
晚松人工林株间株内球果及种子形态变异研究
2018-01-25 01:44胡冬南刘苑秋
中南林业科技大学学报 2018年1期
赖 猛,胡冬南,易 敏,刘苑秋
(江西农业大学 园林与艺术学院,江西 南昌 330045)
晚松Pinus serotina原产美国东南部沿海平原、丘陵和低山区,具有耐干旱、耐贫瘠、生长适应性强、能在不良环境下稳定生长、生长快、热值高、萌生能力强、轮伐期短等特性。20世纪60年代,晚松作为薪炭林、用材林树种开始引进我国种植,引种区域包括山东、浙江、江苏、湖北、江西等省份,目前均表现出良好的生长特性。前期研究方面,主要集中在晚松生长规律和生态适应性[1-3]、无性繁殖[4-5]、组织培养[6-7]、抗性生理[8-9]以及生物质能源利用[10-11]。大部分松属树种的球果成熟期多为2 a,少有3 a,球果成熟后会自然开裂并散播种子。和一般松属树种相比,晚松的球果具有多年宿存的特性,球果成熟后少见自然开裂并散播种子,林下更新少。基于晚松球果多年宿存特性而进行的株间株内球果及种子形态变异研究,国内外未见相关报道。
植物的表型性状是环境与遗传因素共同作用的结果,表型多样性是遗传多样性研究的重要内容,亦是遗传多样性与环境多样性的综合体现[12-14]。研究林木群体的表型变异,不仅能够揭示遗传因素和环境因素在物种的生态适应和分化过程中扮演的角色,同时也能为该物种林木遗传改良和育种提供理论依据[15-16]。球果和种子是物种繁殖系统的重要特征,受较强的遗传控制,具有区分和比较的意义。大量研究表明,植物球果及种子形态性状在种群间、种群内、甚至个体间具有很大差异。种子形态性状对植物的繁殖更新和种群扩散具有重要意义,特别是种子单粒质量,其表征了种子内营养物质的多少,对种子萌发、幼苗定居及存活具有很大影响。球果性状是植物有性生殖效率的体现,其形态大小直接影响种子形态的大小、数量以及单粒质量[17-22]。本研究通过测定晚松人工林种群内株间、株内球果及种子形态性状,分析晚松球果及种子形态性状在株间、株内表型变异模式及变异规律,探究晚松球果形态性状与种子形态性状间的相互关系,通过表型变异分析和相关分析,找出晚松株间、株内差异大的表型特征,为晚松后期的良种选育工作提供理论基础。
1 材料和方法
1.1 试验地概况
试验地位于江西峡江林木良种场,东经115°33′,北纬 27°62′,属中亚热带季风性湿润气候,年平均气温为17.7 ℃,年平均降水量为1 557.9 mm,年蒸发量为1 470.4 mm,年均日照时数为1 872.3 h,土壤类型为红壤。晚松人工林于1983年营造,1982年从美国引种,造林面积约0.67 hm2,株行距为4 m×4 m。
1.2 试验材料
2014年12月在江西峡江林木良种场晚松人工林中采集果实,从林分中挑选5株球果宿存年龄较为完整的样木进行球果收集,各样木之间相距25 m以上,每株样木基本概况如表1所示。每株样木从枝条年龄来推算球果结实年代和宿存延迟开放时间,其中2、3年生球果采集30个,4年生球果采集20个,5年生球果采集10个,每株标准木共采集90个球果。随着球果宿存年龄的增加,枝条上球果数也越来越少,因此在采样过程中,球果样品数随着宿存年龄的增加而减少。对球果进行烘干脱粒处理并取得种子,单株内每年龄随机抽取30粒种子进行种子形态测定,每单株共120粒种子。
