
同/异型乳酸菌对青贮玉米开窖后品质及微生物的影响
2018-06-05 06:55王旭哲张凡凡马春晖唐开婷李菲菲
农业工程学报 2018年10期
王旭哲,张凡凡,马春晖,苗 芳,唐开婷,李菲菲
(石河子大学动物科技学院,石河子 832003)
0 引 言
随着畜牧业的发展,玉米青贮饲料的重要性日益突显[1-2]。玉米青贮主要依靠乳酸菌厌氧发酵产生乳酸降低pH值,并通过降低叶片呼吸和酶活性,抑制霉菌、梭状芽孢杆菌、酵母菌和其他不利微生物生长,从而使玉米青贮得以保存[3-4],因此乳酸菌被认为是青贮饲料发酵过程中最重要的因素之一[5]。其发酵产生高浓度的乳酸,但在开窖后,不良微生物的大量消耗致使青贮饲料有氧稳定性降低,即青贮过程中微生物群落的变化将影响最终青贮饲料的品质和有氧稳定性[6-8]。主要由于酵母菌等微生物在好氧条件中利用这些化合物作为底物产生CO2、水和热量,进而导致pH值上升,致使不耐低pH值的丝状真菌和芽孢菌等有害菌生长增殖[9]。
为提高玉米青贮有氧稳定性,很多学者提出使用乳酸菌添加剂[10-11],认为乳酸菌添加剂能有效地改善发酵青贮饲料的质量,主要通过改变pH值,抑制有害的附生微生物生长来提高有氧稳定性[12-14]。Kharazian等[4]从伊朗玉米青贮饲料中分离得到的乳酸菌具有较高的抗真菌活性,可用作青贮饲料添加剂提高青贮玉米的质量和有氧稳定性。青贮饲料引入添加微生物菌剂这一概念是为了控制发酵,加入快速发酵的同型乳酸菌提高青贮饲料品质[15]。根据Yuan等[5]研究可知,青贮饲料添加同型乳酸菌植物乳杆菌(Lactobacillus plantarumas)导致乳酸质量分数较对照有所提高。Liu等[16]的研究中同样发现相对于未经处理的青贮饲料,接种植物乳杆菌能提高青贮饲料发酵品质。此外,乳酸杆菌属(Lactobacillus)或片球菌属(Pediococcus)的其他菌也经常被使用[17]。但同型乳酸菌对于提高有氧稳定性作用不显著,近年来为提高有氧稳定性并降低青贮发酵损失,异型发酵乳酸菌物种如布氏乳杆菌(Lactobacillus buchneri)作为青贮添加剂添加到青贮饲料中[15,18]。布氏乳杆菌可产生抗菌的乙酸和细菌素等副产物,减少有氧恶化并改善青贮品质[19]。微生物菌剂必须具备足够数量以有效控制发酵,目前普遍认为1×105cfu/g的乳酸菌添加量可以压制青贮附生乳酸菌成为优势种群[20-21]。因此,应用适当的乳酸菌接种量可以确保青贮饲料质量,防止青贮饲料变质,提高经济效益[22]。此外,已经提出同型发酵乳酸菌与布氏乳杆菌复合进行添加,因为这 2类微生物的结合有可能提高发酵速度并提高有氧稳定性[23-24]。然而,2种同型发酵乳酸菌共同添加或2种同型发酵乳酸菌与布氏乳杆菌复合添加提高玉米青贮有氧稳定性及发酵特性的能力是未知的。
基于此,本研究设置添加不同发酵类型的乳酸菌处理,探究同时添加 2种同型发酵乳酸菌、单一添加布氏乳杆菌以及 2种同型发酵乳酸菌与布氏乳杆菌复合添加对开窖后玉米青贮发酵品质、微生物数量及有氧稳定性的影响。旨在明确添加不同发酵类型的乳酸菌处理在玉米青贮开窖后,能否改善发酵品质、降低有害微生物数量、提高有氧稳定性,为生产实践提供参考。
