
柞蚕微孢子虫的研究进展
2017-11-29 06:34刘中文张英华杨新峰朱绪伟
中国蚕业 2017年4期
刘中文 徐 欣 张英华 杨新峰 郭 剑 朱绪伟
(河南省蚕业科学研究院, 河南郑州 450008)
柞蚕微孢子虫的研究进展
刘中文 徐 欣 张英华 杨新峰 郭 剑 朱绪伟
(河南省蚕业科学研究院, 河南郑州 450008)
随着柞蚕微孢子虫(NosemapernyiWen et Ding)cDNA文库及EST(expressed sequence tags,表达序列标签)测序的完成,基因克隆测序、功能蛋白分析、抗体免疫等研究手段也逐渐应用于柞蚕微孢子虫的研究,并在病原的分离检测、感染发病规律、传播途径、综合防治等方面取得较大进展。介绍了柞蚕微孢子虫的分离纯化与检测、感染传播规律及其分子机制的研究进展和柞蚕微孢子虫侵染后柞蚕免疫应答的研究进展,展望了柞蚕微孢子虫的研究方向和内容。
柞蚕; 微孢子虫; 微粒子病; 分离检测; 垂直传播
柞蚕生产上的重要病害——柞蚕微粒子病,是由柞蚕种生产中的唯一检疫对象柞蚕微孢子虫(NosemapernyiWen et Ding)诱发的,易造成丝茧生产减产,并经柞蚕卵垂直传播影响种茧生产,威胁种质资源的保育及生产的安全[1]。一直以来,关于柞蚕微孢子虫的分离检测、感染发病规律、传播途径、综合防治等方面的研究不断有新的进展,直到2011年重庆师范大学XU等[2]初步鉴定了其核型,并对全基因组进行组装,但由于组装结果差异较大,全基因组草图还需进一步研究确定;2014年,沈阳农业大学完成了柞蚕微孢子虫cDNA文库的构建及EST(expressed sequence tags,表达序列标签)测序[3],使得柞蚕微孢子虫的研究在分子生物学方面有了巨大进展,同时关于柞蚕微孢子虫感染后柞蚕在转录组学和蛋白质组学的研究成为热点,为探究柞蚕对柞蚕微孢子虫的免疫应答模式、柞蚕微粒子病的发病机理和开发新的防治药剂提供了直接依据[4]。本文介绍了柞蚕微孢子虫的分离纯化与检测、感染传播规律及其感染发病分子机制的研究进展和柞蚕微孢子虫侵染后柞蚕免疫应答的研究进展,以期在阻断柞蚕微粒子病的传播以及提高柞蚕的免疫能力等方面提供理论依据,为今后应用于柞蚕生产、降低微粒子病的发病率与传播力提供帮助。
1 柞蚕微孢子虫的分离纯化与检测技术
1.1 柞蚕微孢子虫的分离纯化
微孢子虫孢子的分离纯化方法有差速离心法、蔗糖(氯化铯) 密度梯度离心法和Percoll 密度梯度离心法等多种,其中应用最多的是Percoll密度梯度离心法。姜义仁等[5]采用传统离心的方式分离得到柞蚕微孢子虫孢子,再根据Percoll溶液自行分层的特性,设计3个单一浓度和2个不连续密度梯度的Percoll NaCl溶液进行纯化试验,最终得到了折光性和均一性一致,无组织碎片的柞蚕微孢子虫孢子,最佳离心方法为25%、50%、75%、100%不连续梯度,15 000 r/min下离心30 min,NaCl溶液浓度为0.15 mol/L。
1.2 柞蚕微孢子虫的检测技术
目前,快速免疫检测技术、酶学及分子生物学研究方法已广泛应用于病原微生物的检测鉴定,如抗体检测技术、酶联免疫吸附剂测定法(ELISA法)等技术。
1.2.1 抗体检测技术 王伯阳等[6]通过制备柞蚕微孢子虫多克隆抗体,建立了柞蚕微孢子虫胶体金免疫层析快速检测法,但由于抗体效价、柞蚕血淋巴本身具有颜色等原因导致检测灵敏度较低,后期希望配合亲和性更高的单克隆抗体优化完善检测方法。
1.2.2 ELISA法 ELISA法测定因具有较多优点而在生物科学各领域广泛应用,姜义仁等[7]也用间接竞争ELISA法检测柞蚕微孢子虫,特异性及灵敏度较好,为开发柞蚕微孢子虫单克隆抗体、解决检测假阳性、提高检测灵敏度及特异性等方面奠定了基础。但ELISA法仍存在抗体捕获孢子的能力较弱、灵敏度的稳定性较差等不足之处,实用化检测技术有待进一步研究[8]。
