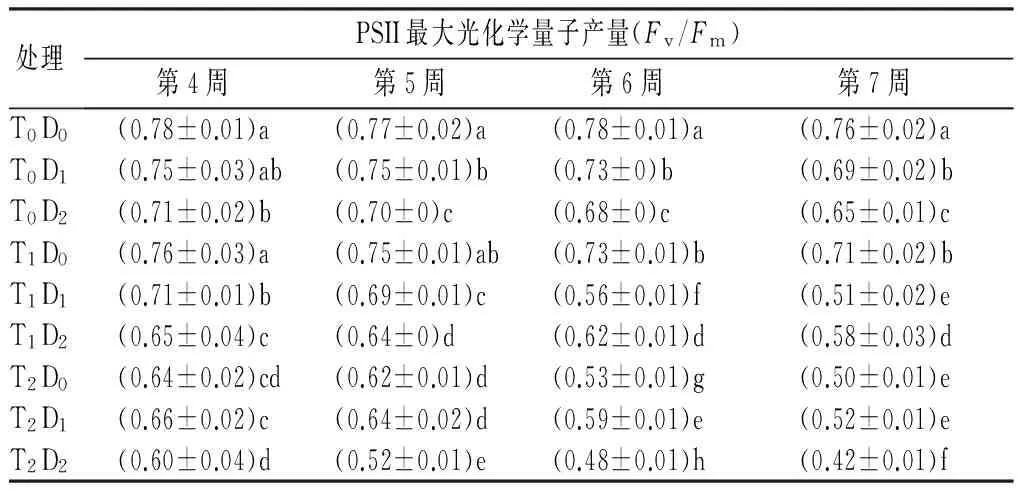
处理PSII最大光化学量子产量(Fv/Fm)第4周第5周第6周第7周T0D0(0.78±0.01)a(0.77±0.02)a(0.78±0.01)a(0.76±0.02)aT0D1(0.75±0.03)ab(0.75±0.01)b(0.73±0)b(0.69±0.02)bT0D2(0.71±0.02)b(0.70±0)c(0.68±0)c(0.65±0.01)cT1D0(0.76±0.03)a(0.75±0.01)ab(0.73±0.01)b(0.71±0.02)bT1D1(0.71±0.01)b(0.69±0.01)c(0.56±0.01)f(0.51±0.02)eT1D2(0.65±0.04)c(0.64±0)d(0.62±0.01)d(0.58±0.03)dT2D0(0.64±0.02)cd(0.62±0.01)d(0.53±0.01)g(0.50±0.01)eT2D1(0.66±0.02)c(0.64±0.02)d(0.59±0.01)e(0.52±0.01)eT2D2(0.60±0.04)d(0.52±0.01)e(0.48±0.01)h(0.42±0.01)f
注:表中数据为平均值±标准差。同列数据后不同小写字母表示处理间差异显著(Plt;0.05)。
2.2.3对叶绿素PSII最大光化学量子产量(Fv/Fm)的影响
Fv/Fm可以反映PSⅡ的潜在活性。从表4可见,高温、干旱单因子和复合胁迫下,Fv/Fm总体上呈下降趋势。T1D0、T0D1处理下从第6周开始与对照差异显著,T2D0、T0D2处理下各个时期显著低于对照,干旱胁迫处理间从第5周开始差异显著,高温胁迫在各个时期内处理间差异均显著。胁迫末期,T1D0、T2D0、T0D1、T0D2处理下,Fv/Fm分别下降7.19%、34%、9.12%、14.91%,表现为重度高温胁迫对Fv/Fm的影响大于干旱胁迫。复合胁迫下,T1D1、T1D2、T2D2在各时期显著高于单一胁迫处理,T1D1、T1D2、T2D1、T2D2处理在胁迫末期分别下降32.7%、23.9%、31.7%、45.6%,且温度越高、干旱胁迫越严重,Fv/Fm降低越显著。高温和干旱双重逆境对Fv/Fm胁迫效应由大到小依次为T2D2、T2D1、T2D0、T1D2、T1D1、T0D2、T0D1、T1D0(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫极显著影响Fv/Fm(表3)。
2.3 不同处理对杨树幼苗抗氧化酶活性的影响
2.3.1 对超氧化物歧化酶(SOD)活性的影响
