
灰木莲二次开合开花动态与雌雄异熟特征
2018-01-16 09:52潘丽琴徐建民陈建全卢立华刘志龙
浙江农林大学学报 2018年1期
潘丽琴, 郝 建, 徐建民, 陈建全, 卢立华, 刘志龙
(1.中国林业科学研究院 热带林业实验中心,广西 凭祥532600;2.中国林业科学研究院 热带林业研究所,广东 广州 510520;3.西南林业大学,云南 昆明 650204;4.广西友谊关森林生态系统定位观测研究站,广西 凭祥 532600)
植物的开花方式和生殖同步性是影响植物繁殖适合度的重要因子,也是决定植物生殖成功的重要因素[1]。对大部分植物来说,开花通常是一次性绽放,之后通过花的闭合或花瓣的凋谢来终止开花进程,而有些植物的开花过程,开放和闭合是交替进行[2],称之为 “二次开合”。多种木兰科Magnoliaceae植物存在此种开花方式, 如广玉兰Magnolia grandiflora[3], 白玉花M.denudata[4], 厚叶木莲Manglietia pachyphylla[5], 大果木莲 M.grandis[6]和中缅木莲 M.hookeri[6]等。 该种开花方式受到诸如气温、 湿度和光照等气候因子的影响,其中气温条件在控制开花中起主导作用[7-8]。灰木莲Manglietia glanca原产越南及印度尼西亚,为木兰科木莲属Manglietia植物,具干形通直、抗逆性强、生长快、木材纹理细致及易加工的特性[9],是一种极具有发展潜力的速生用材及园林绿化树种。自1960年以来,灰木莲在中国西南和华南地区进行引种并规模化种植[10]。目前,有关灰木莲的研究主要集中于栽培繁育技术[11]、早期生长适应性[12]、材性[13]以及生态效应等方面。在引种地灰木莲开花后自然结实率极低[11,14],而针对该现象的研究报道较少[14-15]。为了探究灰木莲开花及雌雄异熟特征对其生殖成功的影响,笔者着重研究灰木莲的花部结构、不同花期的二次开合开花进程、花粉活力和柱头可授性,以探究灰木莲二次开合开花及雌雄异熟特征对其生殖的影响,也为深入了解木莲属的系统进化或演化过程提供资料。
1 研究地区概况和试验材料
试验点设在中国林业科学研究院热带林业实验中心,位于广西壮族自治区凭祥市,属南亚热带季风气候区,年均气温为21.5℃,≥10℃积温7 500℃,年降水量为1 220~1 400 mm,降水集中期为4月中旬至8月末,年均日照约为1 260 h。以伏波实验场、白云实验场和中心树木园栽培的灰木莲人工林为研究对象,各林分造林苗木均为1.5年生的实生苗,种源来自越南尚河,无病虫侵害,健康状况良好,基本情况见表1。观测区域花期内气温因子基本情况见表2。

表1 观测林分基本情况Table 1 General information of the observation stands
2 研究方法
2.1 林分水平物候的观测
2014-2016年连续3 a对灰木莲林分进行定点、定时观察并记录其开花物候。林分水平1%~10%的个体开花时视为始花期,10%~50%的个体开花时为过渡期,50%~95%的个体达到开花高峰时为盛花期,95%植株开花结束时为终花期。

表2 观测区域环境气温基本情况Table 2 General information of the atmosphere temperature in observation region