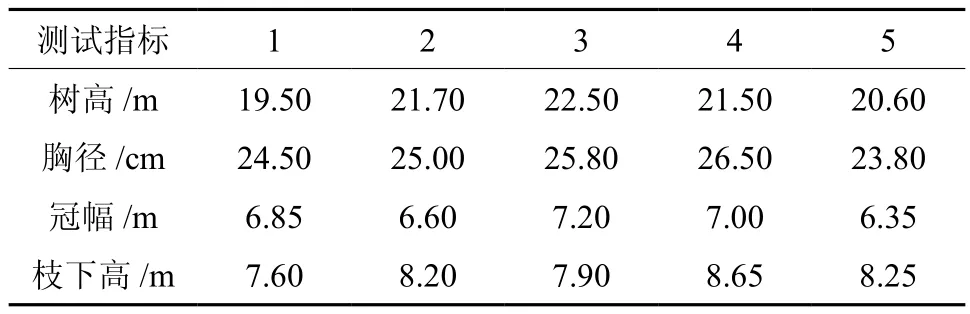
表1 采种母树的基本概况Table 1 Basic information of the mother trees for seed collection
1.3 球果及种子形态性状的测定
用游标卡尺分别测定球果的长和宽(果实的最长和最宽处,均在种鳞未开裂前测量),用千分之一天平测定球果质量;用果长×果宽作为果实大小指标,用果长/果宽表示果实的形状指数。用游标卡尺测量种子长、种子宽及种子厚(长度、宽度、厚度的指标均选在种子的最长、最宽、最厚处测量),用万分之一天平测定种子单粒质量(带翅);用种长×种宽×种厚表示种子的大小指标,用种长/种宽、种长/种厚及种宽/种厚表示种子的形状指数。
1.4 数据处理与分析
利用Excel对数据进行初步处理;球果及种子形状分别进行方差分析及相关性分析,并进行显著性检验。
建立种子单粒质量的多元回归模型,由各变量标准化回归系数的绝对值占标准化回归系数绝对值总和的百分比计算各自贡献率,研究各性状对种子单粒质量贡献的大小。模型为:

式中:因变量为Y;n个自变量分别为X1,X2,…,Xn;B1,B1,B2,…,Bn是模型的参数。
采用回归分析方法建立种子单粒质量与相应性状的回归方程,模型为:

式中:y为自变量;x为因变量;a为常数项;b为回归系数。
上述各项分析均采用SPSS软件进行处理,其中球果和种子性状的表型变异分析采用单个数据,相关分析、多元回归分析和线性回归分析采用单株内不同年龄均值数据进行处理。
2 结果与分析
2.1 晚松株间株内球果及种子形态变异
2.1.1 晚松株间球果及种子形态变异
晚松株间球果及种子形态性状方差分析结果见表2。由表2可知,晚松球果及种子性状的株间差异均达极显著水平。晚松不同母株间球果及种子形态性状表型变异分析的统计参数见表3。

表2 晚松株间球果及种子形态性状方差分析Table 2 Analysis of variance for the morphological traits of cone and seed among Pinus rigida trees

表3 晚松株间球果及种子形态性状表型变异分析Table 3 Phenotypic variation analysis of morphological traits of cone and seed among Pinus rigida trees
从各性状的变异系数来看,果质量和种子单粒质量变异系数最大,分别达到了25.76%和36.51%;其次为果长×果宽和种长×种宽×种厚,变异系数分别为21.08%和24.00%。
2.1.2 晚松株内球果及种子形态变异
晚松球果及种子形态性状株内表型变异分析的统计参数见表4。各母株中,母株4的表现最优,大部分性状在各母株中均排名第一;除母株4外,其余母株晚松球果和种子形态性状的分化较为明显,无明显的分布规律。从变异系数来看,与晚松株间球果及种子形态性状分析结果一致,果质量和种子单粒质量变异系数最大,其分布范围分别达到了17.64%~22.76%和20.82%~42.25%,其次为果长×果宽和种长×种宽×种厚,变异系数的分布范围分别为13.46%~18.08%和16.52%~21.82%。