2 柞蚕微孢子虫的感染传播规律及其分子机制研究进展
2.1 具有致病性柞蚕微孢子虫种类
目前发现对柞蚕具有感染致病性的微孢子虫有5种[9],包括柞蚕微孢子虫、修氏内网虫(Endoreticlatusschubergi)、链孢变形孢虫(VairimorphachainsporumWen et Ding)、讷卡变形孢虫(Vairimorphanecatrix)和柞蚕微孢子虫新种(Nosemasp.),其中,柞蚕微孢子虫是优势种。
根据宿桂梅等[10]的文献,可以从主要侵染部位及顺序、主要寄主范围、孢子大小与形状、极丝绕核圈数、孢子母细胞的产孢形式等方面对5种微孢子虫进行分类鉴定,为研究各种微孢子虫的致病性做准备。在此基础上,徐亮等[11]比较5个柞蚕品种的抵抗能力发现,柞蚕品种582的幼虫生长发育受微孢子虫侵染的影响最小,柞蚕品种青6号和宽青的幼虫生长发育受微孢子虫侵染的影响最大,为选育对微孢子虫侵染具有较强抵抗力的柞蚕品种提供了更充分和更准确的依据。
2.2 柞蚕微孢子虫的感染传播规律
柞蚕微孢子虫的生命周期一般分为感染期、裂殖增殖期和孢子增殖期3个阶段[12]。孢子阶段是微孢子虫生活史中成熟且具有侵染力的阶段,也是研究微孢子虫侵染机制、生活史及检测诊断的关键阶段。与微孢子虫侵染有关的重要结构有3个:极膜体、极丝及后极泡,其中与侵染联系最紧密的细胞器是极丝[12]。目前至少有5种极丝蛋白被鉴定,这些蛋白在微孢子虫种间具有一定的保守性,在与宿主细胞粘附及侵染的最后阶段可能发挥作用[13];极丝蛋白还具有用于开发血清学检查抗原的潜力[14]。可以根据微孢子虫极丝蛋白的研究克隆得到柞蚕微孢子虫的相关极丝蛋白,为进一步研究其侵染机制提供理论依据。
关于微孢子虫感染细胞途径的研究一直在进行,目前认为主要有2种方式:一是孢子发射极丝注射孢原质到宿主细胞以达到感染的目的,有的同时形成寄生空泡再次感染[15];二是通过微孢子虫孢壁的主要成分孢壁蛋白与宿主细胞的相互作用,来实现微孢子虫对宿主的侵染,这也是病原体免疫检测的重要靶标蛋白[16],关于柞蚕微孢子虫孢壁蛋白的研究也逐渐成为热点。
柞蚕微孢子虫的传播途径有水平传播和垂直传播2种[1]。水平传播即经口感染,柞蚕微孢子虫进入到柞蚕体内后,微孢子虫不断增殖,吸收了蚕体的大量营养,导致柞蚕生理机能发生障碍;垂直传播即胚种传染,是柞蚕微孢子虫的主要传染途径,也是造成检测及防治难度较大的原因[17]。关于柞蚕微孢子虫致病性与传播规律相互关系的研究还比较少,但已有一些研究发现,病原的毒力和垂直传播特点具有一定的关系,病原的毒力越强,胚种垂直传染力越弱,反之亦然[18]。同时,在家蚕中发现,对家蚕具有病原性的不同微孢子虫所表现出的垂直传播特点不一致;另外,研究人员还从山东蚕区分离出一种纳卡变形微孢子虫,虽对家蚕具有较强的致病性[19],但不具有垂直传播的能力[20]。
2.3 侵染相关分子生物学方面的研究进展
龚娟娟等[21]利用Percoll-NaCl梯度纯化和LC-MS/MS(液相色谱串联质谱)鉴定技术,得到了柞蚕微孢子虫的610个蛋白,再进行GO蛋白质功能分析,发现多种与侵染、增殖相关的功能蛋白,为研究其侵染机制提供了分子依据。
孢壁蛋白在信号识别、细胞粘附和离子转运等方面起着重要作用,SOUTHERN等[22]通过研究功能基因EnP1,证实了孢壁蛋白是病原菌侵染过程中的重要蛋白,还发现兔脑炎微孢子虫孢壁蛋白EnP1和家蚕微孢子虫表面蛋白NbSWP26有相似的功能[22-23]。同时,还有研究发现微孢子虫孢壁的某些蛋白质被破坏,如利用抗体或其他化学物质处理微孢子虫孢子,其孢子的感染能力显著下降[24]。