1 材料与方法
1.1 材料和样地
试验选用新饲玉10号作为青贮原料。试验选定在石河子大学的牧草试验站(N 44°20′,E 88°30′,H 420 m)开展。其生长期:4月10日~8月20日(2015年),共计112 d,青贮玉米的刈割期定于乳熟末期/蜡熟初期进行(玉米乳线超过2/3时),收割全株玉米,将青贮玉米切碎至1~2 cm。真空袋规格为50 cm×40 cm×0.24 mm。全株玉米青贮原料营养成分和微生物数量见表1。
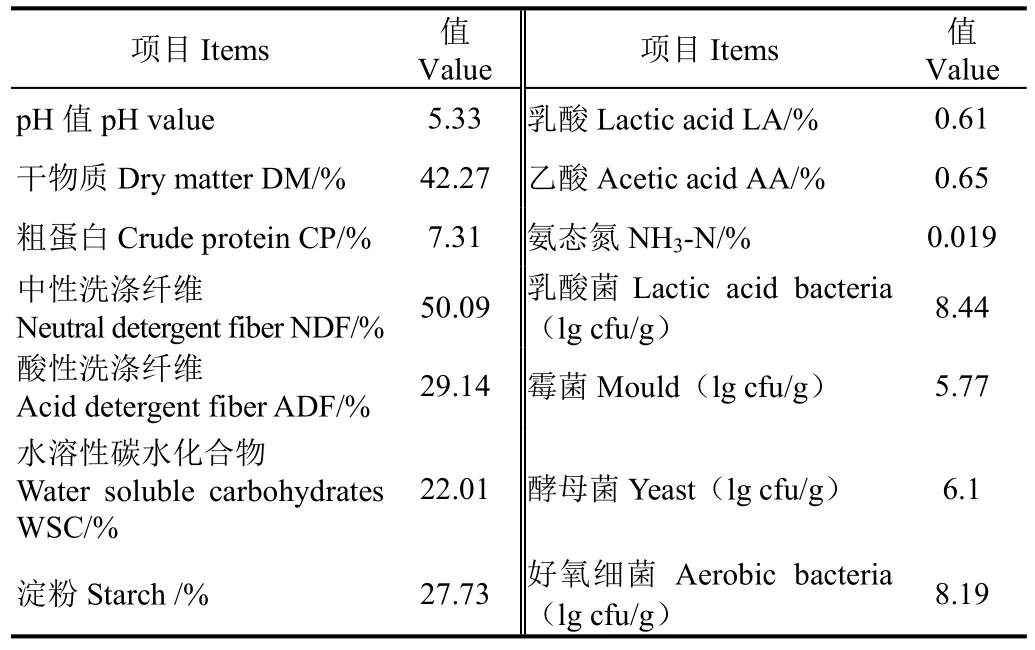
表1 玉米青贮原料营养组成和微生物数量Table 1 Nutrients composition and microbial populations of corn silage of ensiling material
1.2 试验设计
试验将添加乳酸菌设计为4个处理,CK处理(不加任何菌剂);T处理(同型发酵乳酸菌):Lactobacillus plantarum和Pediococcus acidilactici复合添加,添加量为1∶1,1×105cfu/g;Y 处理(异型发酵乳酸菌):Lactobacillus buchneri,添加量为1×105cfu/g;TY处理(同型+异型发酵乳酸菌):Lactobacillus plantarum、Pediococcus acidilactici和Lactobacillus buchneri复合添加,添加量为1∶1∶1,1×105cfu/g。并将各菌种在MRS(de man rogosa sharpe)液体培养基中培养,平板计数后确定其数量,按比例配置菌液。用灭菌喷壶均匀的喷洒至待贮青贮玉米表面(CK处理喷洒等量的去离子水)。喷洒完毕后立刻进行青贮。采用真空袋法调制,每个处理4袋,每袋2.0 kg,室温(25±3)℃下发酵60 d后(开窖0 h)开窖。检测开窖后第0、48、96、144、192,240 h青贮pH值、可溶性碳水化合物(water soluble carbohydrate,WSC)、乳酸(lactic acid,LA)、乙酸(acetic acid,AA)、氨态氮(NH3-N)、乳酸菌、霉菌、酵母菌以及好氧细菌数量的变化,对 CO2产量进行分析。开窖全程动态监测各处理青贮温度变化。应用隶属函数法对青贮发酵品质及微生物数量进行综合评判。
1.3 测定方法
DM 采用 105 ℃烘干法测定[25];pH 值利用酸度计(PHS-3C,上海雷磁)测定;CP测定采用凯氏定氮法[10];NDF和ADF采用范氏法(Van Soest)[25];WSC测定采用蒽酮比色法测定[26];有机酸(乳酸、乙酸、丙酸、丁酸)用液相色谱法测定[18],氨态氮采用苯酚-次氯酸钠比色法测定[27];微生物数量采用平板计数法计数,乳酸菌、霉菌、酵母菌以及好氧细菌分别用MRS培养基、高盐察氏培养基、麦芽糖浸粉琼脂培养基和营养琼脂培养基进行培养。