2.2 晚松球果和种子性状的相关性分析
表5列出了晚松球果和种子性状的相关系数。从表5中可看出,球果性状中,果长、果宽、果质量、果长×果宽之间的相关性均达极显著水平,表现出极强的正向相关关系(0.716~0.952);表示种子形状指数的种长/种宽、种长/种厚、种宽/种厚与种长、种宽、种厚、种长×种宽×种厚的相关程度不一,总体来看,种长、种厚与表示种子形状指数各性状间的相互关系更紧密些,但种厚与种子形状指数性状为负相关;种子单粒质量除与种子形状指数性状相关不显著外,与其余种子性状相关均达极显著水平,分布范围为0.629~0.835。
与球果性状测量相比,种子性状测量过程更为复杂,其所耗费的人力和物力也高于球果性状。如果已知球果性状与种子性状的相互关系,则可以利用较容易测定的球果性状对种子性状进行预测和间接选择,从而提高选择效率,降低种质改良的成本。由表5可知,表示种子形状指数的各性状与除果长/果宽外的大部分球果性状相关不显著,而球果性状中的果长、果宽、果质量、果长×果宽与种子性状中的种长、种宽、种厚、种长×种宽×种厚、种子单粒质量彼此间的正相关性可达到显著或极显著水平。
2.3 晚松球果及种子性状对种子单粒质量贡献率剖析
种子单粒质量作为衡量种子品质的主要指标,是一种重要的亲代特征指标,在种质遗传改良过程中起着重要作用。种子单粒质量越大,通常出苗率越高,苗期各项指标表现也越好。由球果和种子性状的相关分析结果可知,种子单粒质量与果长、果宽、果质量、果长×果宽、种长、种宽、种厚、种长×种宽×种厚的相关性均达极显著水平。为进一步了解上述性状对种子单粒质量的影响程度,将种子单粒质量与这8个性状进行多元回归分析,采用的模型如下:Y=B0+B1X1+B2X2+B3X3+B4X4+B5X5+B5X6+B7X7+B8X8。

表4 晚松株内球果及种子形态性状表型变异分析Table 4 Phenotypic variation analysis of morphological traits of cone and seed within Pinus rigida trees
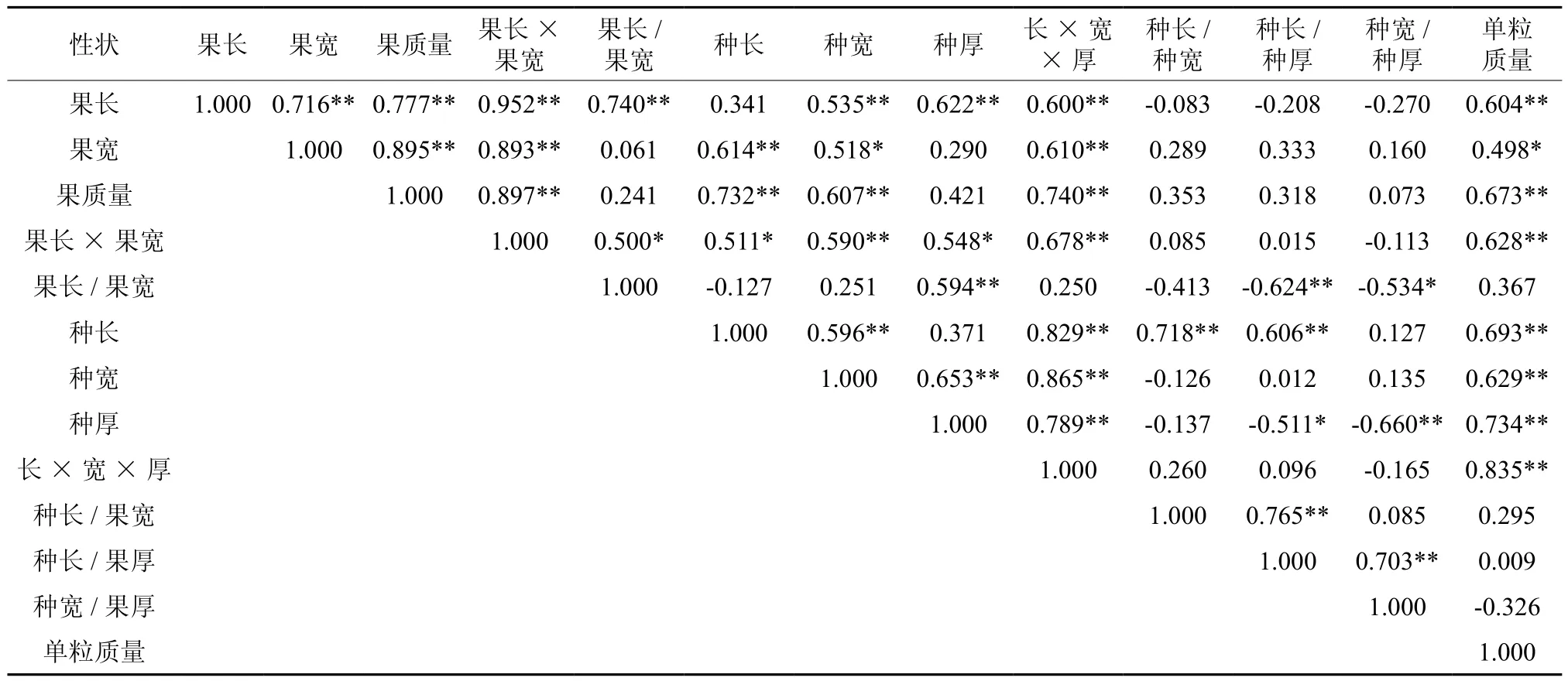
表5 晚松球果及种子形态性状相关性†Table 5 The correlations among morphological traits of cone and seed of Pinus rigida
多元回归分析结果见表6。由表6可知,种子单粒质量与果长、果宽、果质量、果长×果宽、种长、种宽、种厚、种长×种宽×种厚的多元回归方 程 为Y=–23.176 + 0.024X1– 0.030X2+ 0.196X3–2.760E-05X4+ 27.210X5–6.108X6+ 119.466X7–105.324X8。标准回归系数是消除了因变量Y和自变量X1,X2,…,X8所取单位的影响之后的回归系数,其绝对值的大小直接反映了X对Y的影响程度,由此可计算出各性状对种子单粒质量的贡献率。球果各性状中,果质量对种子单粒质量的贡献率最大(达25.10%),果长×果宽的贡献率最低(仅为0.26%),整个球果性状的贡献率为30.92%。种子各性状中,种长对种子单粒质量的贡献率最大,达28.43%,种厚略低(27.65%),种宽的贡献率最低(2.09%),整个种子性状的贡献率为69.08%。

