
模拟喀斯特生境土壤干旱胁迫对蚬木苗木的影响1)
2017-12-27 01:12欧芷阳庞世龙蒙芳谭长强郑威曹艳云申文辉
东北林业大学学报 2017年12期
欧芷阳 庞世龙 蒙芳 谭长强 郑威 曹艳云 申文辉
(广西壮族自治区林业科学研究院,南宁,530002)
模拟喀斯特生境土壤干旱胁迫对蚬木苗木的影响1)
欧芷阳 庞世龙 蒙芳 谭长强 郑威 曹艳云 申文辉
(广西壮族自治区林业科学研究院,南宁,530002)
为揭示喀斯特土壤干旱环境下植物的生长及形态适应性,模拟构建了喀斯特“土壤层-岩石层-岩溶水层”水分供应装置,以2年生蚬木(Excentrodendronhsienmu)苗木为试验材料,研究了岩溶水层岩溶水分(有/无水分)和土壤水分2个因素对蚬木幼苗生长及形态发育的影响。结果表明:岩溶水分和土壤水分极显著影响蚬木幼苗生长,苗木净生长量随土壤干旱增强显著下降,岩溶水层有水处理下(W)苗木净生长量显著高于无水处理(N)。两者极显著影响着苗木总生物量及茎、叶生物量,岩溶水分极显著影响根生物量;生物量随干旱胁迫增强显著下降,W处理的显著高于N处理。土壤水分极显著影响着根冠比和根、茎生物量比,岩溶水分显著影响茎生物量比,因素间交互作用显著影响根冠比和根生物量比、极显著影响着茎生物量比。中度和重度干旱下根冠比及根生物量比显著高于对照,茎生物量比随干旱增强显著下降,W处理茎生物量比显著高于N处理。两个因素极显著影响苗木地径、株高、根长等,岩溶水分显著影响叶片数和一级分枝数、土壤水分的影响达极显著水平,因素间交互作用对地径产生显著影响、且极显著影响叶面积。蚬木幼苗生长受土壤水分条件影响较大,土壤干旱胁迫下表现为生长下降、生物量分配和形态发育发生改变等适应策略,岩溶水分对苗木的影响与土壤水分条件密切相关。
干旱胁迫;蚬木;生长特性;生物量分配;叶面积
我国西南喀斯特地区虽然降雨丰富,但由于降雨时间分布的不均匀性、喀斯特地貌特征形成的地表渗漏性强以及土壤浅薄、保水能力差等多种因素的综合作用[1],导致临时性土壤干旱的经常形成,限制了植物在该区域的生长、分布及植被恢复。认识喀斯特适生植物对干旱生境的适应机制可为石灰岩退化山地的植被恢复提供科学依据,因此,喀斯特适生植物对土壤干旱胁迫的响应成为近年来的关注重点[2-6],研究者从植物在干旱胁迫下的生理生态适应性[7-8]、苗木结构特征[9-10]以及叶绿素荧光特性变化[11]等方面进行了深入研究。喀斯特地貌特殊的二元水文地质结构常形成周期性土壤水分短缺[12],地表水向下渗漏至岩石裂隙或表层岩溶带,或汇入地下河流,这些水分能否被植物利用?Nie等[13]采用稳定氢氧同位素测定了喀斯特植物的水分来源,发现在干旱季节植物可较多地利用岩溶裂隙水分及岩溶地下水。丁亚丽等[14]的研究显示喀斯特深层土壤同位素值整体较稳定且明显区别于浅层,在旱季上坡尾巨桉(Eucalyptusurophylla×E.grandis)主要利用较深层水分、下坡依赖浅层土壤水分,这些研究说明喀斯特地区表层土壤的水分亏缺未必限制植物生长。为探寻植物在喀斯特干旱生境下的适应性,张中峰等[15-16]设计了一套模拟野外喀斯特生境的装置,利用这套装置对喀斯特生境干旱胁迫下的青冈栎(Cyclobalanopsisglauca)苗木进行了研究,发现在岩溶水层无水条件下,青冈栎的生长及生理变化受干旱胁迫影响显著。