有氧稳定性测定在发酵60 d后打开全部实验室青贮发窖袋,袋口用双层纱布包裹,防止果蝇等其他杂质污染和水分散失,空气可自由进入发酵袋中,置于室温条件下保存。将多点式温度记录仪(i500-E3TW,玉环智拓仪器科技有限公司)的多个探头分别放置于发酵袋的几何中心,同时在环境中放置3个探头,用于测定环境温度,温度记录仪测量时间间隔设置为5 min,每个处理放置 3个温度探头。如果样品温度高于环境温度2 ℃,说明青贮开始腐败变质,此时记录时间即为有氧稳定时间。CO2的测定采用CO2产气装置进行测定。CO2产气装置:按Ashbell等[28]的描述,自制评定有氧稳定性的CO2产气装置。每套装置包括2个500 mL 矿泉水塑料瓶,分上(放置25 g样品)下(KOH溶液)2部分。取20 mL 20% KOH溶液,放入上述产气装置内,置于30 ℃的恒温培养箱中进行CO2产气量测定。
1.4 数据处理
在Excel中作数据的基本处理,用SPSS 20.0对各处理的pH值、WSC、NH3-N、乳酸、乙酸、丙酸、丁酸含量、CO2、乳酸菌、霉菌、酵母菌以及好氧细菌数量以及氧稳定时间进行方差分析,通过Duncan法对各处理间的差异进行比较,并对上述指标进行具有互作效应的双因素方差分析。采用Origin 9.0进行绘图,对开窖时间和各微生物数量进行拟合分析。应用 Fzuuy数学中隶属函数法进行青贮发酵品质以及微生物数量的综合评判。其中,与发酵品质呈正相关的指标(WSC、乳酸、乙酸、乳酸菌、有氧稳定性)采用公式(1)计算,与发酵品质呈负相关的指标(pH值、NH3-N、NH3-N/TN、霉菌、酵母菌、好氧细菌和CO2含量)采用公式(2)计算。

式中Fij为第i个处理第j个指标的隶属度;Xij为第i个处理第 j个指标测定值;Xmax、Xmin为所有供试对象中第 j项指标的最大值、最小值。
2 结果与分析
2.1 添加不同乳酸菌有氧稳定性变化及CO2产气分析
玉米青贮中添加乳酸菌处理的有氧稳定性如图 1所示,各处理有氧暴露后稳定的时间分别为CK:131.71 h,T:131.54 h,Y:195.58 h,TY:196.21 h。其中Y和TY处理有氧稳定时间显著高于其余处理 63.87~64.67 h(P<0.05),且二者差异不显著(P>0.05)。同时CK和T处理间差异不显著(P>0.05)。
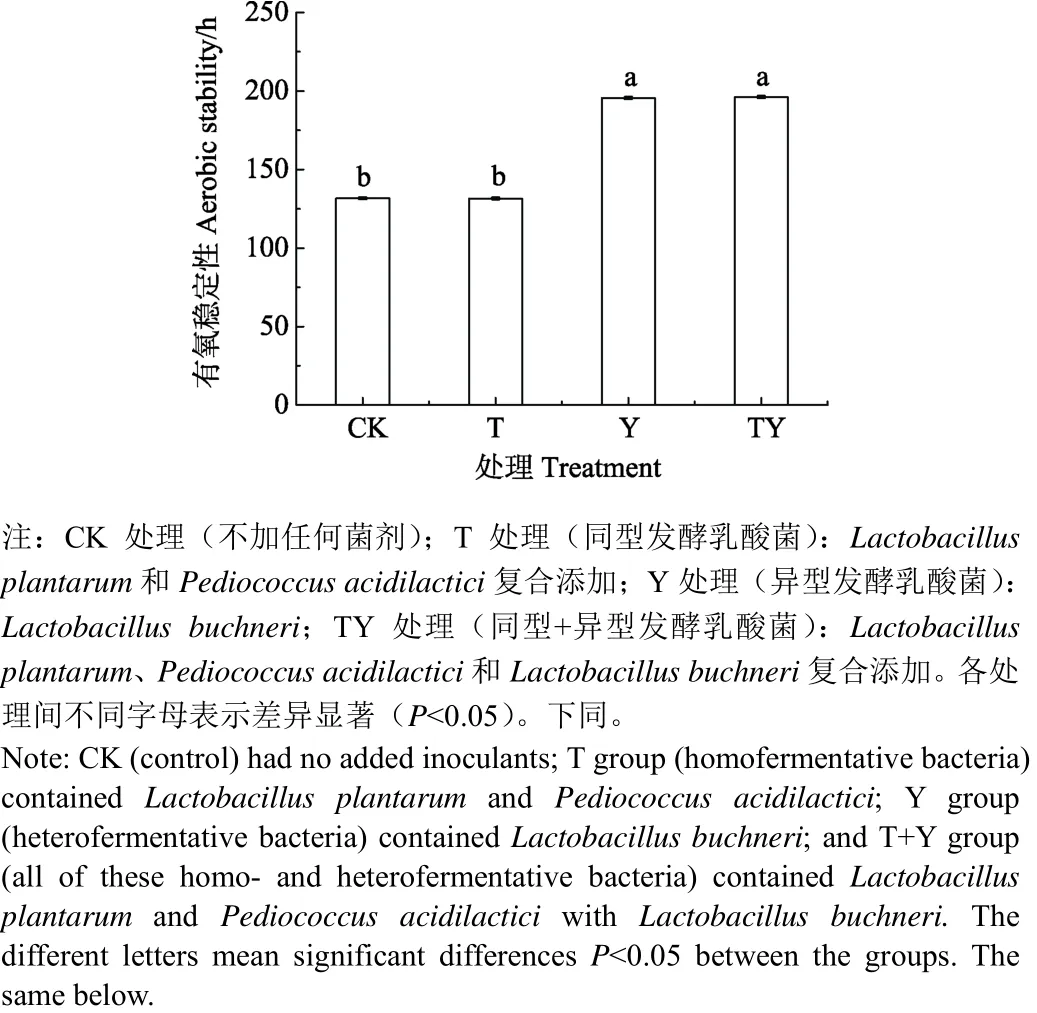
图1 不同处理玉米青贮的有氧稳定性Fig. 1 Aerobic stability of different treatments corn silage
开窖后,对各处理青贮样品进行CO2产气分析(图2),发现各处理的CO2产量和开窖时间满足Allometric1非线性关系(P<0.01),其关系式为(3)。
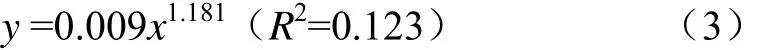