表6 种子单粒质量与球果及种子性状的多元回归分析Table 6 The multiple regression analysis for single seed weight and corresponding cone and seed properties
2.4 晚松球果及种子性状和种子单粒质量线性回归模型的建立
对种子单粒质量贡献率排名前4位的性状别为果质量、种长、种厚及种长×种宽×种厚;建立相应的线性回归方程,各参数值和方程的决定系数如表7所示。结果表明,种子单粒质量与相对应球果及种子性状间均存在极显著的回归关系,说明各方程均具有较好的预测效果。种子单粒质量与果质量、种长、种厚及种长×种宽×种厚间方程的决定系数分别为0.618、0.529、0.428和0.666,其中种子单粒质量与果质量及种长×种宽×种厚(0.618和0.666)的决定系数要高于种长和种厚(0.529和0.428)。
3 结论与讨论
(1)晚松株间球果及种子形态性状方差分析结果表明,晚松球果及种子性状的株间差异达极显著水平,说明晚松球果及种子形态性状在母株间存在较大差异,各性状多形现象十分明显,受到较强的遗传控制。该结果与松科其它树种的研究结果相似。Beaulieu和Simon[23]对北美乔松Pinus strobus的球果及种子形态指标进行变异分析,发现北美乔松各性状的形态变异大部分存在于种群内的母树间,种群间的变异分量只占少量部分。Maley和Parker[24]对斑克松Pinus banksiana天然种群的球果、种子和针叶形态进行表型变异分析,发现各性状大部分的变异来自种群内部,种群间的差异只占1.6%~18.9%。然而,在王小平等[25]对白皮松Pinus bungeana种子及球果形态特征地理变异的研究中,发现白皮松同一种源内不同单株的种子和球果形态指标差异不大,个体之间的变异未达显著水平,其结果预示着如果白皮松种子和球果形态指标之间存在差异,则可能来自于群体差异而非个体差异。

表7 种子单粒质量与球果及种子相关性状的线性回归分析Table 7 The linear regression analysis of single seed weight and corresponding cone and seed properties
(2)表型多样性是遗传多样性研究的重要内容,是遗传因素和环境条件共同影响的结果[26-28]。晚松不同母株球果及种子性状均值差异明显,其中,母株4的表现最优,除了表示球果和种子形状指数的性状有所波动外,其余性状在各母株中均排名第一。变异系数可以表示性状值的变异程度,变异系数越大,则性状值变异程度越大。在晚松球果及种子的13个表型性状中,无论是株间变异亦或是株内变异,果质量和种子单粒质量变异系数均为最大,表明果质量及种子单粒质量性状内差异要高于其它性状,具有较大的选择潜力,该结果与张飞琳等[18]对太白山油松Pinus tabulaeformia的研究结果一致。大量研究[29-30]表明,种子质量分异对种子散播具有重要作用。本研究中,种子单粒质量的变异系数分布范围在所有性状中位居第一,这对晚松种子扩散、传播至林窗、增加幼苗幼树的成活率具有重要影响。