2.2 花部结构及开花动态的观测
在伏波实验场、白云实验场和中心树木园内试验点各选3株标准株,从青蕾期开始观察其开花动态,各个开花阶段分别标记10朵花,观察开花进程中的花部、柱头和花粉的形态变化特征,用佳能(Cannon)数码相机拍照,花部解剖结构在奥林帕斯(Olympus)体视显微镜成像系统下观察并拍照。
2.3 花粉活力及柱头可授性检测
在标准株上的上中下3个冠层各选取3个标准枝,各个标准枝于青蕾期选取15个花蕾。于始花期、过渡期和盛花期,分别选取3朵未散粉的花朵带回室内,人工挑出雄蕊,收集花粉,用于测定不同花期阶段花粉活力;于始花期、过渡期和盛花期,分别标记5朵青蕾,于露白(T1),初次展花(T2),闭合(T3),完全展花(T4)和衰败期(T5)等5个阶段分别采集花粉用于测定单花不同开花阶段花粉活力,同时取柱头样品用于柱头可授性检测。
2.3.1 花粉活力检测 根据正交设计的花粉离体萌发试验结果,采用最佳萌发效果的培养液对各开花阶段花粉进行离体萌发培养。萌发条件为:质量浓度为60.00 g·L-1蔗糖+0.50 g·L-1硼酸+0.08 g·L-1氯化钙,pH值为6,温度25℃。花粉萌发6 h后,把培养皿放置在倒置显微镜观察并拍照,选择5个视野统计花粉的萌发率。
2.3.2 对柱头可授性检测 采用联苯胺-过氧化氢法[16]检测,联苯胺-过氧化氢反应液为V(1%联苯胺溶液)∶V(3%过氧化氢溶液)∶V(水)=4∶11∶22的混合液。分别采各开花阶段花朵9朵,切除花被片,将雌蕊群浸入含有联苯胺-过氧化氢反应液的10 mL透明样品瓶中。若柱头具有可授性,则反应液呈现蓝色,并伴有大量气泡,反之则无。根据雌蕊群柱头所冒出气泡数量和染为蓝色的柱头数量判断其可授性。将柱头可授性分为5个等级[17]: “-”为不具有可授性,气泡出现慢且少,颜色无变化; “+/-”极少部分柱头有可授性, “+”为具有较弱的可授性,表示少部分柱头有可授性,大部分没有,气泡出现慢且少,着色慢、颜色浅; “++”为具有较强的可授性,气泡出现快且多,着色快、颜色浅; “+++”为具有很强的可授性,气泡出现快且多,着色快、颜色深。
2.4 不同花期阶段灰木莲人工授粉
在标准株的上中下3个冠层各选取5个标准枝,在青蕾期每个标准枝选取30个花蕾。在始花期、过渡期和盛花期,分别标记10朵花,在初次展花阶段进行自花、同株异花和异株异花授粉;于盛花期按各单花露白阶段、初次展花阶段、(长)闭合阶段和完全展花阶段进行异株异花授粉,各个标准枝各阶段分别授粉6朵花。授粉后2个月统计结实率。单个果实明显膨大,子房数量占其果实总子房数量50%以上为授粉成功。
3 结果与分析
3.1 灰木莲花部结构与群体开花物候
灰木莲为两性单花,展花直径15~25 cm,花被片乳白色,厚肉质,汤勺形,长6~10 cm,宽5~7 cm,共6~8片;多心皮的雌蕊群在上,雄蕊群聚合于雌蕊群下方基部的花托上。雌蕊群由45~85枚单生心皮组成,心皮绿色,短纺锤形,长0.4~0.6 cm,紧密组合呈锥形,每心皮内含双排胚珠8~12颗,成熟柱头向外微卷曲;雄蕊群着生于雌蕊群的下部,高度仅达到雌蕊群的下缘,单个雄蕊长0.7~1.1 cm,宽0.8~1.5 mm,乳白色,2对细长的花粉囊平行分布于花药的2侧。