试验结果表明,随着开窖时间的延长,各处理的CO2产气量逐步增加。对比各处理的产气量发现,在开窖全程 CK处理的 CO2产量始终显著高于 T处理(1.8%~87.2%)、Y处理(33.7%~88.6%)和TY处理(9.9%~88.3%)(P<0.05)。除开窖240 h时添加乳酸菌的处理间差异不显著(P>0.05)外,其余开窖过程中Y和 TY处理的CO2产量始终显著低于T处理(P<0.05),且Y处理显著低于TY处理。即玉米青贮中乳酸菌的添加有助于降低在开窖过程中玉米青贮中的 CO2浓度,同时异型发酵乳酸菌(布氏乳杆菌)的添加更易于减少CO2的产生。
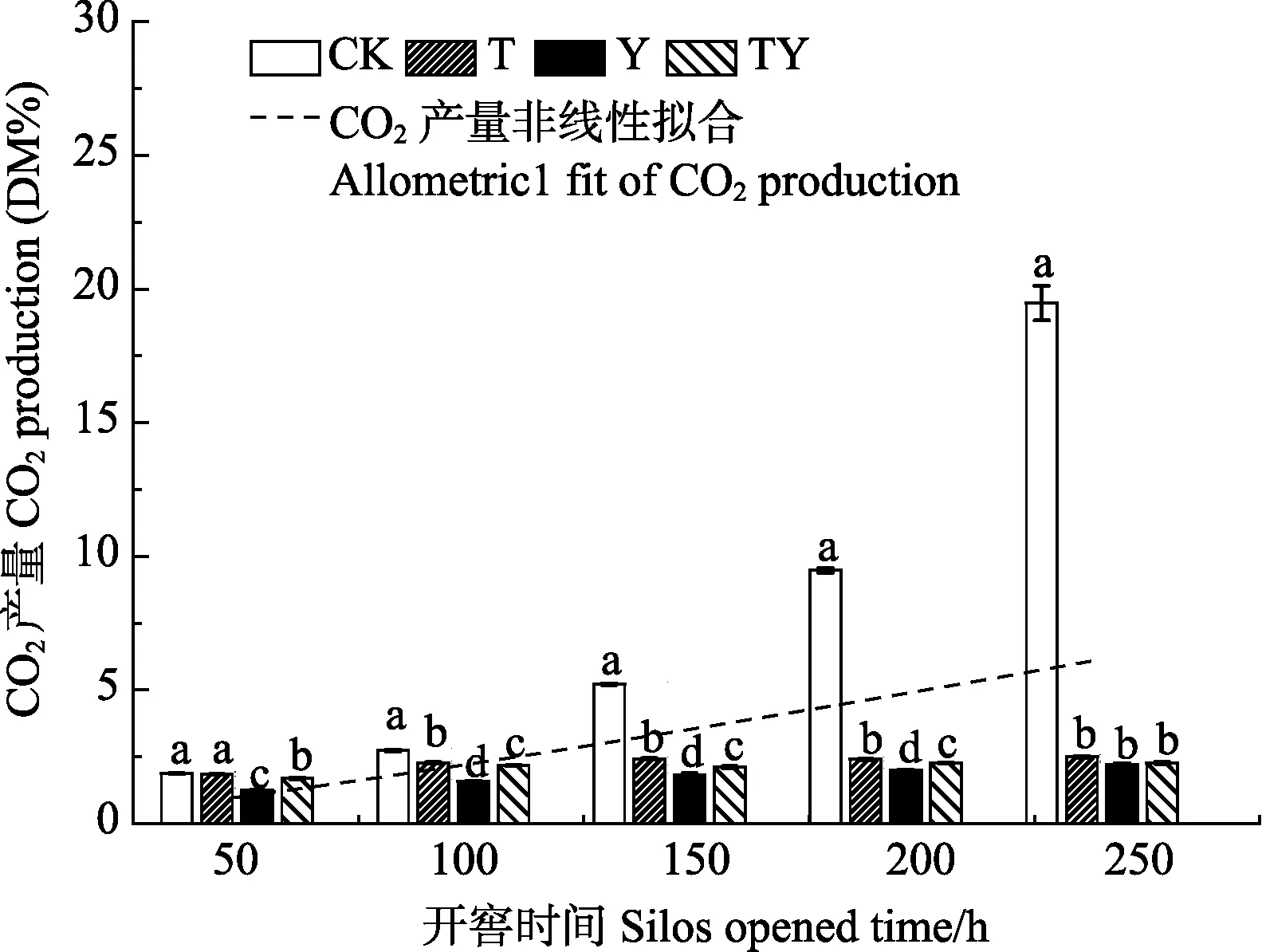
图2 开窖后添加不同乳酸菌玉米青贮CO2产量变化Fig. 2 Change of CO2 production of corn silage adding different lactic acid bacteria after silos opened
2.2 添加不同乳酸菌处理开窖后发酵品质动态变化
玉米青贮开窖后,各处理pH值逐渐增加(表2),且在开窖240 h时较开窖0 h时上升了9.1%~15.1%。开窖后CK处理pH值始终显著高于其余处理3.5%~13.2%(P<0.05),且在开窖240 h后CK处理的pH值超过劣质青贮的规定值4.80。而其余pH值均属于青贮品质一般的pH值范围,T处理pH值始终显著低于CK处理(6.0%~15.2%)、Y处理(2.4%~5.6%)和 TY处理(0.5%~3.9%)(P<0.05)。Y 处理的 pH 值仅在开窖96,144和192 h显著高于TY处理(P<0.05),在开窖结束时二者差异不显著(P>0.05)。由上可知,添加乳酸菌处理在开窖后pH值均较未添加处理低,即玉米青贮中乳酸菌的添加有助于抑制开窖后青贮饲料pH值的升高。
各玉米青贮处理 WSC质量分数在开窖后的变化见表2,开窖240 h时各处理WSC质量分数均较开窖0 h时减少了31.4%~45.4%,即玉米青贮开窖后各处理WSC质量分数逐渐在减少,尤其CK 处理在开窖0 h时WSC质量分数显著高于其余处理11.1%~11.8%(P<0.05),但开窖0 h之后其WSC质量分数迅速降低,直至开窖144 h时显著低于其余处理(P<0.05)。由表2可知,玉米青贮开窖0 h之后,Y处理WSC质量分数始终显著高于CK处理(10.3%~23.9%)、T处理(3.5%~16.2%)和TY处理(9.7%~24.5%)(P<0.05),即单一添加异型发酵乳酸菌(布氏乳杆菌)可在开窖后减缓玉米青贮中WSC质量分数的减少。
开窖后,各玉米青贮处理乳酸的质量分数随开窖时间的延长而减少,开窖240 h时各玉米青贮乳酸质量分数处理较开窖0 h时降低了52.2%~69.5%。开窖后T处理乳酸质量分数始终显著高于CK处理(2.4%~46.5%)和Y处理(8.3%~31.5%)(P<0.05),同时除开窖96 h外显著高于 TY处理 3.8%~25.3%(P<0.05)。除开窖48和96 h外,其余开窖时间CK处理的乳酸质量分数始终显著低于其余处理(P<0.05)。而 Y处理在开窖过程中乳酸质量分数始终显著低于 TY处理 5.0%~9.6%(P<0.05)。由上可知,玉米青贮中乳酸菌的添加可以降低开窖后乳酸的减少量,尤其是添加同型发酵乳酸菌效果较为明显。