(3)探讨球果与种子表型性状之间的相关关系,对指导晚松选择育种和林木种质资源保存中样本策略的制定具有十分重要的意义。从相关分析结果来看,表示种子形状指数的各性状与大部分球果性状相关不显著,说明种子形状指数性状相对而言是比较独立的,可进行独立选择。而球果性状中的果长、果宽、果质量、果长×果宽与种子性状中的种长、种宽、种厚、种长×种宽×种厚、种子单粒质量彼此间相关紧密,说明晚松球果越大,果质量越重,相应的其种子也越大,种子单粒质量也越重,这与前人的研究结果有所不同。有研究认为,球果越大,种子质量越小,种子变小变轻,相应的种子籽粒变多[31-32]。但亦有研究与本文的研究结果一致。在齐立志等[33]对长白落叶松Larix olgensis雌雄花及球果、种子性状的变异研究中,发现球果大的长白落叶松,其种鳞数多,种子大,千粒重也大;王润辉等[34]对江南油杉Keteleeria cyclolepis球果及种子性状进行相关分析,发现球果与种子各性状间均为紧密的正相关关系,呈现出球果性状越大其种子性状也越大的趋势。除上述两种研究结果之外,也有学者认为,种子单粒质量与球果质量并不是简单的线性关系,而是先随着球果的增加而增加,当达到峰值后,又随着球果质量的增加而减小[17]。
(4)种子单粒质量是种子的重要生物学特性之一[35]。其表征了种子内营养物质的多少,对种子萌发、幼苗定居及存活具有很大影响[36-38]。相对于球果性状及一般的种子性状,种子单粒质量较难测定,需要专门的称量仪器才能进行(如万分之一电子天平),而这一条件在一般情况下较难满足。因此,以易于测定的球果性状及一般的种子性状参数来估算种子单粒质量是一种较好的间接方法。在相关分析中,果长、果宽、果质量、果长×果宽、种长、种宽、种厚、种长×种宽×种厚等8个性状与种子单粒质量的相关性均达极显著水平;建立种子单粒质量与这8个性状的多元回归模型,发现对种子单粒质量贡献率排名前4的性状分别为种长、种厚、果质量及种长×种宽×种厚,其贡献率分别为28.43%、27.65%、25.10%及10.92%,这4个性状的总贡献率达92.1%。为进一步了解这4个性状对种子单粒质量的预测精度及彼此之间的关系,需建立相应的线性回归模型。分析结果表明,种子单粒质量与果质量、种长、种厚及种长×种宽×种厚间方程的决定系数分别为0.618、0.529、0.428和0.666。其中,果质量及表示种子大小的种长×种宽×种厚的决定系数要高于种长和种厚,说明果质量和种长×种宽×种厚对种子单粒质量的预测效果要优于种长和种厚,其能更好地反映种子单粒质量的变化趋势。
[1]唐万鹏,史玉虎, 漆 良, 等. 辐射松、马尾松、长叶松与晚松人工林的生长规律比较研究[J]. 湖北林业科技, 2004,129(3):1-4.