在中心树木园(海拔为240 m),灰木莲花期主要从3月上旬持续至5月上旬,少数从1月下旬开始,仅个别单株出现1 a中2次花期,第2次花期从10月中旬持续到12月上旬,花量极少;第1次花期3月上、中旬为始花期,3月下旬到4月中旬为盛花期,4月下旬到5月上旬为终花期。灰木莲在高海拔(600 m左右)比低海拔(240 m)的花期推迟15~20 d。在花期内,环境温度分别为始花期的10~20℃,过渡期18~25℃和盛花期的21~30℃,不同气温下灰木莲表现出不同的开花方式。
3.2 灰木莲不同气温单花二次开合开花时序特征
灰木莲开花过程存在二次开合的开花现象(图1)。先后经历:①露白(T1):内部白色花被片开始松动,绿色外苞片露出花被片,形成白绿相间的花蕾。②初次展花(T2):花被片初次展开,外苞片和内层花被片同时展开30°~50°。③闭合阶段(T3):最外层花苞片沿花柄向下展开180°,白色花瓣向相反方向收拢至完全闭合,多发生于18:00-24:00,根据气温高低存在2种闭合方式,高温时闭合时间约为10~12 h,气温较低时闭合时间约为48~72 h。④完全展花(T4):花被片再次展开约90°,不再闭合,雄蕊群松散脱落,花粉散出,多发生于长闭合形成白蕾后第2天17:00-20:00;在盛花期,完全展花前会出现一次短闭合阶段。⑤衰败阶段(T5):完全展花的第2天,花被片逐渐褐化、萎蔫、凋落,至完全凋落需3~7 d。灰木莲在不同花期阶段的开花方式特征见表3。

图1 盛花期灰木莲二次开合开花动态Figure 1 Floral dynamic of Manglietia glanca at full-bloom stage
3.3 灰木莲不同花期花粉活力和柱头可授性特征
由表4可知:灰木莲花粉活力随时间推移而下降,盛花期时花粉活力最高,平均活力为72.13%,其次为过渡期63.15%,始花期最低仅为43.28%。盛花期时,因环境温度最高(21~30℃),散粉后花粉活力急剧下降,散粉后18 h时,花粉已完全失活。而在环境温度较低的过渡期(18~25℃)及始花期(10~20℃),散粉后花粉活力维持时间相对较长,分别达到24 h,36 h。
灰木莲在始花期的完全展花、过渡期的初次展花及盛花期的白蕾期柱头黏液分泌量最多;单花完全展花时,始花期与过渡期的柱头均有少量的黏液,而盛花期的柱头黏液已变干。经过联苯胺-过氧化氢检测,柱头黏液分泌量较多时,柱头颜色多呈乳白色和淡黄色,柱头可授性较强,柱头在纯白色和褐(黑)色时,黏液较少,可授性均较弱(表5)。柱头在过渡期时的初次开花和完全展花时、盛花期时的白蕾期及初次展花时、始花期时的完全展花时,均具有较强的可授性,其余时期柱头可授性较弱或基本失活。

表3 灰木莲不同气温单花二次开合开花时序特征Table 3 The flowering temporal characteristics of Manglietia glanca
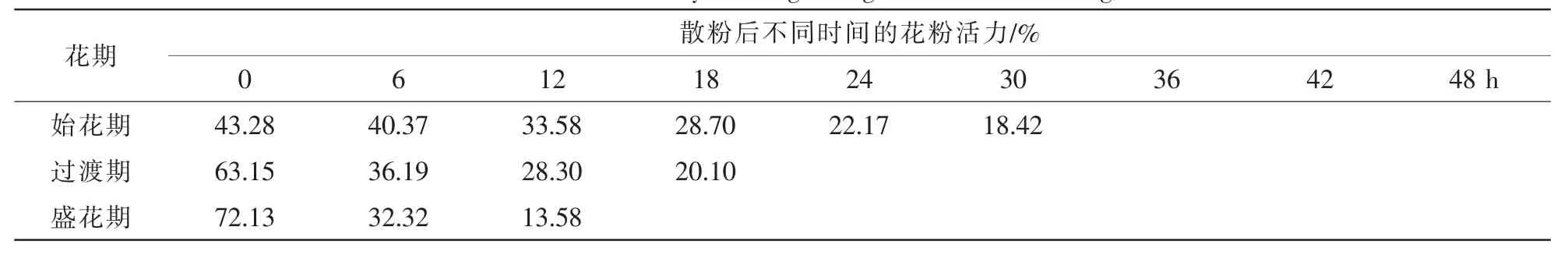
表4 灰木莲不同花期阶段的花粉活力Table 4 Pollen viability of Manglietia glanca at different stages
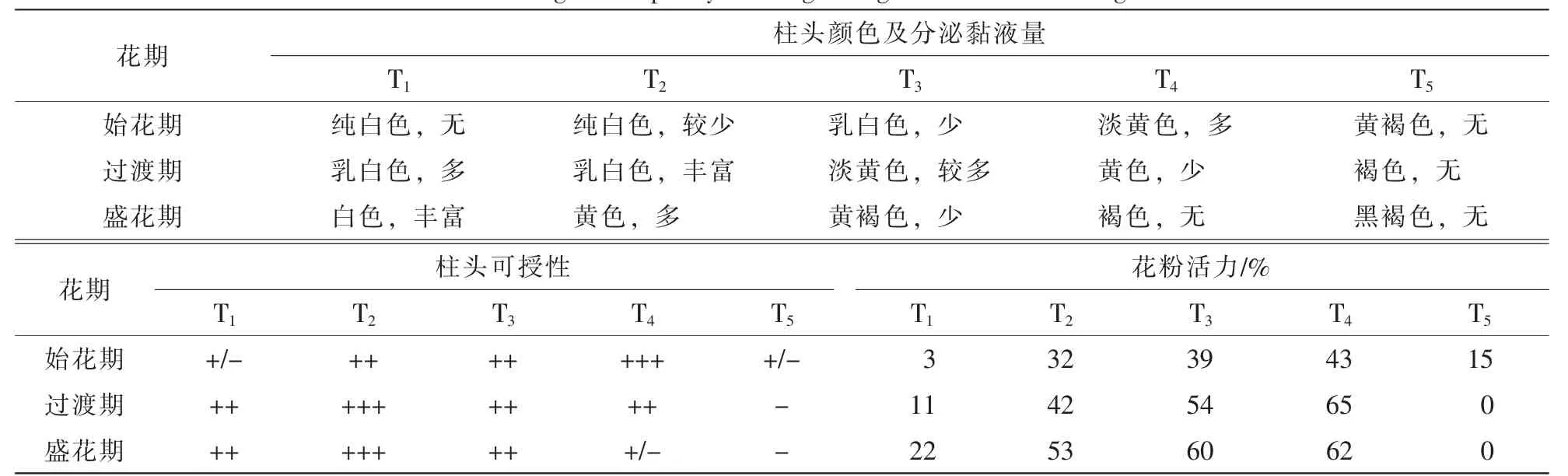