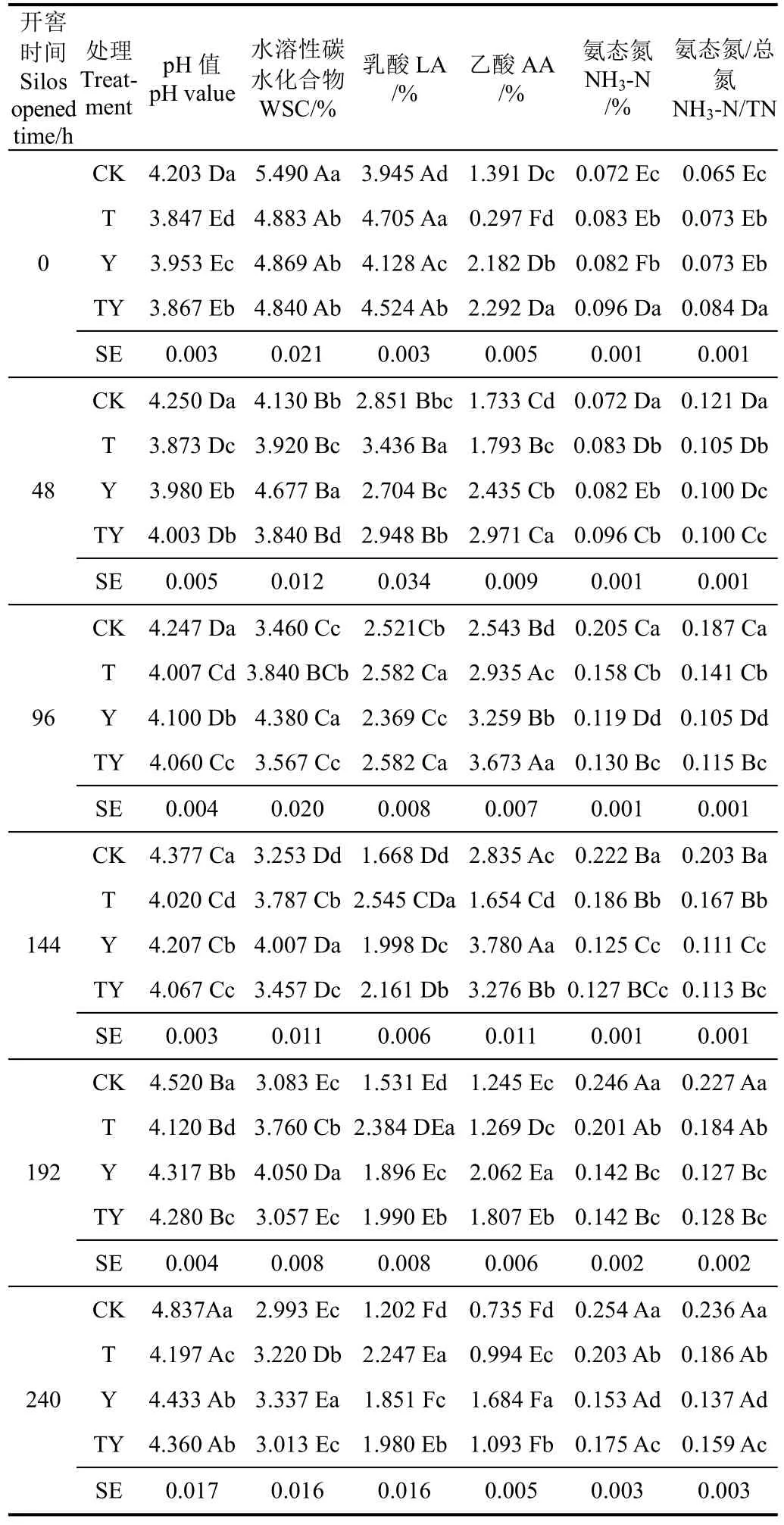
表2 开窖后添加不同乳酸菌玉米青贮发酵品质变化Table 2 Change of fermentation characteristics of corn silage adding different lactic acid bacteria after silos opened
由表 2可知,玉米青贮开窖后,各处理的乙酸质量分数均出现一定程度的上升趋势(T和TY0~96 h,CK和Y0~144 h),之后逐渐下降,除T处理外,其余处理在240 h的乙酸质量分数均较开窖0 h减少22.9%~52.3%。其中开窖后Y和TY处理乙酸质量分数始终显著高于CK和T处理(P<0.05)。而在开窖0~96 h,TY处理乙酸质量分数始终显著高于 Y处理(P<0.05),此后直至开窖结束时始终显著低于 Y处理(P<0.05)。由此可知,玉米青贮中添加含有异型发酵乳酸菌(布氏乳杆菌)的处理均对开窖后抑制乙酸的损失具有积极作用。丙酸和丁酸质量分数较低,低于检出线。
玉米青贮开窖后,各处理NH3-N质量分数迅速增加,开窖后240 h较开窖0 h时增加了82.8%~251.5 %(表2)。其中CK处理的NH3-N质量分数增加量最大,达251.5%,且开窖0 h之后的有氧暴露过程中始终显著高于T处理(11.1%~23.1%)、Y处理(14.9%~43.7%)和TY处理(15.0%~42.7%)(P<0.05)。Y和TY处理 NH3-N质量分数除开窖0和48 h外,其余时间均显著低于T处理(P<0.05),但仅在开窖0,96和240 h,Y处理NH3-N质量分数显著高于TY处理(P<0.05)。同时NH3-N/TN的变化与NH3-N质量分数变化极为相似。因此,玉米青贮添加乳酸菌后在开窖过程中可减少NH3-N的产生,且异型发酵乳酸菌(布氏乳杆菌)的添加更易于减少NH3-N的产生。
2.3 添加不同乳酸菌处理开窖后微生物数量动态变化
各玉米青贮处理中开窖后 240 h乳酸菌数量显著高于开窖0 h时的10.7%~16.1%(P<0.05,图3a),其中添加乳酸菌处理的玉米青贮中 240 h乳酸菌数量极显著高于开窖0 h时(P<0.01)。将各处理的乳酸菌数量与开窖时间进行拟合发现满足ExpDec1非线性关系(P<0.01),其关系式为式(4)。
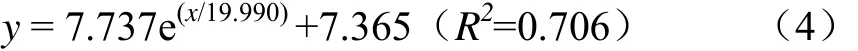
试验结果表明,随着开窖时间的延长,各处理的乳酸菌数量逐步增加。同时,T和TY处理乳酸菌数量除开窖0和48 h外其余时间均显著高于CK处理(P<0.05),即玉米青贮中乳酸菌的添加有助于增加开窖过程中玉米青贮中乳酸菌数量。
开窖后,对各处理青贮样品进行霉菌数量分析(图3b),将各处理的霉菌数量和开窖时间进行拟合发现满足一元线性关系(P<0.01),其关系式见式(5)。