蚬木(Excentrodendronhsienmu)是热带石灰岩特有植物、桂西南石山季节性雨林的优势成分,其根系非常发达,适应性强,具有良好的水土保持及减少地质灾害发生功能,常作为桂西南石漠化地区植被恢复的先锋树种。研究蚬木在喀斯特干旱生境下的适应策略,可为退化喀斯特生态系统的植被恢复提供理论依据。
1 材料与方法
试验地点位于广西壮族自治区南宁市北效老虎岭附近,属南亚热带季风气候区,年平均气温21 ℃,全年积温7 500 ℃,年降水量为1 200~1 500 mm,年蒸发量1 250~1 600 mm,相对湿度80%[17]。以2年生蚬木实生苗为试验材料,平均株高(48.85±6.15)cm、平均地径为(6.68±0.91)mm,长势基本一致。2016年3月底,将蚬木幼苗移栽到高100 cm、上口径65 cm、下口径为45 cm大小的白色塑料大桶内,每桶种植4株,放入遮雨棚内。供试基质来自于广西马山县喀斯特山地的棕色石灰土,其基本理化性状为:土壤密度1.36 g·cm-3,最大田间持水量32.1%,pH=7.30,有机质质量分数53.90 g·kg-1,全氮质量分数3.77 g·kg-1,全磷质量分数3.84 g·kg-1,全钾质量分数13.16 g·kg-1,碱解氮质量分数253.96 mg·kg-1,速效磷质量分数30.26 mg·kg-1,速效钾质量分数98.97 mg·kg-1。
1.1 实验设计与处理
参考张中峰等[15]的方法构建模拟装置,研究蚬木幼苗在模拟喀斯特干旱生境下的生长及形态响应。模拟装置主体由上口径65 cm、下口径45 cm、高100 cm的白色塑料大桶构成。桶下层由大石灰岩石块堆砌形成的大空隙,用以模拟岩溶水层,厚度约为40 cm;中层为由石灰岩碎石砾构成的岩石层,厚度约为25 cm;上层为约25 cm厚的土壤层。在表层土壤种植蚬木幼苗,植物正常生长2个月后开始进行土壤水分控制试验。
试验设岩溶水层有/无水分(以下简称岩溶水分)和土壤层水分(以下简称土壤水分)2个处理,岩溶层水分为有水(W)和无水(N)2个水平,土壤水分为对照(CK,每隔1 d浇水1次,每次浇水至饱和)、轻度胁迫(LS,每7 d浇水1次,每次浇水至饱和)、中度胁迫(MS, 每14 d浇水1次,每次浇水至饱和)、重度胁迫(SS,每21 d浇水1次,每次浇水至饱和)4个水平。2个处理共设置8组,即WCK、WLS、WMS、WSS、NCK、NLS、NMS、NSS,每组3个重复。在对土壤层进行水分控制试验180d后,采集样品进行各项指标测定。