表5 灰木莲不同花期阶段的柱头可授性Table 5 Stigma receptivity of Manglietia glanca at different stages
灰木莲雌性先于雄性表达的特征稳定,交配类型为雌先型。不同气温条件下,雌雄性成熟的间隔时间有所差异,气温越高雌性表达发生时间越早,雌雄异熟的间隔时间越短;雄性表达在初次展花时均已表达,至完全展花时,花粉活力最强。盛花期露白阶段时柱头可授性最高,雌性表达在青蕾阶段就已开始,而雄性特征还未表达;当完全展花花粉完全散开成熟时,雌性特征最弱,雌雄异熟间隔时间约为2 d。过渡期初次展花时柱头可授性最强,但花粉未完全成熟,完全展花时花粉活力最高,柱头仍具可授性,雌雄异熟的间隔时间为24~36 h。始花期闭合阶段时柱头可授性最强,但花粉活力较弱,完全展花时花粉活力和柱头可授性均较高,雌雄异熟的间隔时间为2~3 d。
3.4 灰木莲花期不同阶段单株二次开合花过程可授性重叠期
根据图2:灰木莲在盛花期开花时,前一轮花完全展花的散粉期和后一轮花初次展花的可授期重叠。在这个重叠期里,前一轮完全展花,花粉自然散开,花粉活力也较强,第2轮花初次展时柱头可授性较强,但花粉未散开,为异花授粉提供极好的机会;完全展花花粉散出时,同一花朵柱头可授性极低,发生自花授粉的机率极低;在过渡期开花时,异花和同花授粉均可能发生,前一轮花完全展花的散粉期和后一轮花初次展花的散粉期重叠期花粉活力和柱头可授性均较强,为异花授粉提供可能;此时同一花朵花粉活性和柱头可授性均较强,自花授粉也可发生;始花期开花时,第1轮完全展花的散粉期与第2轮初次展花的可授期没有重叠期,与第3轮初次展花时可授期重叠,可能发生异花授粉;而第1轮完全展花时同一花朵柱头可授性与花粉活力均很强,有利于自花授粉的发生。

图2 灰木莲每轮开花在不同时间的柱头可授性与花粉活力Figure 2 Stigma receptivity and pollen viability of each round flowering of Manglietia glaucaat different times
图3显示:灰木莲在初次展花阶段进行自花、同株异花和异株异花方式均能成功授粉,异株异花授粉成功率明显高于自花和同株异花授粉,异株异花在不同花期人工授粉平均成功率为57.67%,同株异花授粉平均成功率为45.00%,自花授粉的平均成功率为5.67%,表明灰木莲的繁育习性是以异交为主,但存在自交亲和的现象。始花期、过渡期和盛花期人工授粉平均成功率分别为26.00%,39.33%和43.00%,表明气温对授粉成功率有所影响,相对于过渡期(18~25℃)与盛花期(21~30℃),在始花期(10~20℃)时授粉率最低,说明低温不利于灰木莲的受精及胚胎发育。灰木莲在开花过程中的可授性是不断变化的,在初次展花前1~2 d,即露白期柱头已具有可授性,完全展花后,柱头可授性急剧下降,约1~6 h后完全失活。在不同开花阶段通过异株异花方式进行人工授粉的授粉成功率如图4所示,闭合阶段与完全展花阶段的平均授粉成功率最高,分别为52.33%,49.67%,初次展花次之,为48.00%,露白阶段的平均授粉成功率最低,为21.33%,与上述各阶段柱头可授性检测结果基本一致。
3.5 不同花期阶段灰木莲人工授粉的成功率