从表5可见,T1D0处理下,SOD活性呈上升趋势,T2D0、T0D1、T0D2处理下SOD呈先升后降的趋势,说明一定程度的高温或干旱胁迫可以使SOD活性上升,但在严重胁迫下,SOD活性受到抑制。T0D1处理下,SOD最大值出现在第6周。T2D0、T0D2处理下,最大值出现在第5周。干旱单因子胁迫下,各时期与对照差异显著,处理间从第5周开始出现显著差异。高温单因子胁迫下,各处理时段间差异均显著。复合胁迫,T1D1、T1D2处理下SOD活性呈先升后降趋势,最大值均出现在第5周。T2D1、T2D2处理下SOD活性呈下降趋势,并从第6周开始显著低于对照。复合胁迫末期SOD活性低于单一胁迫。高温和干旱双重逆境对SOD胁迫效应由大到小依次为T2D1、T2D2、T1D2、T0D2、T2D0、T1D1、T0D1、T1D0(表2)。双因素方差分析结果表明,高温以及复合胁迫显著影响SOD,干旱单一胁迫对SOD无显著影响(表3)。
2.3.2 对过氧化物酶(POD)活性的影响
从表5可见,T1D0处理下POD活性呈上升趋势,从第6周开始显著高于对照;T2D0处理下POD呈下降趋势,从第5周开始显著低于对照;T0D1、T0D2处理下,POD呈先升后降的趋势,最大值分别出现在第6周、第5周,与SOD相同,适度的高温或者干旱对POD活性有促进作用。复合胁迫下,除T1D1处理,POD活性总体上呈下降趋势,T1D1、T1D2、T2D2复合处理POD活性在胁迫末期均低于单一胁迫处理。高温和干旱双重逆境对POD胁迫效应由大到小依次为T2D2、T2D0、T2D1、T1D1、T1D0、T1D2、T0D1、T0D2(表2)。双因素方差分析结果表明,高温、干旱以及复合胁迫显著影响POD(表3)。
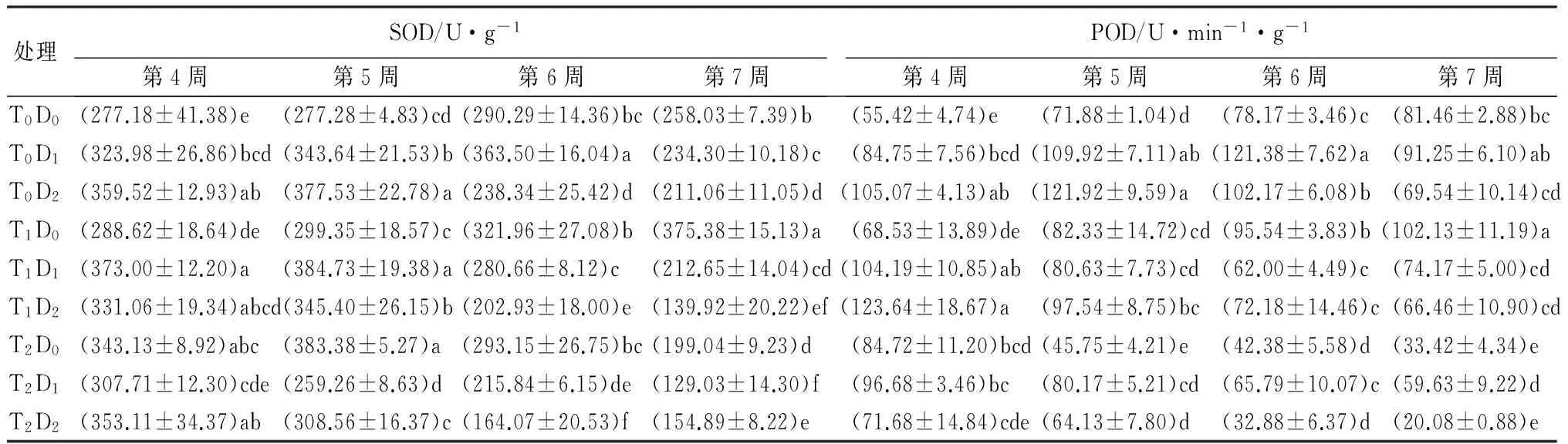
表5 高温和干旱胁迫对杨树幼苗抗氧化酶活性的影响
注:表中数据为平均值±标准差。同列数据后不同小写字母表示处理间差异显著(Plt;0.05)。
3 讨论
3.1 高温和干旱胁迫对光合特性的影响
植物光合作用的抑制包括气孔限制和非气孔限制,分别由气孔关闭和光合器官自身活性下降引起[12]。在中度高温或干旱胁迫下,Gs一直降低,Ci先降低后升高,说明中度高温和干旱胁迫下气孔和非气孔因素共同限制Pn,重度高温和干旱胁迫下,Gs下降,Ci升高,说明重度高温和干旱胁迫下Pn主要受到非气孔因素限制。韩瑞宏等[13]研究也发现紫花苜蓿在中度和重度干旱胁迫下Pn的降低是非气孔限制起着主要作用。孙宪芝等[14]研究在重度高温胁迫下,Pn在胁迫前期受到气孔限制,后期受到非气孔限制,与本研究不一致,可能是与胁迫时间长短有关。为了不降低光合速率,植物适应干旱的机制是最大限度地降低蒸腾速率[15]。