1.2 测定指标及方法
土壤相对含水量采用烘干法进行测定。不同梯度的土壤含水量见表1。
幼苗存活率=(处理后幼苗存活数/处理前幼苗数)×100%。
生长指标测定:干旱胁迫开始前及干旱胁迫第45、90、135和180 d时,记录植株叶片数,用精确至0.01 mm的游标卡尺测量植株地径,用直尺测量株高(精确至0.1 cm)。 干旱胁迫结束时,每组随机挑选3株幼苗,测定其叶面积,每株随机测定10片叶片,仪器为CI-203手持式叶面积仪。然后,每组随机收割4株幼苗,统计每株的一级须根数、一级分枝数、叶片数,用卷尺(精度为0.1 cm)测量株高、分枝长、根长,用精度为0.01 mm的游标卡尺测量地径。
生物量测定:用精度为0.001 g的电子天平分别称量收割植株根、茎、叶鲜质量,放入烘箱中经105 ℃烘15 min杀青后,温度调至75 ℃烘至恒质量后称量各部位干质量。

表1 不同处理组的土壤含水量
1.3 数据处理与分析
根据以下公式计算下列参数:根生物量比(RMR)=根生物量/整株生物量;茎生物量比(RBM)=茎生物量/整株生物量;叶生物量比(RLM)=叶生物量/整株生物量;根冠比=根生物量/地上生物量。
采用双因素方差分析检验岩溶水分和土壤水分两个因素对苗木的影响,采用成对比较和Duncan’s法对数据进行比较,显著性检验水平为a=0.05。数据统计分析由SPSS 19.0软件完成,采用Simaplot 10.0软件进行作图。
2 结果与分析
2.1 干旱胁迫对蚬木幼苗成活率的影响
供试苗木在不同干旱条件下生长180 d后,所有处理组的成活率均为100%。W处理组从中度胁迫才开始出现生长缓慢现象, N处理组干旱胁迫下出现了生长缓慢,新叶卷曲、老叶枯萎及凋落等现象。
2.2 干旱胁迫对蚬木幼苗生长的影响
由图1可知,WCK叶片数、株高和地径的生长最快,WLS其次。受干旱胁迫的影响,NSS生长最慢,NMS其次。WSS的生长也受到一定影响,但总体上以岩溶水层无水条件下的干旱胁迫组受影响较大。
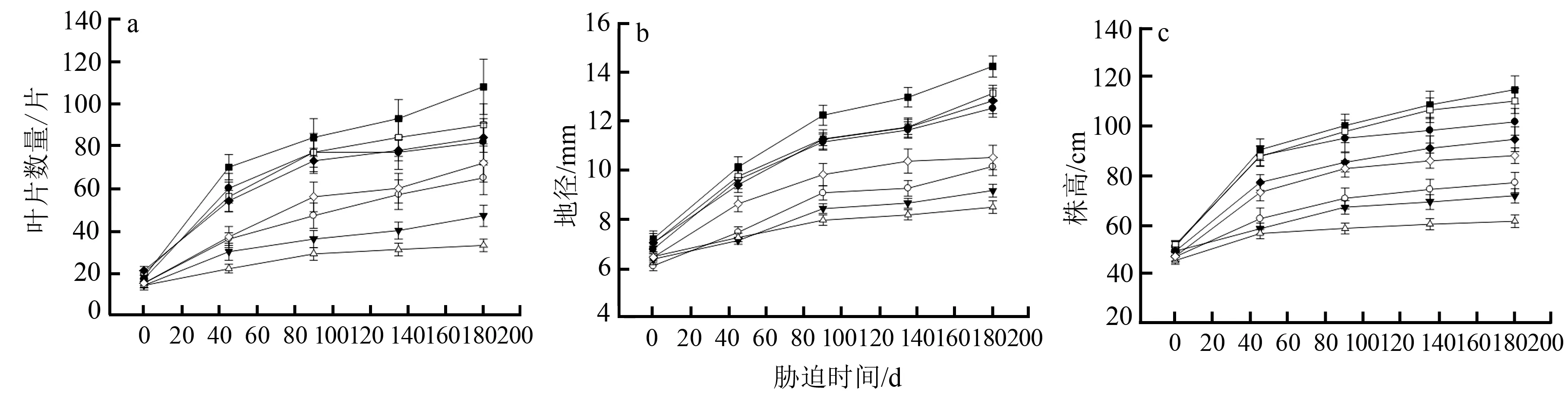
—●—NCK;—○—NLS;—▼—NMS;—△—NSS;—■—WCK;—□—WLS;—◆—WMS;—◇—WSS。
表2显示,岩溶水分和土壤水分对蚬木幼苗的叶片数、地径和株高净生长量均具有极显著影响,二者的交互作用影响不显著。表3多重比较(Duncan’s法)结果显示,蚬木幼苗的叶片数、株高和地径净生长量随土壤干旱胁迫程度增强而下降,干旱胁迫组显著低于对照。其中,苗木地径受影响最大,株高其次,叶片数受影响最小。由表3可知, 岩溶水层有水(W水平)情况下,苗木叶片数、地径和株高净生长量均显著高于无水(N水平)处理组。
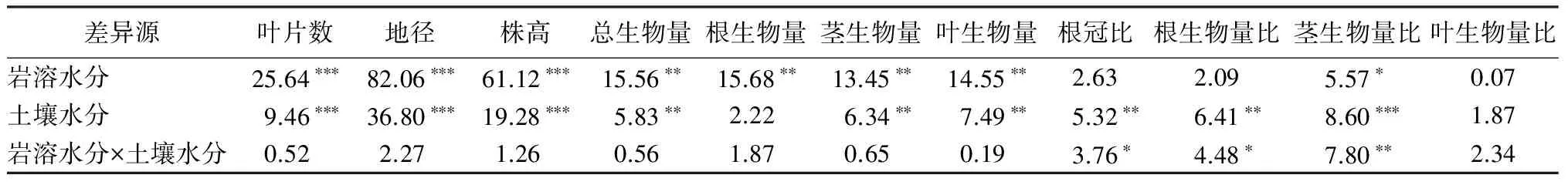
表2 不同岩溶水分和土壤水分条件对蚬木幼苗净生长量、生物量及生物量分配的方差分析(F值)