4 结论与讨论
4.1 灰木莲二次开合开花的生物学意义
灰木莲的开花特征与多种木兰科树种相似[4,18]。在花期不同阶段,灰木莲的开花方式受温度影响而产生开合交替不同的开花模式,存在二次开合现象。有关开花过程中的闭合现象有几个解释:①保护花朵不被昆虫吃食[19];②给传粉昆虫提供庇护场所,特别是对于一些直立闭合的花朵,能增加携带着花粉的甲虫类的传粉机率[20];③对于在白天发生闭合的花朵,则是为了保护花粉不被散落或使花粉保持一定的湿度以维持更好的活力[21]。这种开花方式还可以保护花粉不易被风吹走,利于昆虫传粉[6],利于花内形成小温室促进散落在柱头的发粉萌发[3]。
灰木莲在单花内表现为雌雄异熟,雌蕊先熟,雌蕊位于雄蕊上部,在时间和空间上均不利于自花传粉。单花水平上,花朵在初次展花时雌雄性征均已表达,但花粉仍未释放,待完全展花,花粉释放时,雌性征减弱或完全失活,避免了自花传粉发生的可能;在植株水平上,灰木莲表现为异步雌雄异熟且同步集中式开花,各轮花二次开合交替出现,前一轮花的完全展花也即雄性表达时,正好为下一轮花初次展花也即雌性表达时,此时2轮开花雌雄性征均较强,雌雄表达重叠可达1~6 h。这种重叠和集中式开花可使植株开花时有较多的花粉资源进行同株异花和异株异花授粉,保证其在有限的可授期内授粉成功。这种开花方式下,传粉者的数量是否充足、环境条件是否允许传粉者在花期内进行有效的传粉活动成为灰木莲在引种地生殖成功的关键因素。

图3 不同授粉方式对灰木莲授粉成功率影响Figure 3 Pollination success rate of Manglietia glauca withdifferent pollination ways
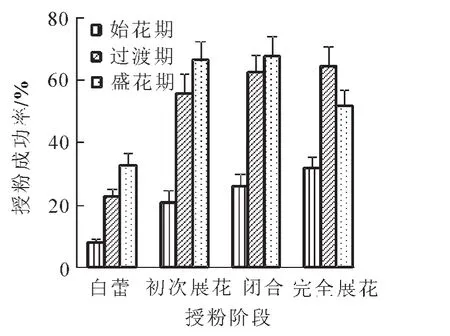
图4 不同授粉阶段对灰木莲授粉成功率的影响Figure 4 Pollination success rate of M.glauca in different stages
4.2 雌雄异熟与二次开合的关系
花被片展开的动力是由于花部内外温度差异导致花被片两面细胞扩张程度不均产生的[2]。木兰属植物在开花进程中存在开花生热效应,使花器官内部温度明显高于环境温度而促使开花[4]。木兰属存在雌雄异熟的特征,在雌蕊成熟时促使其初次展花,之后花朵闭合,雄蕊成熟时促使第2次完全展花[3]。气温较高时,灰木莲雌蕊成熟较快,短时间花内聚集较多的热量,当花内积温高于外部温度,发生第1次展花,之后花内温度迅速下降,外部温度高于花内温度,花瓣内部细胞发生可逆性收缩,花瓣闭合。闭合后雄蕊经过一段时间成熟,在花内部再次生热积温,花瓣完全展开,花瓣内面细胞拉升到最大限度,花瓣内部细胞不再发生可逆性收缩而使花朵闭合。气温较低时,第1次展花发生的时间较晚,2次展花间隔期也更长。
4.3 灰木莲开花特征与生殖适合度
植物的开花时间与生殖同步性可以在多方面影响其生殖成功与否[1]。灰木莲在广西凭祥地区的花期主要集中在3月上旬至5月下旬,个别单株1月下旬出现开花,偶尔出现 1 a中2次花期。第2次花期从10月中旬持续到12月上旬。灰木莲不同花期经历的气温不同,表现出不同的二次开合开花特征,不同开花阶段发生和持续的时间差异较大。在温度较高的过渡期和始花期,单花雌雄表达间隔期较短,单株上各轮花雌雄表达重叠时间也相应缩短,但开花量较多,传粉者数量增加,传粉效率更高。而在温度最低的始花期,开花量和传粉者均较少,但各开花阶段持续时间较长,各轮开花的雌雄性表达重叠的时间较长。这可能是为较少传粉者提供较长传粉时间的生殖适应性策略。