本研究表明,随着气孔导度的降低,中度干旱胁迫下,蒸腾速率先升后降,重度干旱胁迫下,蒸腾速率显著下降。这与干旱胁迫下,Tr均下降的研究结果[16]不符,可能是由于在一定的胁迫时间内,植物对胁迫具有一定的抗性。有研究表明,高温胁迫下,Tr大幅度上升[17];也有研究表明,高温胁迫下,Tr显著低于对照[18]。本研究的结论与后者相同。复合胁迫下,Pn、Gs均下降,Ci上升,表明Pn主要受非气孔限制。随着胁迫时间的延长和程度的加强,Pn、Tr下降越显著,各个时期都低于单因子的水平,说明复合胁迫对杨树Pn、Tr具有叠加效应。重度高温和干旱复合胁迫末期,光合性能均低于其他处理,说明杨树幼苗在这2个处理下,受害最严重。从表1可以看出,温度对Pn、Gs、Ci的影响大于水分,对Tr的影响小于水分。说明与干旱相比,温度在植物光合作用中起主导作用。
植物水分利用效率(WUE)的高低能够反映对于逆境胁迫调节能力的大小。有研究表明在适度高温或干旱胁迫下,WUE会显著升高,而重度高温或干旱胁迫下,WUE下降[19-20]。也有研究表明,较高的温度和水分胁迫伴随着较高的WUE[21-22]。本研究结果显示,中度高温或干旱胁迫下,植物水分利用效率与对照相比无明显差异,重度高温干旱胁迫下,水分利用效率上升,这可能是由于不同的植物对逆境胁迫具有不同抗性造成的。复合胁迫下,WUE先下降后升高,说明植物在降低Pn的同时更大限度地降低了Tr,也说明南林895杨在胁迫后期产生抗性,通过提高水分利用效率来适应胁迫,提高自身的适应性。
3.2 高温和干旱胁迫对荧光参数的影响
PSII对于胁迫非常敏感,因此常用叶绿素荧光参数来判断植物是否遭受逆境胁迫[23]。有研究表明,高温、干旱或复合胁迫环境,会导致植物Fo上升、Fm、Fv/Fm下降[24]。本研究也证实了这一点。FV/Fm常被用来衡量植物PSII是否因遭受光抑制而造成损伤[25]。本研究中,中度高温和干旱胁迫时,FV/Fm在前期与对照差异不显著,后期显著低于对照,而重度高温和干旱胁迫下Fv/Fm显著低于对照,进一步说明在中度高温和干旱胁迫下,Pn是由气孔和非气孔因素共同限制,在重度高温和干旱胁迫下,Pn是由非气孔因素限制。中度高温、干旱胁迫下Fo上升幅度小于重度高温、干旱胁迫。中度高温、干旱胁迫下Fm、Fv/Fm的下降幅度小于重度高温、干旱胁迫,说明在重度高温和干旱胁迫下叶片PSⅡ的电子传递能力变弱,光能利用率降低,PSⅡ潜在活性受到抑制,叶片捕获光能转化为化学能的能力受到抑制[26-27]。同时也表明杨树在中度高温或干旱胁迫时可以通过将过剩的光能转化为化学能,来延缓光破坏进程[28]。复合胁迫下,Fo、Fm、Fv/Fm的变化趋势与单因子胁迫一致,但其对荧光参数的影响与对照相比更加显著,表明复合胁迫对PSII影响大于单一胁迫,PSII产生可逆或不可逆的损伤。由此可见,高温、干旱单一和复合胁迫下,植物叶片的PII受到不同程度的损伤,导致细胞ATP供应受阻从而影响光合作用的正常进行。