[2]汪企明, 李晓储, 孟在祥, 等. 火炬松、湿地松、短叶松、晚松在宝应京杭大运河堤引种研究[J]. 江苏林业科技, 1994(4):11-14.
[3]李锦清, 郑勇平. 长乐林场湿地松火炬松晚松生长规律的调查分析[J]. 浙江林学院学报, 1986, 3(1): 59-64.
[4]欧阳菁. 晚松萌蘖机理及扦插技术研究[D]. 南昌:江西农业大学, 2012.
[5]宋永庆, 周克熙, 余翔华, 等. 晚松扦插试验简报[J]. 江西林业科技, 1992(4): 35-36.
[6]阙国宁, 张守英. 晚松悬浮细胞系的建立和原生质体的分离[J]. 林业科学研究, 2002, 15(2): 247-251.
[7]阙国宁, 房建军, 葛万川, 等. 火炬松、湿地松、晚松组培繁殖的研究[J]. 林业科学研究, 1997, 10(3): 227-232.
[8]程 磊, 刘苑秋, 文 娟. 盐胁迫对晚松生理特性的影响[J].江西农业大学学报, 2006, 28(3): 382-385.
[9]黄桂萍, 刘苑秋, 程 磊. 土壤铅污染对晚松生理特性的影响[J]. 江西农业大学学报, 2006, 28(6): 833-837.
[10]柳恒饶, 刘光斌, 李林检, 等. 响应面法分析优化晚松生物质成型燃料制备工艺[J]. 林业工程学报, 2016, 1(1): 93-99.
[11]陈丹凤, 刘苑秋, 黄国贤, 等. 能源树种晚松生物量及其能源利用研究[J]. 林业实用技术, 2014(9):21-24.
[12]Hornung C, Sosa V. Morphological variation in Puya(Bromeliaceae): an allometric study[J]. Plant Systematics and Evolution, 2006, 256: 35-53.
[13]Feil R, Fraga M F. Epigenetics and the environment: emerging patterns and implications[J]. Nat. Rev. Genet., 2012, 13:97-109.
[14]Keppel G. Morphological variation, an expanded description and ethnobotanical evaluation ofCycas seemanniiA. Braun(Cycadaceae) [J]. South Paci fi c Journal of Natural Science, 2009,27: 20-27.
[15]Khalil M A K. Genetic of cone morphology of Black Spruce(Picea mariana) in New foundland, Canada[J]. Silvae Genetica,1984, 33: 4-5.
[16]Li C X. A review of the studies on the phenotypic variation of forest trees in natural stands[J]. Forest Research, 1988, 1(6): 657-664.
[17]徐 亮, 包维楷, 何永华. 4个岷江柏种群的球果和种子形态特征及其地理空间差异[J]. 应用与环境生物学报, 2004,10(6): 707-711.
[18]张飞琳, 郭美丽, 齐天进, 等. 太白山油松球果和种子形态变异分析[J]. 陕西林业科技, 2007(4): 1-4, 28.
[19]文珊娜, 姜清彬, 仲崇禄, 等. 灰木莲不同种源种子形态变异分析[J]. 中南林业科技大学学报, 2016, 36(7): 7-11.
[20]王旭军, 张日清, 许忠坤, 等. 红榉不同种源种子形态性状变异[J]. 中南林业科技大学学报, 2015, 35(1): 1-6.
