
草菇芳香醇氧化酶基因vvaao1的序列特征与差异表达
2018-05-07 08:33严俊杰仝宗军刘媛媛张磊张云谢宝贵
生物技术通报 2018年4期
严俊杰 仝宗军 刘媛媛 张磊 张云 谢宝贵
(福建农林大学生命科学学院菌物研究中心,福州 350002)
草菇(Volvariella volvacea)又称中国蘑菇,是一类重要的高温褐腐型食用菌,在热带、亚热带地区广泛种植[1]。草菇可以利用稻草、废棉、玉米芯、甘蔗渣、药渣以及各种食用菌栽培菌渣等为主料进行栽培,不仅可以实现农业废料的再次利用,还可带来巨大的经济效益,变废为宝[2-3]。然而,草菇的生物转化率较低,通常只能达到20%左右,制约了草菇的高效生产和产业的发展[4]。对基质降解相关酶及其编码基因的研究与改造以提高草菇基质降解能力是提高其生物转化率的有效手段。
植物组织是草菇栽培料的重要组成成分,也是草菇碳源的主要来源。在植物组织中,木质素和半纤维素以共价键形式接合,将纤维素分子包埋于其中,形成坚固的天然屏障,阻碍了微生物进入其中分解纤维素[5]。因此,木质素降解能力的高低是决定草菇能否高效利用纤维素获取子实体发育所需碳源的关键。芳香醇氧化酶(Aryl alcohol oxidase,AAO,EC 1.3.3.7)属于 GMC(Glucose-methanol-choline)氧化还原酶超家族,是一种分泌型黄素蛋白酶,其氧化苯甲醇释放出来的过氧化氢可以与过氧化物酶及漆酶相互作用形成羟基自由基用于降解木质纤维素;同时,AAO还可以抑制漆酶氧化产物的再聚合,以维持对木质纤维素的持续降解[6]。随着基因组测序技术的发展,已有大量食用菌的芳香醇氧化酶编码基因获得注释[7]。Varela等[8]在 2000 年成功解析出侧耳属Pleurotus pulmonarius的AAO晶体结构,这为食用菌AAO的结构及木质素降解机制研究提供了重要的参考。本实验室已对草菇同核体菌株PYd21的基因组进行了de novo测序,并对同、异核体菌株进行转录组、表达谱及蛋白质组测序[9,10]。基于这些组学数据,我们获得了一个芳香醇氧化酶编码基因vvaao1,并对其序列特征、表达情况进行系统的分析,以期为深入研究vvaao1在草菇基质降解和子实体发育过程中的生物学功能提供依据。
1 材料与方法
1.1 材料
供试草菇菌株PYd21、PYd15为福建省主栽品种“屏优一号”两个可正常配对的单孢,配对纯化后获得杂交菌株H1521。对PYd21菌丝进行DNA提取并采用Illumina平台进行de novo测序,SOAPdenovo拼接结果上传至NCBI,登录号为:PRJNA171553。对PYd21、PYd15、H1521菌丝以及H1521的子实体菌柄样品等量混合后进行转录组测序;并对3个菌株分别进行数字基因表达谱(Digital gene expression profiling,DGE)及蛋白质组(iTRAQ)测定。
1.2 方法
1.2.1 基因序列获取与结构分析 以基因组注释获得的vvaao1序列及其上下游2 000 bp为参考(Reference sequence),采用 ZOOM 软件[11]将混合转录组测序获得的clean reads与之定位获取转录本的全长区段;以全长转录本及其上下游500 bp为reference进行基因组 reads定位,验证序列的准确性;验证准确的序列再次进行转录组reads定位,对reads规律性错配区域进行分析并结合GT-AG保守结构判断内含子位置;软件设置与内含子判断按照严俊杰等的方法进行[12],基因组reads定位时设置错配碱基数为0,转录组reads定位设置错配碱基数为40。获得DNA序列的相关信息后,用DNAMAN软件进行序列结构模拟图绘制。