3.3 高温和干旱胁迫对抗氧化酶活性的影响
植物抗氧化酶活性的变化在植物对逆境反应的研究中应用广泛[29]。前人研究表明,逆境胁迫下,PSII活性降低会导致过量的化学能产生,MDA含量增加,从而增加质膜的相对透性,使植物体内活性氧积累,最终造成氧化伤害[30-31]。体内活性氧升高时,抗氧化酶活性也会相应升高。本研究表明,在中度高温胁迫下,SOD、POD活性上升,说明杨树对于脂质过氧化产生了防御,重度高温、中度干旱、重度干旱下,SOD、POD活性呈先升后降的趋势,且随着时间的增加,降低越显著,说明高温和干旱胁迫的时间和强度超过了抗氧化酶的适应程度,细胞膜系统受损严重,细胞内活性氧代谢失调。也说明植物在遭受高温和干旱胁迫时,抗氧化酶的活性不一定总升高,胁迫时间和强度均会对其产生影响。复合胁迫下SOD活性显著低于单因子胁迫,说明复合胁迫对SOD的影响大于单一胁迫,植物受到严重的氧化伤害。方差分析表明,温度对SOD、POD的影响高于水分,这也与Li et al.[32]对云南杨的研究结果一致。
4 结论
本研究表明,高温、干旱单一或复合胁迫使植物光合性能下降;高温胁迫对幼苗光合性能和抗氧化酶活性的影响比干旱胁迫更显著;复合胁迫对植物Pn、Gs、Ci、Tr、Fv/Fm、SOD的胁迫效应大于单一胁迫且对Pn、Tr具有叠加效应。综合考虑胁迫时间和强度发现,杨树幼苗在高温和干旱复合胁迫条件下受害最严重。
[1] 李力,刘玉民,王敏,等.3种北美红枫对持续高温干旱胁迫的生理响应机制[J].生态学报,2014,34(22):6471-6480.
[2] 李扬,黄建辉.库布齐沙漠中甘草对不同水分和养分供应的光合生理响应[J].植物生态学报,2009,33(6):1112-1124.
[3] 赵丽英,邓西平,山仑.渗透胁迫对小麦幼苗叶绿素荧光参数的影响[J].应用生态学报,2005,16(7):1261-1264.
[4] SHARMA D K, ANDERSEN S B, OTTOSEN C O, et al. Wheat cultivars selected for highFv/Fmunder heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter[J]. Physiologia Plantarum,2015,153(2):284-298.
[5] 司福艳,乔匀周,姜净卫,等.干旱高温及高浓度CO2复合胁迫对冬小麦生长的影响[J].应用生态学报,2014,25(9):2605-2612.