注:*表示P<0.05水平差异显著;** 表示P<0.01水平差异显著;*** 表示P<0.001水平差异显著。

表3 不同水分条件下蚬木幼苗净生长量的比较
注:表中数据为平均值±标准差,同列数据后不同字母表示差异显著(P<0.05)。
2.3 干旱胁迫对蚬木幼苗生物量积累和分配的影响
表2显示,岩溶水分对蚬木幼苗的总生物量及各部位生物量产生极显著影响,土壤水分极显著影响着总生物量及茎、叶生物量,因素间交互作用对苗木生物量无显著影响。岩溶水分显著影响茎生物量比,土壤水分极显著影响着根冠比及根、茎生物量比;因素间交互作用显著影响着根冠比和根生物量比,对茎生物量比具有极显著影响。多重比较显示(表4),苗木总生物量及根、茎、叶生物量随土壤干旱胁迫增强而下降,重度干旱胁迫下显著低于对照;根冠比和根生物量比随干旱胁迫增强而增加,茎生物量比随干旱胁迫增强而下降,叶生物量比没有显著差异。由表5可知,岩溶水层有水(W水平)条件下的生物量显著低于无水处理组(N水平);除W水平茎生物量比显著高于N水平外,根冠比及根、叶生物量比差异不显著。

表4 不同土壤水分条件下蚬木幼苗生物量及生物量分配的比较
注:表中数据为平均值±标准差,同列数据后不同字母表示差异显著(P<0.05)。

表5 不同岩溶水分条件下蚬木幼苗生物量及生物量分配的比较
注:表中数据为平均值±标准差,同列数据后不同字母表示差异显著(P<0.05)。
2.4 干旱胁迫对蚬木幼苗形态特征的影响
表6显示,岩溶水分和土壤水分对蚬木幼苗地径、株高、根长,累加分枝长以及叶面积具有极显著影响;岩溶水分显著影响着叶片数和一级分枝数,土壤水分对它们的影响达极显著水平,两个因素对苗木一级须根数影响不显著;因素间交互作用对幼苗地径产生显著影响、对叶面积产生极显著影响。多重比较显示(表7),不同土壤水分条件下,除一级须根数差异不显著之外,其余7个形态特征值基本随干旱胁迫程度增强而降低,重度干旱胁迫下的数值显著低于对照。岩溶水分两个水平间的比较显示(表8),除一级须根数之外,岩溶水层有水情况下蚬木幼苗的形态特征值均显著高于无水处理组。

表6 不同岩溶水分和土壤水分条件对蚬木幼苗形态特征的方差分析(F值)
注:*表示在P<0.05水平差异显著;** 表示在P<0.01水平差异显著;*** 表示在P<0.001水平差异显著。

表7 不同土壤水分条件下蚬木幼苗形态特征的比较
注:表中数据为平均值±标准差,同列数据后不同字母表示差异显著(P<0.05)。

表8 不同岩溶水分条件下蚬木幼苗形态特征的比较
注:表中数据为平均值±标准差,同列数据后不同小写字母表示差异显著(P<0.05)。
3 结论与讨论
3.1 干旱胁迫对蚬木幼苗生长的影响