随着开窖时间的延长,各处理的霉菌数量逐步增加。同时发现各处理开窖后240 h霉菌数量著高于开窖0 h时2.7%~21.7%(P<0.05),其中添加乳酸菌处理的玉米青贮中240 h霉菌数量极显著高于开窖0 h时(P<0.01)。Logistic开窖后Y处理的霉菌数量始终显著低于CK处理(4.6%~31.3%)、T处理(24.7%~34.4%)和 TY处理(31.0%~45.2%)(P<0.05)。而 T和 TY处理的霉菌数量在开窖后始终显著高于其余处理(P<0.05),即单一添加异型发酵乳酸菌(布氏乳杆菌)可在开窖后抑制玉米青贮中霉菌的增长,但添加同型发酵乳酸菌的效果不明显,且复合添加 2种发酵类型的乳酸菌反而使开窖后霉菌数量增加。
玉米青贮开窖后,各处理酵母菌数量变化如图3c所示,开窖后240 h酵母菌数量均极显著高于开窖0 h时117.7%~206.3%(P<0.01),并从各处理的酵母菌数量与开窖时间拟合分析中发现二者呈显著的 Logistic非线性关系(P<0.01),且关系式为式(6)。

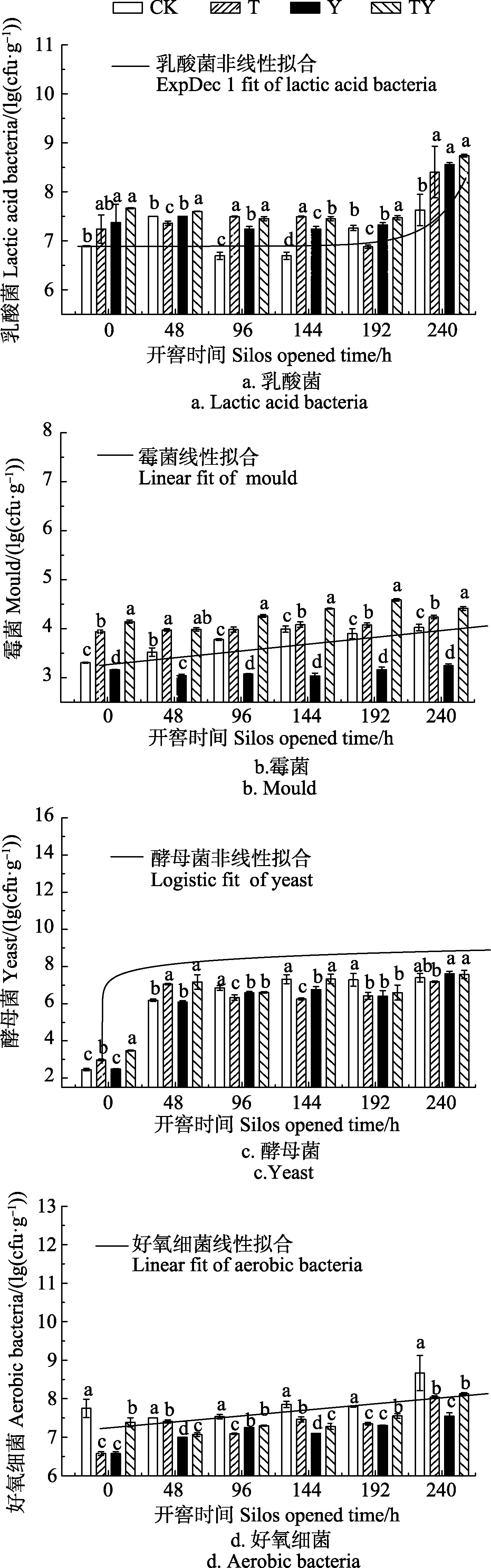
图3 开窖后添加不同乳酸菌玉米青贮微生物数量变化Fig. 3 Change of microbial populations of corn silage at adding different lactic acid bacteria after silos opened
随着开窖时间的延长,各处理的酵母菌数量逐步增加。除48和192 h外,其余开窖时间T处理酵母菌数量始终显著低于TY处理1.7%~17.4%(P<0.05)。而CK处理的酵母菌数量在 96~192 h显著高于 T和 Y处理(P<0.05)。即添加2种同型发酵乳酸菌或单一添加异型发酵乳酸菌(布氏乳杆菌)可在开窖后大部分时间内抑制玉米青贮中酵母菌的增长,但复合添加 2种发酵类型的乳酸菌反而效果不佳。
各处理玉米青贮开窖 240 h好氧细菌数量均极显著高于开窖0 h时10.0%~22.2%(P<0.01,图3d),将各处理的好氧细菌数量和开窖时间进行拟合发现二者满足一元线性关系,其关系式为式(7)。
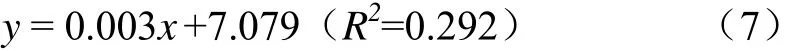
随着开窖时间的延长,各处理的好氧细菌数量逐步增加。CK处理好氧细菌数量始终显著高于 T处理(1.2%~15.1%)、Y处理(3.8%~15.1%)和 TY处理(3.1%~6.2%)(P<0.05)。同时,Y 处理好氧细菌数量除开窖96 h外均显著低于TY处理(P<0.05),并在开窖第48、144和240 h显著低于T处理(P<0.05)。由此可知,玉米青贮中添加乳酸菌有助于降低开窖过程的好氧细菌数量,同时异型发酵乳酸菌(布氏乳杆菌)的单一添加更易于减少好氧细菌的产生。
2.4 开窖时间及添加不同乳酸菌处理对开窖后玉米青贮品质的交互作用
开窖时间及试验设置的添加不同乳酸菌处理均对玉米青贮发酵品质和微生物数量产生极显著影响(P<0.01,表3),通过双因素方差分析可知开窖时间与处理的交互作用对上述所有指标均产生极显著影响(P<0.01)。
2.5 添加不同乳酸菌处理开窖后玉米青贮发酵品质及微生物数量的综合评价