[21]刘仁林, 胡明娇, 李 江, 等. 乌饭树果实大小的地理变异研究[J]. 经济林研究, 2016, 34(3): 114-120.
[22]林 萍, 姚小华, 曹永庆, 等. 油用牡丹‘凤丹’果实性状及其脂肪酸组分的变异分析[J]. 经济林研究, 2015,33(1):67-72.
[23]Beaulieu J, Simon J P. Variation in cone morphology and seed characters inPinusstrobe in Quebec[J]. Can. J. Bot, 1995, 73:262-271.
[24]Maley M L, Parker W H. Phenotypic variation in cone and needle characters ofPinus banksiana(jack pine) in northwestern Ontario[J]. Can. J. Bot, 1993, 71: 43-51.
[25]王小平, 刘晶岚, 王九龄, 等. 白皮松种子及球果形态特征的地理变异[J]. 北京林业大学学报, 1993, 20(3): 25-31.
[26]Francisco Limo´n, Jorge Gonza´lez-Astorga, Fernando Nicolalde-Morejo´n,et al. Phenotypic variation ofZamia loddigesiiMiq. andZ. prasinaW.Bull. (Zamiaceae, Cycadales):the effect of environmental heterogeneity[J]. Plant Syst. Evol.,2016, 302:1395-1404.
[27]Pe´rez-Farrera M A, Vovides A P, Avendano S. Morphology and leaflet anatomy of theCeratozamia norstogii(Zamiaceae,Cycadales) species complex in Me´xico with comments on relationships and speciation[J]. Int. J. Pl Sci., 2014, 175:110–121.
[28]Prabodh K, Bajpai, Ashish R,et al.High phenotypic variation inMorus albaL. along an altitudinal gradient in the Indian trans-Himalaya[J]. Journal of Mountain Science, 2015, 12(2):446-455.
[29]Janzen D H. Variation in seed size within a crop of a Costa Rican Mu-cuna and reana (Leguminosae) [J]. Am. J. Bot., 1977, 64:347-349.
[30]Willson M F. Plant reproductive ecology[M]. New York: John Wiley& Sons, 1983: 1-90.
[31]马绍宾, 姜汉侨. 小檗科鬼臼亚科种子大小变异式样及其生物学意义[J]. 西北植物学报, 1999, 19(4): 715-724.
[32]王庆华, 刘永刚, 沈立新, 等. 秃杉球果形态的变异研究[J].西部林业科学, 2013, 42(5): 47-52.
[33]齐立志, 刘吉雨, 刘桂丰, 等. 长白落叶松雌雄花及球果、种子性状的变异[J]. 东北林业大学学报, 1995, 23(2): 7-12.
[34]王润辉, 胡德活, 郑会全, 等. 江南油杉球果种子特性种源变异分析[J]. 中南林业科技大学学报, 2013, 33(2): 19-24.
[35]Silvemown J W. Seed size, life span and germination date as coadapted femurs of plant, life history[J]. Nature, 1981,18:860-864.
[36]Ove Eriksson. Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis[J]. Acta Oecol, 1999, 20(1): 61-66.
[37]Ekta Khurana, Singh J S. Influence of seed size on seedling growth ofAlbizia proceraunder different soil water levels[J].Ann. Bot., 2000, 86: 1185-1192.
[38]Raquel G R, Keith R P, Malcolm E R,et al. Effect of seed size and testa colour on saponin content ofSpanish lentilseed[J].Food Chem., 1997, 58(3): 223-226.
猜你喜欢
四川蚕业(2022年1期)2022-06-06
中国食品(2021年21期)2021-11-07
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
农机使用与维修(2020年6期)2020-06-11
四川蚕业(2020年4期)2020-02-10
中国森林病虫(2019年1期)2019-03-04
中国林副特产(2019年3期)2019-01-10
百科知识(2015年18期)2015-09-10
少儿科学周刊·儿童版(2014年1期)2014-03-10