土壤干旱胁迫影响着植物个体的形态发育,抑制植物正常生长。本研究中,岩溶水分和土壤水分对蚬木幼苗生长都具有极显著影响,苗木叶片数、地径和株高净生长量随干旱胁迫程度增强显著下降,地径受影响最大,叶片数受影响最小。研究显示土壤干旱胁迫会限制植株叶片数目的增长[18],抑制植物苗高和地径的生长[19]。由于与植物吸收运输水分过程有关,干旱胁迫下植物器官受抑制由大到小表现为根、茎、叶[20]。本研究蚬木幼苗也具有同样的规律。岩溶水层有水情况下,蚬木幼苗的叶片数、地径和株高净生长量显著高于无水处理组。张中峰等[16]发现在岩溶水层有水时,青冈栎幼苗地径生长量高于无水处理组,岩溶水层无水条件下的土壤干旱显著抑制了青冈栎幼苗的地径生长。Nie等[13]的研究证实了干旱季节喀斯特地区的植物主要利用岩石裂隙水分。这些研究表明在土壤干旱条件下,植物可通过根系利用岩溶水层水分,植株生长受影响相对较小,土壤干旱时下层岩溶水分对植物生长十分重要。
3.2 干旱胁迫对蚬木幼苗生物量积累及分配的影响
土壤干旱胁迫对植物的影响还体现在生物量积累及分配上。本研究岩溶水分和土壤水分极显著影响着蚬木幼苗的总生物量及茎、叶生物量,岩溶水分显著影响根生物量;苗木根冠比及根、茎生物量比主要受土壤水分所影响,因素间交互作用对它们也具有显著影响。干旱胁迫影响着蚬木幼苗生物量分配,根冠比和根生物量比随干旱胁迫增强而增加,茎生物量比随干旱胁迫增强而下降。植物在逆境下具有较强的自我调节能力,土壤干旱时通常将更多生物量分配至根系、提高根冠比来适用土壤层的水分亏缺[21-23];多数喀斯特适生植物在土壤干旱条件下根冠比增加[6,24-25],有的则通过增加一级须根数、改变根系构型来适应土壤干旱[26]。蚬木幼苗在土壤干旱条件下,也具有通过提高根冠比以增加水分吸收的适应策略。W水平蚬木幼苗生物量显著高于N水平,表明在岩溶水层有水条件下,植株可通过根系利用这部分水分以保证生长所需,生物量积累较多。N水平根冠比及根生物量比高于W水平,差异不显著;W水平茎生物量比显著高于N水平。研究结果说明岩溶水层有无水分在一定程度上影响着蚬木幼苗的生物量分配,但其影响与土壤层水分条件密切相关。
3.3 干旱胁迫对蚬木幼苗形态特征的影响
干旱胁迫对植物表型可塑性的影响已有一些研究,发现植物可通过调整形态来适应土壤水分条件的变化[27-28]。在本研究中,岩溶水分和土壤水分显著影响着蚬木幼苗的地径、株高、根长及叶面积等。其中,根长和叶面积主要受岩溶水分所影响,因素间交互作用显著影响着苗木地径和叶面积。表层土壤干旱抑制了蚬木幼苗的形态发育,岩溶水层有水条件下苗木形态特征值基本显著高于无水处理组。如W处理组蚬木幼苗的平均根长为(87.38±19.21)cm,显著高于N处理,意味着该组苗木根系可下扎至岩溶水层吸收水分。土壤干旱胁迫下可抑制植物根系生长[29-30],但在模拟喀斯特生境岩溶水层有水情况下,植株根系可扩散分布至岩溶水层[16]。这是由于岩溶水层的水分诱导了根系向下生长,当表层土壤水分干旱时,岩溶裂隙或岩溶水层的水分可保证植物生长需求。蚬木幼苗叶面积受土壤干旱所抑制,岩溶水层有水条件下的叶面积显著高于无水处理组。表明在表层土壤干旱下,蚬木幼苗可通过降低叶面积、减少水分蒸腾以适应干旱生境。以往的研究也证实了植物在干旱生境下的这种自我调节策略[31-33]。而在岩溶水层有水情况下,表层土壤干旱时苗木可从岩溶水层吸收水分补充生长所需,不必通过减小叶面积来适应土壤水分亏缺,植株叶面积受影响相对较小。此外,因素间交互作用对叶面积的影响达极显著水平,表明岩溶水分对叶面积的影响,与表层土壤水分条件密切相关。