[1]O’NEIL P.Selection on flowering time:an adaptive fitness surface for nonexistent character combinations [J].Ecology,1999,80(3):806 - 820.
[2]van DOORN W G,van MEETEREN U.Flower opening and closure:a review [J].J Exp Bot,2003,54(389):1801 -1812.
[3]孟希,王若涵,谢磊,等.广玉兰开花动态与雌雄异熟机制的研究[J].北京林业大学学报,2011,33(4):63-69.MENG Xi,WANG Ruohan,XIE Lei,et al.Flowering dynamics and dichogamous mechanism in Magnolia grandiflora[J].J Beijing For Univ,2011,33(4):63 - 69.
[4]王若涵.木兰属生殖生物学研究及系统演化表征探析[D].北京:北京林业大学,2010.WANG Ruohan.A Study on Reproductive Biology in Magnolia and Its Systematic Characterization [D].Beijing:Beijing Forestry University,2010.
[5]杨晓丽.濒危植物厚叶木莲的保护生物学研究[D].北京:中国科学院大学,2013.YANG Xiaoli.Studies on Conservation Biology of Endangered Species Manglietia pachyphylla [D].Beijing:University of Chinese Academy of Sciences,2013.
[6]付玉嫔,陈少瑜,吴涛.濒危植物大果木莲与中缅木莲的花部特征及繁育系统比较[J].东北林业大学学报,2010, 38(4): 6 - 10.FU Yupin,CHEN Shaoyu,WU Tao.Comparison of floral traits and breeding system of endangered plant Manglietia grandis and non-endangered plant Manglietia hookeri[J].J Northeast For Univ,2010,38(4):6 - 10.
[7]BYNUM M R,SMITH W K.Floral movements in response to thunderstorms improve reproductive effort in the alpine species Gentianaalgida (Gentianaceae) [J].Am J Bot,2001,88(6):1088 - 1095.