1.2.2 氨基酸序列生物信息学分析 氨基酸序列的理化性质采用Expasy ProtParam(https://web.expasy.org/cgi-bin/protparam/protparam)分析,采用SignalP-4.1(http://www.cbs.dtu.dk/services/SignalP)预测信号肽,亚细胞定位采用TargetP 1.1 Server(http://www.cbs.dtu.dk/services/TargetP/) 分 析;蛋白质三级结构的比较分析采用Phyre 2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi? id=index) 进行 ;从 NCBI(https://www.ncbi.nlm.nih.gov/pubmed/)、UniProt(http://www.uniprot.org/)网站获取白腐真菌GMC氧化还原酶超家族氨基酸序列,同时根据文献信息[13]下载目前已报道具备AAO序列的三个褐腐 菌(Gloeophyllum trabeum、Fomitopsis pinicola及Rhodonia placenta)基因组数据,并获取相应的AAO氨基酸序列,应用MEGA 5.1[14]进行MUSCLE法比对并构建进化树分析。
1.2.3vvaao1转录水平的差异表达分析 应用ZOOM软件将PYd15、PYd21、H1521三个菌株的数字基因表达谱clean tags与vvaao1基因全长转录本进行定位,设置错配为0。参照刘朋虎等[15]进行表达量标准化及差异表达分析,获得vvaao1在不同菌株的TPM值(transcript per million clean tags)。
采用实时荧光定量PCR(RT-qPCR)技术进一步验证vvaao1基因的表达模式。收集固体PDA上生长相同时间的菌丝样品以及H1521不同发育时期的子实体样品,液氮速冻后用于总RNA提取。分别采用E.Z.N.A.TMPlant RNA kit试剂盒(美国Omega Bio-Tek公司)和TransScript All-in-one First-Strand cDNA Synthesis SuperMix for qPCR(One Step gDNA Removeal)试剂盒(北京全式金公司)提取总RNA并进行第一链cDNA合成,操作均按照说明书进行。实时荧光PCR采用TransStart Top Green qPCR SuperMix(北京全式金公司)定量试剂盒,在CFX96实时荧光定量PCR仪(美国BIO-RAD公司)进行,以甘油醛-3-磷酸脱氢酶基因vv-gapdh为内参,退火温度设为60℃,循环数设为40。应用Primer 6.0设计定量PCR引物并委托上海生工生物工程有限公司合成,引物序列见表1。每种样品进行3个生物学重复,采用2-ΔΔCt法[16]计算基因的相对表达量。

表 1 荧光实时定量PCR所使用的引物
1.2.4vvaao1翻译水平的差异表达分析 采用iTRAQ获得草菇蛋白质组数据,以草菇PYd21的基因组序列预测获得的氨基酸序列为模版,应用PEAKS stutio6.0软件[17],将带有不同iTRAQ标记的肽段进行匹配,并获得可视化的蛋白相对表达量。具体参数设置参见文献[18]。
1.2.5 菌丝生长速度测定与出菇试验 参考王圣铕等[19]的方法配制大试管栽培料,高压灭菌后打孔接种活化两次后的草菇菌丝,于34℃避光培养;参照朱坚等[20]的方法测定菌丝在栽培料上的生长速度;参考陆娜等[21]的方法进行栽培料制作和出菇管理。
1.2.5 统计分析 采用 SPSS 2.0软件对数据结果进行统计学分析,差异显著性用 LSD方法进行分析比较。采用皮尔森相关系数(Pearson correlation coefficient)分析菌丝生长速度与vvaao1表达量的相关性。
2 结果
2.1 vvaao1基因序列验证与结构分析