综上所述,干旱胁迫下蚬木幼苗生长受土壤水分条件影响较大,在土壤水分严重亏缺时,蚬木幼苗表现为生长下降、生物量分配和形态发育发生改变等适应策略,岩溶层水分条件对苗木的影响与表层土壤水分条件密切相关。
[1] 赵中秋,蔡运龙,白中科,等.典型喀斯特地区不同土地利用类型土壤水分性能对植物生长及其生态特征的影响[J].水土保持研究,2007,14(6):38-40,37.
[2] 魏媛,喻理飞.西南喀斯特地区构树苗木对土壤干旱胁迫的生理响应[J].水土保持研究,2010,17(2):164-167.
[3] 刘锦春,钟章成,何跃军.干旱胁迫及复水对喀斯特地区柏木幼苗活性氧清除系统的影响[J].应用生态学报,2011,22(11):2836-2840.
[4] 王丁,姚健,杨雪,等.干旱胁迫条件下6种喀斯特主要造林树种苗木叶片水势及吸水潜能变化[J].生态学报,2011,31(8):2216-2226.
[5] BINH L V, THINH N V, KÖPP R, et al. Responses of native tree species to soil water stress in a tropical forest on limestone, vietnam[J]. Open Journal of Forestry,2015,5(7):711-722.
[6] 洪文君,徐瑞晶,魏依娜,等.干旱胁迫对圆叶乌桕和毛果巴豆幼苗生长,形态和生理特性的影响[J].华南农业大学学报,2015,36(1):79-84.
[7] 刘长成,刘玉国,郭柯.四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性[J].植物生态学报,2011,35(10):1070-1082.
[8] 吴显芝.模拟干旱胁迫对喀斯特森林喜钙树种伞花木生理特征的影响[J].中国水土保持,2012(5):33-35.
[9] 王丁,姚健,薛建辉.土壤干旱胁迫对樟树(Cinnamomumcamphora(L.) Presl)苗木水力结构特征的影响[J].生态学报,2009,29(5):2725-2731.
[10] 潘学军,张文娥,杨秀永,等.喀斯特山区野生葡萄实生苗叶片解剖结构与抗旱性的关系[J].贵州农业科学,2010,38(9):176-178.
[11] 张显强,王世杰,孙敏.干旱和复水对喀斯特石生反叶扭口藓(BarbulafallaxHedw.)叶绿素荧光特性的影响:以贵阳市花溪区附近严重石漠化区域为例[J].中国岩溶,2014,33(1):77-81.