[8]舒素芳,毛俊萱,蔡敏.白玉兰始花期与气象因子的关系分析[J].浙江农业学报,2013,25(2):248-251.SHU Sufang,MAO Junxuan,CAI Min.Analysis on relationships between first-flowering dates of Magnolia denudata and meteorological factors [J].Acta Agric Zhejiang,2013,25(2):248 - 251.
[9]刘玉壶.中国木兰[M].北京:科学技术出版社,2003:142-143.
[10]王克建,蔡子良.热带树种栽培技术[M].南宁:广西科学技术出版社,2008:71-72.
[11]乔梦吉.广西优良珍贵树种灰木莲的组织培养[J].南方农业科学,2013,44(6):989-993.QIAO Mengji.Tissue culture of rare species Manglietia glauca in Guangxi[J].J Southern Agric,2013,44(6):989 -993.
[12]卢立华,何日明,农瑞红,等.坡位对灰木莲生长的影响[J].林业科学研究,2012,25(6):789-794.LU Lihua,HE Riming,NONG Ruihong,et al.Effect of slope position on the growth of Manglietia glauca [J].For Res,2012,25(6):789 - 794.
[13]陈松武.人工林灰木莲木材材性及其薄木贴面工艺研究[D].南宁:广西大学,2011.CHEN Songwu.Study on Properties of Wood from Manglietia glauca Plantation and Technics of Veneer Faced the Wood [D].Nanning: Guangxi University,2011.
[14]招礼军,韦善华,朱栗琼,等.灰木莲的开花特性及繁育系统的研究[J].西部林业科学,2015(2):24-28.ZHAO Lijun,WEI Shanhua,ZHU Liqiong,et al.Flowering characteristics and breeding system of Manglietia glauca[J].J West China For Sci,2015(2):24 - 28.
[15]招礼军,龙永宁,朱栗琼,等.灰木莲花粉的萌发率和生活力[J].广西林业科学,2014,43(4):405-408.ZHAO Lijun,LONG Yongning,ZHU Liqiong,et al.Germination rate and viability of Manglietia glauca pollen [J].Guangxi For Sci,2014,43(4):405 - 408.
[16]刘林德,张萍,张丽,等.锦带花的花粉活力、柱头可授性及传粉者的观察[J].西北植物学报,2004,24(8): 1431 - 1434.LIU Linde,ZHANG Ping,ZHANG Li,et al.Pollen viability,stigma receptivity and pollinators of Weigela florida [J].Acta Bot Boreal-Occident Sin,2004,24(8):1431 - 1434.
[17]张瑞,李晖,彭方仁,等.薄壳山核桃开花特征与可授性研究[J].南京林业大学学报(自然科学版),2014, 38(3): 50 - 54.ZHANG Rui,LI Hui,PENG Fangren,et al.Flowering morphological characteristics,pollen viability and stigma receptivity of Carya illinoinensis [J].J Nanjing For Univ Nat Sci Ed,2014,38(3):50 - 54.
[18]PAN Yuezhi,LIANG Hanxing,GONG Xun.Studies on reproductive biology and endangerment mechanism of the endangered plant Manglietiaaromatica [J].Acta Bot Sin,2003,45(3):311 - 316.
[19]STIRTON C H.Nocturnal petal movements in the Asteraceae [J].Bothalia,1983,14(3/4):1003 - 1006.
[20]GOLDBLATT P,BERNHARDT P,MANNING J C.Pollination of petaloid geophytes by monkey beetles (Scarabaeidae:Rutelinae:Hopliini) in southern Africa [J].Ann Missouri Bot Gard,1998,85(2):215 - 230.
[21]DAFNI A.Autumnal and winter pollination adaptations under Mediterranean conditions [J].Bocconea,1996,5(1):171-181.
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
气象水文海洋仪器(2022年2期)2022-07-08
贵州林业科技(2022年2期)2022-06-22
农产品市场周刊(2022年4期)2022-05-21
农产品市场周刊(2022年4期)2022-03-23
新疆农垦科技(2022年1期)2022-03-10
华东科技(2021年9期)2021-09-23
新青年(2019年8期)2019-08-22
食品与健康(2018年8期)2018-08-23
创作评谭(2014年3期)2014-05-12