由于微孢子虫特有的几丁质孢壁结构,常规的蛋白质提取方法难以获得仅含微孢子虫表面蛋白的检测样品,而水溶性维生素生物素和糖蛋白亲和素相结合的层析分离技术可特异性标记和富集细胞表面蛋白,具备灵敏、稳定、专一、适用广泛等特点[25-27]。赵唯希等[28]运用生物素-亲和素层析分离技术得到柞蚕微孢子虫表面蛋白,并与家蚕微孢子虫表面蛋白进行比对,已鉴定出3个柞蚕微孢子虫孢子表面蛋白NpSWP30、NpSWP26、NpSWP12,并通过间接免疫荧光亚细胞定位分析确证为孢子外表面的蛋白质,为进一步研究柞蚕微孢子虫孢壁蛋白细胞定位以及功能奠定了基础,也为检测和侵染机制的研究提供了新的靶标蛋白。
3 柞蚕微孢子虫侵染后柞蚕免疫应答的研究进展
柞蚕微孢子虫的侵染对柞蚕体内各种生理生化指标均造成不同程度的影响,主要表现在中肠和血淋巴的一系列反应中。宿桂梅等[10]探究了不同温度条件对柞蚕微孢子虫侵染组织顺序及发病时间的影响不同,验证了其是通过血液循环侵染其他组织,并发现侵染的过程都是从柞蚕中肠开始的。
3.1 柞蚕微孢子虫侵染后柞蚕中肠相关免疫反应
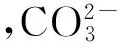
基于柞蚕微孢子虫侵染后柞蚕中肠的重要作用,姜义仁等[12]构建了柞蚕中肠蛋白质数据库,分析筛选得到差异基因和差异蛋白所在的位置和参与的代谢,并发现与柞蚕微孢子虫发病相关的2种蛋白——丝氨酸蛋白酶和芳基贮存蛋白,为进一步阐明柞蚕微粒子病的发病机制奠定了基础。之后,王勇等[32]运用qRT-PCR(定量即时聚合酶链锁反应)技术对柞蚕中肠内ApHSC70基因进行研究,发现其表达量在微孢子虫感染后最高可上调近5倍,推测该基因可能参与了柞蚕的免疫防御过程。
3.2 柞蚕微孢子虫侵染后柞蚕血淋巴相关免疫反应
相关研究认为血细胞总数的增加及蛋白含量的减少可以作为衡量柞蚕微粒子病的标志[33],说明柞蚕血淋巴在柞蚕免疫系统中起重要作用,能够抵抗柞蚕微孢子虫的入侵。臧敏等[34]通过添食柞蚕微孢子虫发现,血淋巴蛋白质含量和组成发生显著变化,影响柞蚕体内营养代谢并且雌雄个体存在抗性差异,说明柞蚕微孢子虫寄生是一个相对缓慢的侵染过程。还有研究证明,热休克蛋白HSC70在免疫调节、炎症反应以及抗病原菌感染过程中起重要作用,参与多种生物的免疫应答反应[35],这可以作为研究柞蚕微孢子虫侵染后柞蚕免疫反应的一个依据和方法。
4 研究展望
柞蚕微孢子虫是生产上最具威胁的病原之一,与产业发展紧密关联,关于其基础理论的研究成果应该与蚕业生产实践结合,以推动产业升级与发展。柞蚕微孢子虫检测技术的研究提高了实际生产中微粒子病的检出率,并能实现柞蚕微粒子病早发现、早处理,降低得病率与损失。但对于寄生柞蚕的5种微孢子虫的研究只集中在柞蚕微孢子虫的研究,而关于其他4种微孢子虫致病力与胚种传染率的异同还有待进一步研究,如可以利用新技术分离纯化各种微孢子虫,为微粒子病的综合治理提供帮助;同时在mRNA水平和蛋白质水平进一步研究柞蚕免疫相关基因,为提高柞蚕抗病性提供分子学依据。
柞蚕微孢子虫的水平传播需要在柞蚕场生产过程中加强防病意识,做好消毒工作,降低相互传染的几率;垂直传播是造成柞蚕微粒子病病原污染的重要原因,而且对柞蚕种的种质检测工作造成诸多麻烦,因此亟需对其进行相关机制研究,以期可以找到阻断柞蚕微孢子虫垂直传播的方法和药剂,达到阻断柞蚕微孢子虫垂直传播的目的。利用现代分子生物学技术,进行柞蚕抗性育种素材的创建,具有广阔的研究前景,今后可以利用已发现的与侵染相关的极丝蛋白、孢壁蛋白的结构与功能差异,测定免疫相关基因的表达量变化,以揭示寄生柞蚕不同微孢子虫垂直传播的机理。还可以寻找阻断垂直传播的分子靶标,构建相关的单克隆抗体,再运用RNA干扰或转基因技术研究柞蚕抗微粒子病新品种。
综合柞蚕微孢子虫近几年的研究发展情况,尽管其核型已获得鉴定,然而具体的遗传规律还不清楚,详细的基因图谱结果还没有获得,相关基因在微孢子虫侵染过程中和宿主的免疫过程中的功能还需深入研究。未来开展对柞蚕微孢子虫侵染和垂直传播机制的研究,并建立便捷、灵敏的检测方法和创建柞蚕高抗病育种素材将是重要的研究方向。
[1] 秦利.中国柞蚕学[M].北京:中国科学文化出版社,2003:347-352.