根据基因组注释结果,vvaao1序列位于scaffold 37上52 319-54 654 bp之间,全长2 336 bp。经过转录组reads定位后获得vvaao1序列的实际全长为3 091 bp,与预测序列相比5'UTR区域和3'UTR区域分别延长了278 bp和477 bp。将基因组测序获得的clean reads与上下游分别延长500 bp后的DNA序列进行定位,结果如图1所示,vvaao1基因的全长均被reads所覆盖,最低覆盖reads数为7条,说明vvaao1的DNA序列正确,可以用于后续分析。

图1 基因组reads与vvaao1序列的定位情况
对基因结构的分析结果如图2所示:vvaao1有9个外显子,编号为E1-E9;8个内含子,编号为I1-I8,内含子长度分别为:53、52、55、55、49、55、61、153 bp。将去除所有内含子的转录本提交ORF Finder,预测获得一条完整的开放阅读框(ORF),全长1 803 bp,5'端非翻译区(5'UTR)长度为278 bp;3'端非翻译区(3'UTR)全长477 bp。基因及氨基酸编码序列均上传NCBI,并获得GenBank登录号为MG938583。
2.2 VvAAO1蛋白的生物信息学分析

图2 草菇vvaao1基因结构模拟图
草菇VvAAO1蛋白由600个氨基酸残基组成,分子量约为65 kD,理论等电点为4.49。信号肽预测结果如图3所示:VvAAO1氨基酸序列的N端包含一段18个氨基酸残基的分泌信号肽,属于分泌蛋白。亚细胞定位的结果也支持该蛋白位于分泌通路(Secretory pathway),得分(NN scores)为0.906,RC值(Relative Class)为2。推测该蛋白可能通过N端信号肽引导分泌到细胞外行使其生物学功能,这与芳香醇氧化酶家族的特征相吻合。

图3 VvAAO1氨基酸序列的信号肽预测结果
以PDB(Protein data bank)数据库上二级、三级结构已获得解析的Pleurotus eryngii芳香醇氧化酶模式蛋白3FIM为参考,对VvAAO1二级结构预测后进行三级结构建模。结果如图4-A所示,成熟的VvAAO1蛋白包含20个α-螺旋和19个β-折叠。与3FIM相比,一致性(Identity)达到48%,置信度(Confidence)达到100%;其中,3FIM的预测结果(3FIM-predict)在第205、355个氨基酸附近较VvAAO1多了一个α-螺旋,在第500个氨基酸附近多了一个β-折叠结构,其余结构基本一致(图4-A,C);然而,3FIM的预测结果与其自身的实际结构(3FIM-real)仍存在一些差异(图4-B,4-C)。

图4 VvAAO1与芳香醇氧化酶模式蛋白3FIM的三级结构比较
采用最大似然法(Maximum Likelihood,ML)构建的系统发育树如图5所示:所有38条真菌GMC超家族蛋白可以分为AAO、PDH、MOX、CDH、POX四个家族分支,自检1 000次的bootstrap值均在97%以上。草菇VvAAO1序列位于AAO家族分支上,与侧耳属菌株P. eryngii和P. pulmonarius的AAO序列最为接近,但与褐腐菌Gloeophyllum trabeum、Fomitopsis pinicola及Rhodonia placenta的AAO序列关系较远。
2.3 草菇vvaao1基因在不同菌株之间的差异表达
PYd15、PYd21及H1521的表达谱分别有263、3和13 453个Clean Tag能够正向定位于vvaao1的转录本上,得到标准化后的TPM值分别为:45.71、0.51、2 359.43;荧光定量PCR的结果(图6-A)显示H1521的表达量是PYd15的167倍、PYd21的71倍。蛋白质组iTRAQ数据也支持H1521的表达量最高且是两个同核体菌株之和的2.13倍,存在协同增效作用(图6-B)。