采用模糊数学隶属函数值法,以pH值、WSC、乳酸、乙酸、NH3-N、NH3-N/TN、CO2产气量、有氧稳定性、乳酸菌、霉菌、酵母菌和好氧细菌数量12个指标,对添加不同乳酸菌处理的玉米青贮发酵品质及微生物数量进行综合评价(表4)。综合评价分值越高则青贮品质越好,反之越差。开窖240 h后,对添加不同乳酸菌处理的玉米青贮发酵特性以及微生物数量的综合排序为:
Y处理>TY处理>T处理>CK处理。

表4 开窖后添加不同乳酸菌玉米青贮发酵品质及微生物数量隶属函数分析Table 4 Analysis of membership function of fermentation characteristics and microbial quantity of corn silage adding different lactic acid bacteria after silos opened
3 讨 论
3.1 添加不同乳酸菌对玉米青贮有氧稳定性及 CO2产量的影响
青贮饲料在青贮容器开封后的稳定程度称之为青贮饲料的有氧稳定性。在生产实践中,有氧稳定性代表青贮饲料对温度升高的抵抗力,而对青贮温度的监测是开窖后最常见的物料稳定性的指标[9]。本研究中各处理的CO2产量随着开窖时间逐渐增加,因为青贮开窖后,厌氧环境立即转变为有氧环境,其中好氧微生物的活动性增强,其利用青贮发酵底物产生水、CO2和热量[29],直接导致青贮温度上升。但添加乳酸菌有助于降低开窖过程中玉米青贮中的 CO2浓度。因为青贮主要依靠乳酸菌进行厌氧发酵产生乳酸,降低 pH从而抑制不良微生物生长,减少营养损失,进一步减少 CO2的产生[21]。同时本研究发现玉米青贮中添加乳酸菌可适当提高有氧稳定性,这与Jungbluth等[30]的研究结果一致,Jungbluth通过生物添加剂成功地抑制了青贮饲料的好样腐败,同时减少 CO2产量并提高有氧稳定性。本研究中异型发酵乳酸菌(布氏乳杆菌)的添加更易于提高有氧稳定性,减少CO2的产生,因为异型发酵的布氏乳杆菌可产生更多的乙酸,抑制好氧变质,延长有氧稳定时间,而在有氧暴露期间,同型发酵乳酸菌所产生的乳酸可作为酵母菌生长的底物,仅产生少量能抑制酵母菌和霉菌生长的短链脂肪酸(乙酸、丙酸、丁酸)[29]。这一结果也从前人研究中得到了大量验证[15,31-32],因此,单一接种同型发酵乳酸菌的玉米青贮有氧稳定性较差。另一方面,从本研究结果中可知同型发酵乳酸菌和异型发酵乳酸菌的复合添加的处理有氧稳定时间最长,显著高于对照组和单一接种同型发酵乳酸菌的玉米青贮处理(P<0.05),但与单一接种异型发酵乳酸菌的玉米青贮有氧稳定性无显著差异(P>0.05),这与Filya[33]报道较为相似。Filya用布氏乳杆菌和植物乳杆菌联合接种高粱与玉米,与单独使用布氏乳杆菌相比,得到折中的实验结果,而联合接种的青贮饲料比未接种的具有更好的有氧稳定性。
3.2 添加不同乳酸菌对开窖后玉米青贮发酵品质的影响