[12] CAO J H, YUAN D X, ZHANG C, et al. Karst ecosystem of Guangxi Zhuang Autonomous Region constrained by geological setting: relationship between carbonate rock exposure and vegetation coverage[J]. Int Confr on Karst Hydrogeology and Ecosystems, USA,2007:211-218.
[13] NIE Y P, CHEN H S, WANG K L, et al. Seasonal water use patterns of woody species growing on the continuous dolostone outcrops and nearby thin soils in subtropical China[J]. Plant and Soil,2011,341(1/2):399-412.
[14] 丁亚丽,陈洪松,聂云鹏,等.基于稳定同位素的喀斯特坡地尾巨桉水分利用特征[J].应用生态学报,2016,27(9):2729-2736.
[15] 张中峰,尤业明,黄玉清,等.模拟岩溶水分供应分层的干旱胁迫对青冈栎光合特性和生长的影响[J].生态学杂志,2012,31(9):2197-2202.
[16] 张中峰,尤业明,黄玉清,等.模拟喀斯特生境条件下干旱胁迫对青冈栎苗木的影响[J].生态学报,2012,32(20):6318-6325.
[17] 吴朝学,严理,秦武明,等.木荷人工林营养元素分布格局[J].西北林学院学报,2017,32(1):54-61.
[18] 张珊珊,康洪梅,杨文忠,等.干旱胁迫下AMF对云南蓝果树幼苗生长和光合特征的影响[J].生态学报,2016,36(21):6850-6862.
[19] 尹丽,胡庭兴,刘永安,等.干旱胁迫对不同施氮水平麻疯树幼苗光合特性及生长的影响[J].应用生态学报,2010,21(3):569-576.
[20] 孙景宽,李田,夏江宝,等.干旱胁迫对沙枣幼苗根茎叶生长及光合色素的影响[J].水土保持通报.2011,31(1):68-71.
[21] 王林龙,李清河,徐军,等.干旱胁迫对不同种源油蒿幼苗的生长和形态可塑性的影响[J].东北林业大学学报,2015,43(10):55-57.
[22] 李冬琴,曾鹏程,陈桂葵,等.干旱胁迫对3种豆科灌木生物量分配和生理特性的影响[J].中南林业科技大学学报,2016,36(1):33-39.
[23] 张希吏,王萍,石磊,等.干旱胁迫对沙芥幼苗根系形态及抗氧化酶活性的影响[J].干旱地区农业研究,2016,34(3):160-164.
[24] 刘锦春,钟章成.水分胁迫和复水对石灰岩地区柏木幼苗根系生长的影响[J].生态学报,2009,29(12):6439-6445.
[25] 杨玉珍,贾遂民,彭方仁.干旱胁迫对不同种源香椿苗木生长的影响[J].林业工程学报,2009,23(6):52-55.
[26] 金静,钟章成,刘锦春,等.石灰岩地区土壤水分对木豆表型可塑性的影响[J].西南大学学报(自然科学版),2005,27(1):89-92.
[27] 何军,赵聪蛟,清华,等.土壤水分条件对克隆植物互花米草表型可塑性的影响[J].生态学报,2009,29(7):3518-3524.
[28] 徐军,陈海玲,李清河,等.土壤水分含量对白刺幼苗表型可塑性生长的影响[J].西北林学院学报,2017,32(2):101-105.
[29] 李芳兰,包维楷,吴宁.白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J].生态学报,2009,29(10):5406-5416.
[30] 吴敏,张文辉,周建云,等.干旱胁迫对栓皮栎幼苗细根的生长与生理生化指标的影响[J].生态学报,2014,34(15):4223-4233.
[31] 蔡海霞,吴福忠,杨万勤.干旱胁迫对高山柳和沙棘幼苗光合生理特征的影响[J].生态学报,2011,31(9):2430-2436.