2.4 菌丝在栽培料上的生长速度与出菇能力
如图6-C所示,H1521在栽培料上生长良好,生长速度为1.2 cm/d,是PYd15的1.46倍,是PYd21的7.50倍;PYd21的生长速度最慢,仅为0.16 cm/d。菌丝生长速度与vvaao1的DGE、RT-qPCR以及iTRAQ结果的皮尔森相关系数为分别为:0.789、0.774及0.758,均达到强相关水平。3个菌株的出菇情况如图6-D-F所示,仅H1521能够出菇,PYd15及PYd21的料面菌丝稀少均无法形成原基。
2.5 vvaao1在草菇不同发育时期的差异表达

图5 VvAAO1与真菌GMC氧化还原酶超家族序列的系统发育树
对异核体菌株H1521进行出菇并取样。结果如图7-A-E所示,草菇子实体发育可以分为原基、纽扣期、蛋形期、伸长期和成熟期5个时期;其中,蛋形期、伸长期和成熟期具有菌盖和菌柄的明显分化,分别取样。对以上样品进行RT-qPCR分析,结果如图7-F所示:vvaao1在原基的表达量最高,且与其他各个时期均存在显著性差异,在成熟期的样品中几乎不表达;另外,vvaao1在同一时期的菌盖和菌柄的表达量差异不显著。
3 讨论
基质降解能力的强弱直接关系到食用菌栽培的产量和效益。草菇作为褐腐真菌,具备较为完善的纤维素降解系统[9];然而,前期的研究结果表明,草菇缺乏锰过氧化物酶、木质素过氧化物酶活性,仅具备漆酶活性,对木质素的降解能力较弱,是草菇降解基质、吸收营养的限制因子,也是草菇生物转化率低的原因之一[4,22]。木质素主要由芳香醇聚合而成[23],作为木质素降解的辅酶,芳香醇氧化酶可以分泌到细胞外,与漆酶协同降解木质素[24]。Carro等[13]2016年对真菌芳香醇氧化酶的研究进行了综述,发现AAO在白腐菌中的分布更为广泛,褐腐菌仅Gloeophyllum trabeum、Fomitopsis pinicola及Rhodonia placenta的基因组中存在1-2个AAO编码基因。本研究首次从褐腐菌草菇中获得一个芳香醇氧化酶基因vvaao1,其编码的蛋白具备信号肽、亚细胞定位与分泌途径,可以有效分泌到细胞外降解基质中的木质素;系统发育树和三级结构均显示VvAAO1与白腐菌P.eryngii芳香醇氧化酶模式蛋白3FIM比较相似,且蛋白的大小、等电点等理化性质也比较接近[25],推测草菇VvAAO1可能具备与3FIM较为接近的酶学功能。
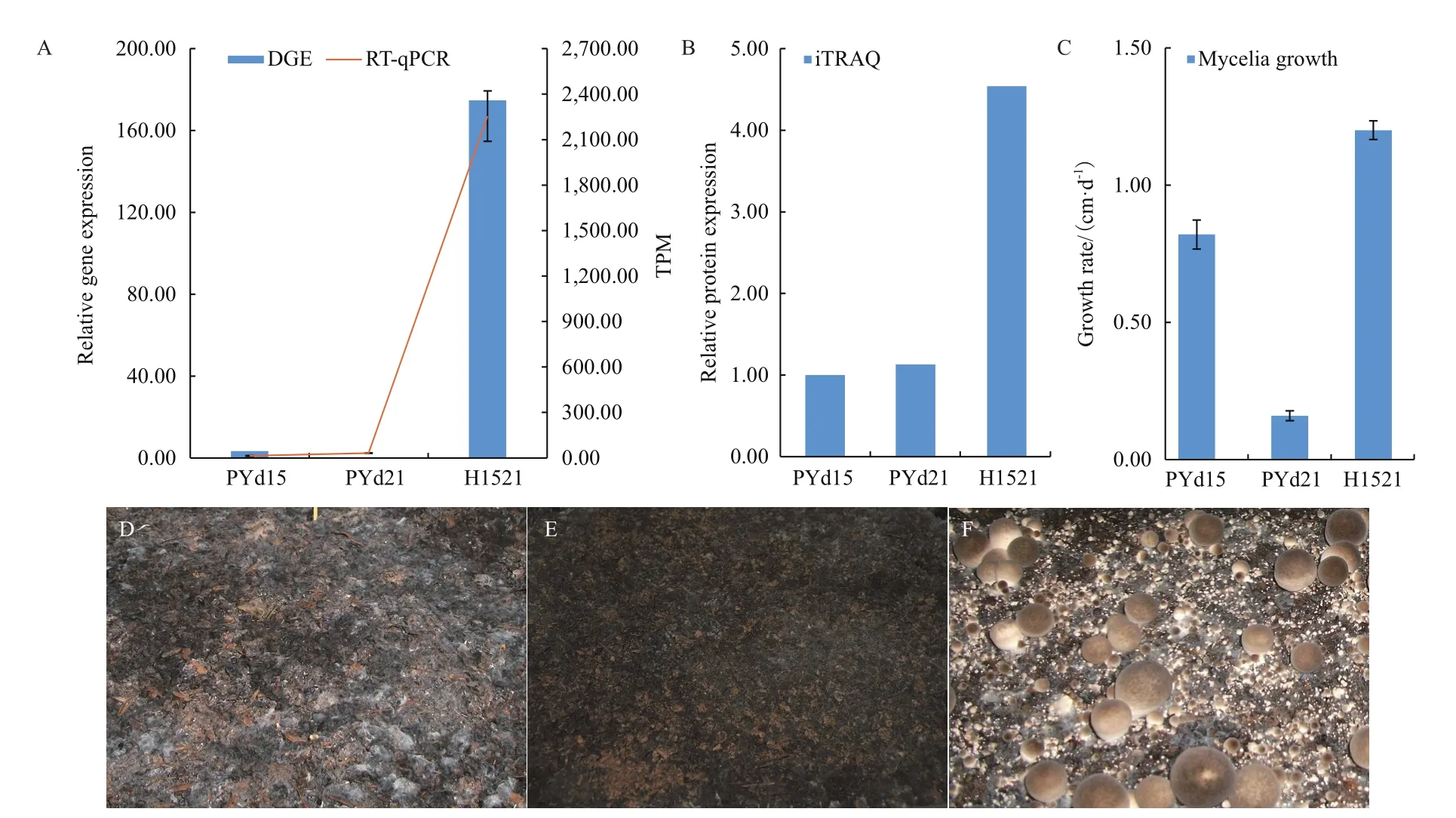
图6 不同草菇菌株的生长、出菇及vvaao1基因的表达情况

图7 vvaao1基因在草菇H1521不同子实体发育阶段的差异表达
不同药物或外源分子与病毒或毒素之间的互作可以有效增强对机体的侵染和毒效,产生协同增效作用[26-27]。本研究发现vvaao1基因的表达量在两个同核体菌株交配后显著上调,且远高于两个同核体菌株表达量之和。进一步用DNAMAN比对PYd15与PYd21的vvaao1基因开放阅读框及其上游2 000 bp调控区的DNA序列,发现两个菌株的序列完全一致,说明协同增效作用并不是其自身序列的作用结果;因为菌丝阶段的细胞核并未发生融合,推测有可能是细胞质中转录因子蛋白不同亚基的互作促进了下游基因的表达。这种异核体菌株基因发生协同增效表达的现象在草菇中较为常见[15,18,28-30],然而其作用机制仍进一步探索和揭示。另外,vvaao1基因在可正常出菇的异核体菌株H1521中表达量显著高于两个不能出菇的同核体菌株,且对子实体不同发育时期的定量检测结果也显示该基因在子实体形成的原基期最高表达;推测vvaao1的高表达可能有利于草菇子实体的形成。
4 结论
草菇芳香醇氧化酶VvAAO1为分泌蛋白,氨基酸序列的三级结构与白腐菌P.eryngii芳香醇氧化酶模式蛋白3FIM接近;该酶的编码基因在可正常出菇的异核体菌丝中存在协同增效表达,且在子实体发育过程的原基期最高表达。