[6] GRIGOROVA B, VASSILEVA V, KLIMCHUK D, et al. Drought, high temperature, and their combination affect ultrastructure of chloroplasts and mitochondria in wheat (TriticumaestivumL.) leaves[J]. Journal of Plant Interactions,2013,7(3):204-213.
[7] ASRAR A A, ABDEL-FATTAH G M, ELHINDI K M. Improving growth, flower yield, and water relations of snapdragon (AntirhinummajusL.) plants grown under well-watered and water-stress conditions using arbuscular mycorrhizal fungi[J]. Photosynthetica,2012,50(2):305-316.
[8] 吴永波,叶波.高温干旱复合胁迫对构树幼苗抗氧化酶活性和活性氧代谢的影响[J].生态学报,2016,36(2):403-410.
[9] WANG G P, TIAN F X, ZHANG M, et al. The over accumulation of glycinebetaine alleviated damages to PSII of wheat flag leaves under drought and high temperature stress combination[J]. Acta Physiologiae Plantarum,2014,36(10):2743-2753.
[10] BEYER W F J, FRIDOVICH I. Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions[J]. Analytical Biochemistry,1987,161(2):559-566.
[11] 李忠光,龚明.愈创木酚法测定植物过氧化物酶活性的改进[J].植物生理学报,2008,44(2):323-324.
[12] FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33(1):317-345.
[13] 韩瑞宏,卢欣石,高桂娟,等.紫花苜蓿(Medicagosativa)对干旱胁迫的光合生理响应[J].生态学报,2007,27(12):5229-5237.
[14] 孙宪芝,郑成淑,王秀峰.高温胁迫对切花菊‘神马’光合作用与叶绿素荧光的影响[J].应用生态学报,2008,19(10):2149-2154.
[15] LIU F, ANDERSEN M N, JACOBSEN S E, et al. Stomatal control and water use efficiency of soybean (Glycinemax(L.) Merr.) during progressive soil drying[J]. Environmental amp; Experimental Botany,2005,54(1):33-40.
[16] 李立辉,王岩,胡海燕,等.初花期干旱对不同抗旱性紫花苜蓿光合特征及荧光参数的影响[J].华北农学报,2015,30(4):126-131.
[17] 刘长利,王文全,崔俊茹,等.干旱胁迫对甘草光合特性与生物量分配的影响[J].中国沙漠,2006,26(1):142-145.
[18] 黄溦溦,张念念,胡庭兴,等.高温胁迫对不同种源希蒙得木叶片生理特性的影响[J].生态学报,2011,31(23):7047-7055.
[19] FRENCH R J, SCHULTZ J E. Water use efficiency of wheat in a Mediterranean-type environment. I. The relation between yield, water use and climate[J]. Crop and Pasture Science,1984,35(6):743-764.
[20] CHESEREK J J, GICHIMU B M. Drought and heat tolerance in coffee: a review[J]. International Research Journal of Agricultural Science amp; Soil Science,2012,2(12):498-501.
[22] KAKANI V, WHEELER T, CRAUFURD P, et al. Effect of high temperature and water stress on groundnuts under field conditions[M]//Mahalingam R.Combined Stresses in Plants. Switzerland: Springer Cham,2015:159-180.DOI.https://doi.org/10.1007/978-3-319-07899-1_8.
[23] 王海珍,韩路,徐雅丽,等.胡杨异形叶叶绿素荧光特性对高温的响应[J].生态学报,2011,31(9):2444-2453.
[24] PRADHAN G P, PRASAD P V V, FRITZ A K, et al. Effects of drought and high temperature stress on synthetic hexaploid wheat[J]. Functional Plant Biology,2012,39(3):190.
[25] 朱成刚,陈亚宁,李卫红,等.干旱胁迫对胡杨PSII光化学效率和激能耗散的影响[J].植物学报,2011,46(4):413-424.
[26] 王荣荣,夏江宝,杨吉华,等.贝壳砂生境干旱胁迫下杠柳叶片光合光响应模型比较[J].植物生态学报,2013,37(2):111-121.