[2] XU J S,WANG L J,TANG F H,et al.The nuclear apparatus andchromosome DNA in microsporidianNosemaantheraeae[J].J Eukary Microbiol,2011,58(2):178-180.
[3] 王勇,姜义仁,王晓惠,等.柞蚕微孢子虫全长cDNA文库的构建及EST分析[J].蚕业科学,2014,40(2):265-271.
[4] 王勇.柞蚕微孢子虫EST分析、孢壁蛋白NpSWP1基因克隆及柞蚕免疫应答研究[D].沈阳:沈阳农业大学,2014.
[5] 姜义仁,邓真华,王伯阳,等.柞蚕微孢子虫孢子分离纯化方法[J].应用昆虫学报,2011,48(2):452-456.
[6] 王伯阳,姜义仁,臧敏,等.柞蚕微孢子虫胶体金免疫层析检测法[J].蚕业科学,2012,38(1):97-101.
[7] 姜义仁,王伯阳,孙影,等.间接竞争ELISA法对柞蚕微孢子虫的检测[J].应用昆虫学报,2013,50(5):1 364-1 370.
[8] 鲁兴萌,蔡顺风,邱海洪,等.蚕病检测中免疫学技术的研究与应用[J].蚕桑通报,2011,42(3):1-4.
[9] 丁杰,宿桂梅,问锦曾.中国柞蚕微粒子病病原研究续报[J].蚕业科学,1994,20(3):161-165.
[10] 宿桂梅,丁杰.柞蚕微孢子虫对柞蚕组织细胞的侵染及组织病变观察[J].北方蚕业,2003,24(3):4-35.
[11] 徐亮,陈悦,孟宪民,等.柞蚕微孢子虫侵染对不同柞蚕品种幼虫生长发育的影响[J].蚕业科学,2014,40(1):172-175.
[12] 姜义仁.柞蚕微孢子虫检测及感染柞蚕中肠转录组及蛋白组学研究[D].沈阳:沈阳农业大学,2012.
[13] POLONAIS V,DELBAC F,VIVARES C.Polar tube proteins in microsporidia[J].Protistology,2007(5):63.
[14] PEEK R,DELBAC F,SPEIJER D,et al.Carbohydrate moieties od microsporidian polar tube proteins are targeted by immunoglobulin G in inmmunocompetent individuals[J].Infect Immun,2005(73):7 906-7 913.
[15] CASPAR F.How do microsporidia invade cell?[J].Folia Parasitol,2005(52):36-40.
[16] BILLER L,MATTHIESEN J,KÜHNE V,et al.The cell surface proteome ofEntamoebahistolytica[J].Mol Cell Proteomics,2014,13(1):132-144.
[17] 汪方炜,鲁兴萌.昆虫的微孢子虫病[J].昆虫知识,2003,40(1):5-8.
[18] REID A J,VERMONT S J,COTTON J A,et al.Comparative genomics of the apicomplexan parasitesToxoplasmagondiiandNeosporacaninum:Coccidia differing in host range and transmission strategy[J/OL].Plos Pathog,2012,8(3):e1002567.
[19] LIU H D,PAN G Q,LI T,et al.Ultrastructure,chromosomal karyotype,and molecular phylogeny of a new isolate of microsporidian Vairimorpha sp.BM(Microsporidia,Nosematidae) fromBombyxmori in China[J].Parasitol Res,2012,110(1):205-210.