[1] Tao Y, Xie B, Yang Z, et al. Identification and expression analysis of a new glycoside hydrolase family 55 exo-β-1, 3-glucanaseencoding gene inVolvariella volvaceasuggests a role in fruiting body development[J]. Gene, 2013, 527(1):154-160.
[2] 李正鹏, 余昌霞, 李巧珍. 草菇工厂化栽培原材料研究进展[J].农业工程技术:温室园艺, 2017, 37(19):67-70.
[3] 关雁桃, 罗国杨, 董炳炎, 等. 变废为宝--广东菇农利用药渣栽培草菇技术[J]. 当代蔬菜, 2005(10):27-27.
[4] 朱刚, 吴林, 陈明杰, 等. 草菇锰过氧化物酶编码基因生物信息学分析及其转录水平和酶活性的测定[J]. 菌物学报, 2013,32(5):919-927.
[5] 杨晔. 木质素降解酶研究进展[J]. 农业工程, 2014(4):48-51.
[6] Goswami P, Chinnadayyala SSR, Chakraborty M, et al. An overview on alcohol oxidases and their potential applications[J]. Applied Microbiology and Biotechnology, 2013, 97(10):4259-4275.
[7] Hernández-Ortega A, Ferreira P, Martínez AT. Fungal aryl-alcohol oxidase:a peroxide-producing flavoenzyme involved in lignin degradation[J]. Applied Microbiology and Biotechnology, 2012,93(4):1395-1410.
[8] Varela E, Böckle B, Romero A, et al. Biochemical characterization,cDNA cloning and protein crystallization of aryl-alcohol oxidase fromPleurotus pulmonarius[J]. Biochimica et Biophysica Acta(BBA)-Protein Structure and Molecular Enzymology, 2000, 1476(1):129-138.
[9] Chen B, Gui F, Xie B, et al. Composition and expression of genes encoding carbohydrate-active enzymes in the straw-degrading mushroomVolvariella volvacea[J]. PLoS One, 2013, 8(3):e58780.
[10] 刘靖宇, 江玉姬, 谢宝贵, 等. 应用 iTRAQ 结合 2D LC-MS/MS技术分析草菇同核和异核菌丝蛋白质组[J]. 食用菌学报,2012, 19(1):1-6.
[11] Zhang Z, Lin H, Ma B. ZOOM Lite:next-generation sequencing data mapping and visualization software[J]. Nucleic Acids Research, 2010, 38:W743-W748.