[32] 井大炜,邢尚军,马海林,等.I-107欧美杨对不同强度干旱胁迫的形态与生理响应[J].东北林业大学学报,2014,42(1):10-13.
[33] ZOKAEEKHOSROSHAHI M, ESNAASHARI M, ERSHADI A, et al. Morphological changes in response to drought stress in cultivated and wild almond species[J]. International Journal of Horticultural Science & Technology,2014,1(1):79-92.
EffectofDroughtStressonExcentrodendronhsienmuSeedlingsunderSimulatingKarstEnvironment
Ou Zhiyang, Pang Shilong, Meng Fang, Tan Zhangqiang, Zheng Wei, Cao Yanyun, Shen Wenhui
(Guangxi Forestry Research Institute, Nanning 530002, P. R.China) Journal of Northeast Forestry University,2017,45(12):16-21.
Drought stress;Excentrodendronhsienmu; Growth characteristics; Biomass allocation; Leaf area
1)广西自然科学基金项目(2014GXNSFAA118132);广西科技计划项目(桂科AB16380300);广西林业科技项目(桂林科字[2014]02号) ;广西优良用材林资源培育重点实验室自主课题(15-A-02-02)。
欧芷阳,女,1975年10月生,广西壮族自治区林业科学研究院,高级工程师。E-mail:ozhiyang@126.com。
2017年5月4日。
潘 华。
Q945.78
To explore plant growth and morphological adaptive strategy to karst soil drought environment, we set up a water supply device combining with a simulated karst structure comprising soil, limestone, and epikarst layer. We studied the growth, biomass accumulation and allocation, and morphology of 2-aExcentrodendronhsienmuseedlings under the simulated karst environment. A two-factor design, having water or having no water in epikarst layer (epikarst layer water) and four levels of soil water condition, was performed to analyze the impacts onE.hsienmuseedlings. Both epikarst layer water and soil water significantly affected the growths of leaf, ground diameter, and height of seedlings. Values of net growth were significantly decreased with the increasing of soil drought stress, which in treatment with epikarst layer having water (W treatment) were greatly higher than those in treatment with epikarst layer having no water (N treatment). They also had very significant influence on the total biomass, leaf and stem biomass, and epikarst layer water affected the root biomass greatly. Total biomass and biomass of root, stem, and leaf were greatly decreased with the increasing of soil drought stress, which in W treatment were significantly higher than those in N treatment. Soil water markedly affected the root-shoot ratio, root and stem mass ratio, and epikarst layer water significantly impacted stem mass ratio. There was a significant epikarst layer water×soil water interaction on root-shoot ratio and root mass ratio, and very significant on stem mass ratio. The root-shoot ratio and root mass ratio in moderate and severe drought stress were greatly higher than those in control, while the stem mass ratio was markedly decreased with the increasing of drought stress. The stem mass ratio in W treatment was greatly higher than that in N treatment. Both of the two factors very significantly affected the ground diameter, height, root length, cumulative length of primary branch, and leaf area ofE.hsienmuseedlings. Epikarst layer water had great influence on the number of leaf and primary branch, while soil water had very significant effect. There was also a significant epikarst layer water×soil water interaction on ground diameter, and very significant on the leaf area. The values of morphological characters were basically decreased with the increasing of drought stress, which in W treatment were greatly higher than those in N treatment. The results indicated that the growth ofE.hsienmuseedlings was mainly affected by soil water condition, seedlings growth decline, biomass allocation and morphology changed under soil drought stress, and the influence of epikarst layer water onE.hsienmuseedlings was closely related to soil water.
猜你喜欢
中国公路(2022年15期)2022-09-25
湖南水利水电(2021年6期)2022-01-18
乡村地理(2019年2期)2019-11-16
中华建设(2019年2期)2019-08-01
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
山东工业技术(2016年15期)2016-12-01
高原山地气象研究(2016年2期)2016-11-10
文化月刊·下旬刊(2014年6期)2014-08-28
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