[20] LUO B,LIU H D,PAN G Q,et al.Morphological and molecular studies ofVairimorphanecatrixBM,a new strain of the microsporidium V.necatrix(Microsporidia,Burenellidae)recorded in the silkworm,Bombyxmori[J].Exp Parasitol,2014(143):74-82.
[21] 龚娟娟,许金山,李治,等.柞蚕微孢子虫长极丝型孢子总蛋白的鉴定及功能分析[J].重庆师范大学学报,2014,31(3):26-30.
[22] SOUTHERN T R,JOLLY C E,LESTER M E,et al.EnP1,a microsporidian spore wall protein that enables spores to adhere to and infect host cells invitro[J].Eukaryot Cell,2007,6(8):1 354-1 362.
[23] LI Y H,WU Z L,PAN G Q,et al.Identification of a novel spore wall protein (SWP26)from microsporidiaNosemabombycis[J].Int J Parasitol,2009,39(4):391-398.
[24] 李治.家蚕微孢子虫(Nosemabombycis)孢壁蛋白SWP5的研究[D].重庆:西南大学,2012.
[25] RYBAK J N,ETTORRE A,KAISSLING B,et al.In vivo protein biotinylation for identification of organ specific antigens accessible from the vasculature[J].Nat Methods,2005,2(4):291-298.
[26] 孔令青,李勇,高洪,等.生物素-亲和素标记技术[J].动物医学进展,2008,29(4):100-102.
[27] SEARS K T,CERAUL S M,GILLESPIE J J,et al.Surface proteome analysis and characterization of surface cell antigen (Sca) or autotransporter family ofRickettsiatyphi[J].Plos Pathogens,2012,8(8):307-309.
[28] 赵唯希,王林玲,刘泽锋,等.应用生物素标记和蛋白质组学方法分离鉴定柞蚕微孢子虫(Nosemapernyi)的表面蛋白[J].蚕业科学,2015,41(4):669-676.
[29] 宿桂梅,李亚洁.微粒子感染对柞蚕体内酶活性及血淋巴蛋白质含量的影响[J].辽宁农业科学,2002(1):46-47.
[30] 刘吉平,汤宇杰,王玉强,等.柞蚕感染微孢子虫前后的酪氨酸活力变化的研究[C]//中国蚕学会,内蒙古自治区蚕学会,中国农业科学院蚕业研究所,等.中国蚕学会第六届家蚕和柞蚕遗传育种学术研讨会论文集.海拉尔:内蒙古自治区呼伦贝尔市蚕业科学研究所,2009:319-324.
[31] 王爱荣,李健男,刘向国,等.柞蚕微孢子虫孢子人工发芽的研究[J].沈阳农业大学学报,2002,33(4):281-284.
[32] 王勇,刘微,王斌赫,等.qRT-PCR检测柞蚕中肠热休克蛋白基因ApHSC70对微孢子虫入侵的响应[J].蚕业科学,2015,41(2):300-307.
[33] MADHUSUDHAN K N,NUNGSHI-DEVI C,LOKESH G,et al.Impact ofNosemamylitta(pebrine) infection on the larval parameters, protein concentration and total hemocyte level in daba ecorace ofAntheraeamylittaD.(tropical tasar silkworm)[J].Microbiol J,2011,1(3):97-104.
[34] 臧敏,秦萍,王勇,等.柞蚕微孢子虫侵染后柞蚕5龄雌雄个体血淋巴蛋白质含量和组成的变化及差异分析[J].蚕业科学,2012,38(1):92-96.
[35] 刘志刚,张其中,张占会,等.近江牡蛎HSC70基因对溶藻弧菌感染的反应[J].中国水产科学,2012,19(3):500-508.
2017-07-13;接受日期2017-09-27
信息: 刘中文(1987—),男,山东沂水,本科,助理农艺师。Tel: 0377-66661675, E-mail: 1020350426@qq.com
信息: 朱绪伟(1968—),男,河南信阳,硕士研究生,高级农艺师。
Tel: 0377-66666115, E-mail: hnsckyzxw@126.com
S885.1
B
1007-0982(2017)04-0043-05
猜你喜欢
中国蚕业(2022年1期)2022-12-07
当代水产(2022年1期)2022-04-26
辽宁丝绸(2022年1期)2022-03-29
中国农业科学(2021年23期)2022-01-14
昆虫学报(2021年3期)2021-05-11
四川蚕业(2021年4期)2021-03-08
中国瓜菜(2020年8期)2020-09-26
四川蚕业(2020年2期)2020-07-10
四川蚕业(2020年2期)2020-07-10
四川蚕业(2020年4期)2020-02-10