[12] Yan JJ, Zhang L, Wang RQ, et al. The sequence characteristics and expression models reveal superoxide dismutase involved in cold response and fruiting body development inVolvariella volvacea[J]. International Journal of Molecular Sciences, 2016,17(1):34.
[13] Carro J, Serrano A, Ferreira P, et al. Fungal aryl-alcohol oxidase in lignocellulose degradation and bioconversion[M]//GuptaVK.Microbial Enzymes in Bioconversions of Biomass. Switzerland:Springer International Publishing, 2016:301-322.
[14] Tamura K, Peterson D, Peterson N, et al. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance, and maximum parsimony methods[J].Molecular Biology and Evolution, 2011, 28(10):2731-2739.
[15] 刘朋虎, 谢宝贵, 邓优锦, 等. 草菇gpi基因结构及其异核体表达协同增效作用[J]. 食用菌学报, 2011, 18(4):1-5.
[16] Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△CTmethod[J].Methods, 2001, 25:402-408.
[17] Ma B, Zhang K, Hendrie C, et al. PEAKS:Powerful Software for Peptide De Novo Sequencing by MS/MS[J]. Rapid Communications in Mass Spectrometry, 2003, 17(20):2337-2342.
[18] 严俊杰, 郭丽羡, 赵静静, 等. 草菇谷胱甘肽 S-转移酶编码基因(vv-gto1)的序列分析及其差异表达[J], 2014, 54(10):1171-1177.
[19] 王圣铕, 廖伟, 黄千慧, 等. 生理生化指标与草菇开伞时间的相关性分析[J]. 基因组学与应用生物学, 2016, 35(4):970-978.
[20] 朱坚. 食用菌品种特性与栽培[M]. 福州:福建科学技术出版社, 2011.
[21] 陆娜, 周祖法, 袁卫东, 等. 不同草菇菌株的室内床架式栽培试验[J]. 食药用菌 , 2010, 18(3):28-29.
[22] 吴林, 朱刚, 陈明杰, 等. 草菇基因组中 11 个漆酶同源基因的生物信息学分析及铜离子对其表达的影响[J]. 菌物学报,2014, 33(2):323-333.
[23] 闫强, 王安建, 王高尚, 等. 全球生物质能资源评价[J]. 中国农学通报, 2009, 25(18):466-470.
[24] Ruiz-Dueñas FJ, Martínez ÁT. Microbial degradation of lignin :how a bulky recalcitrant polymer is efficiently recycled in nature and how we can take advantage of this[J]. Microbial Biotechnology, 2009, 2(2):164-177.
[25] 吴茵, 陈敏, 郭倩. 刺芹侧耳芳基醇氧化酶的分离纯化及其酶学性质[J]. 菌物学报, 2016, 35(6):705-713.
[26] 项林平, 刘小芸, 田强强, 等. 棉铃虫核型多角体病毒几丁质酶的原核表达及其协同增效作用[J]. 西北农林科技大学学报:自然科学版, 2011, 39(2):87-91.
[27] 王莹莹, 任相亮, 姜伟丽, 等. 鳢肠乙醇粗提物与 Cry1Ca 的协同增效及对甜菜夜蛾 SeABCC1-SeABCC3 mRNA 表达量的影响[J]. 中国生物防治学报, 2017, 33(4):446-453.
[28] Meng L, Yan J, Xie B, et al. Genes encoding FAD-binding proteins inVolvariella volvaceaexhibit differential expression in homokaryons and heterokaryons[J]. Microbiological Research,2013, 168(8):533-546.
[29] 刘朋虎, 邓优锦, 江玉姬, 等. 草菇TPI基因的克隆, 结构及其在同核, 异核菌株中的表达量[J]. 福建农林大学学报:自然科学版, 2013(1):63-67.
[30] 刘朋虎, 谢宝贵, 邓优锦, 等. 草菇磷酸果糖激酶(PFK)基因克隆, 结构及不同菌株中表达量分析[J]. 菌物学报,2013, 32(2):253-260.
猜你喜欢
南方农机(2023年3期)2023-01-31
今日农业(2022年3期)2022-11-16
昆明医科大学学报(2020年12期)2021-01-26
江苏广播电视报·新教育(2021年34期)2021-01-03
饮食与健康·下旬刊(2019年3期)2019-03-08
现代农村科技(2019年11期)2019-01-06
食用菌(2018年2期)2018-01-31
上海农业学报(2017年4期)2017-04-10
上海农业学报(2017年3期)2017-04-10
中国粮油学报(2016年5期)2016-01-23