[27] 滕中华,智丽,宗学凤,等.高温胁迫对水稻灌浆结实期叶绿素荧光、抗活性氧活力和稻米品质的影响[J].作物学报,2008,34(9):1662-1666.
[28] 井大炜.杨树苗叶片光合特性和抗氧化酶对干旱胁迫的响应[J].核农学报,2014,28(3):532-539.
[29] 裴斌,张光灿,张淑勇,等.土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响[J].生态学报,2013,33(5):1386-1396.
[30] MARTINEAU J R, SPECHT J E, WILLIAMS J H, et al. Temperature tolerance in soybeans. I. Evaluation of a technique for assessing cellular membrane thermostability[J]. Crop Science,1979,19(1):75-78.
[31] ZHANG P P, FENG B L, WANG P K. Leaf senescence and activities of antioxidant enzymes in different broomcorn millet (PanicummiliaceumL.) cultivars under simulated drought condition[J]. Journal of Food Agriculture amp; Environment,2012,10(2):438-444.
[32] LI X, YANG Y, SUN X, et al. Comparative physiological and proteomic analyses of poplar (Populusyunnanensis) plantlets exposed to high temperature and drought[J]. Plos One,1932,9(9):e107605.
EffectsofElevatedTemperatureandDroughtonPhotosyntheticCharacteristicsandAntioxidantEnzymeSystemofPoplarSeedlings//
Zhang Yanhong, Wu Yongbo, Liu Xuan, Zhu Jiaxin
(Collaborative Innovation Center of Sustainable Forestry in Southern China of Jiangsu Province, Nanjing Forestry University, Nanjing 210037, P. R. China)//
PopulusבNanlin-895’; Elevated temperature; Drought; Photosynthetic characteristics; Antioxidant enzyme; Chlorophyll fluorescence
1)江苏省高校自然科学研究重大项目(15KJA180003);江苏省生物学优势学科建设项目。
张燕红,女,1991年12月生,江苏省南方现代林业协同创新中心(南京林业大学),硕士研究生。E-mail:906736024@qq.com。
吴永波,江苏省南方现代林业协同创新中心(南京林业大学),副教授。 E-mail:yongbowu0920@163.com。
2017年6月17日。
责任编辑:程 红。
Q948
Journal of Northeast Forestry University,2017,45(11):32-38.
An indoor simulation experiment was performed by controlling the different temperature and soil water conditions to study the single and combined stress of elevated temperature and drought on photosynthetic characteristics, chlorophyll fluorescence and the antioxidant enzymes activities inPopulusבNanlin-895’ seedling leaves. As the stress time progressed, net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), maximal fluorescence (Fm), optimal/maximal photochemical efficiency of PSⅡ (Fv/Fm) of Poplar seedlings were decreased, whereas, minimal fluorescence (Fo) was increased under the single or combined stress of elevated temperature and drought. Intercellular CO2concentration (Ci) and water use efficiency (WUE) were decreased first and increased, and the activities of superoxide dismutase (SOD) and peroxidase (POD) had the opposite trend under the moderate stress of drought. Activities of superoxide dismutase (SOD) and peroxidase (POD) were decreased significantly under the severe stress of elevated temperature or their combination at the end of stress time. Decreases in net photosynthetic rate (Pn) were the result of stomatal and non-stomatal limitation under moderate stress of elevated temperature or drought, while the result of non-stomatal limitation was under the severe stress of elevated temperature or drought or their combination. The combined stress showed synergistic effects onPnandTr. The effects of the combined stress onPn,Gs,Ci,Tr,Fv/Fmand SOD were higher than that of a single stress, and poplar seedlings suffered the worst under the combined stress of elevated temperature and drought.
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
装备制造技术(2019年12期)2019-12-25
时代英语·高一(2019年1期)2019-03-13
科技创新与品牌(2019年12期)2019-02-06
意林(2018年20期)2018-10-31
喜剧世界(2017年10期)2017-06-01
中国铸造装备与技术(2015年5期)2015-12-10
创业家(2015年3期)2015-02-27
创业家(2015年2期)2015-02-27
西南军医(2014年4期)2014-01-19