青贮饲料暴露在空气中,pH值的增加是青贮饲料变质的一个重要指标[9]。因为在青贮开窖后便进入有氧发酵阶段,此时大量以有机酸(乳酸、可溶性糖和乙醇)为主要营养基质维持生存的微生物大量生长。而乳酸在降低和维持低pH值青贮饲料中起着极为重要的作用[34]。当大量以酵母菌为主的好氧微生物接触到空气时,消耗乳酸引起pH值的增加。因此,本研究中无论是否添加乳酸菌的处理pH均随开窖时间的延长逐渐增加。但是添加乳酸菌处理在开窖后pH值均较未添加处理低。这与前人研究结果一致[30-31],由于同型发酵乳酸菌在发酵过程中产生较多乳酸,开窖后虽然有所损失,仍然较其余处理质量分数高,可以维持较低pH环境[35]。同时,异型发酵乳酸菌在开窖后可分解乳酸产生乙酸,在一定程度上维持了玉米青贮中低pH水平[36]。
本研究发现,各处理的WSC质量分数均逐渐减少。因为在开窖初期细胞呼吸作用,以及随开窖时间的增加青贮温度的升高促进酶的水解,可能引起部分WSC的分解,造成不消化成分质量分数的相对增加,其中,WSC发酵转化为 CO2[37]。但是单一添加异型发酵布氏乳杆菌可在开窖后减缓玉米青贮中WSC质量分数的减少。因为,异型发酵布氏乳杆菌产生乙酸抑制好氧微生物增殖,降低WSC的消耗。而异型发酵乳酸菌除利用WSC产生乙酸外,还可以利用乳酸产生乙酸和少量的1,2-丙二醇[38],即损失的WSC质量分数较少,乳酸质量分数低,但高于对照组。本研究中添加同型发酵乳酸菌的玉米青贮处理开窖后乳酸质量分数较高。因为在开窖初期氧气质量分数较少,少量的氧气刺激同型发酵乳酸菌利用青贮饲料中的WSC产生乳酸以维持低pH环境防止有氧腐败,与Wang等[27]的研究结果相似。Romero的研究同样表明添加布氏乳杆菌的青贮饲料在增加乙酸生产的同时,其乳酸浓度较未添加任何乳酸菌的对照组高[36]。Kleinschmit等[39]将全株玉米青贮接种布氏乳杆菌和片球菌,在51~361 d中比未接种组得到更高浓度的乙酸和1,2-丙二醇。从本研究中也发现同型发酵乳酸菌和异型发酵乳酸菌的复合添加的处理具有较高的乳酸和乙酸质量分数。其原因是该处理的WSC被大量消耗,主要被同型发酵乳酸菌用以产生乳酸,而其中一部分被异型发酵乳酸菌用以产生乙酸,同时异型发酵乳酸菌还利用乳酸产生乙酸。
NH3-N浓度主要反映蛋白质的降解程度,是评价青贮饲料品质的一个重要指标。玉米青贮开窖后,pH值逐渐上升,当pH值>4.2时不良微生物的活动,氨基酸被分解成氨、硫化氢和胺类,降解率增大,使得NH3-N质量分数增加[40]。本研究发现玉米青贮添加乳酸菌后在开窖过程中可减少NH3-N的产生。Junior等[9]研究发现添加乳酸菌处理相比对照处理,能有效地降低青贮饲料中蛋白质的降解。Wang等[27]的研究中也得到了相应的证实,该研究通过设置玉米和苜蓿混合青贮中添加植物乳杆菌,在开窖后发现青贮饲料的发酵特性提高且保持低pH值,具有较高乳酸质量分数,同时降低氨态氮浓度。而本研究结果显示异型发酵乳酸菌(布氏乳杆菌)的添加更易于减少NH3-N的产生,归咎于异型发酵乳酸菌所产生的乙酸抑制好氧微生物的活动,减缓对氨基酸的分解[11]。由于异型发酵的乳酸菌的作用,在复合添加乳酸菌的处理中NH3-N质量分数较低。
3.3 添加不同乳酸菌对开窖后玉米青贮微生物数量的影响
青贮饲料发酵过程中乳酸菌数量占主导地位,使好氧微生物的活动受到限制[15]。本研究发现玉米青贮中乳酸菌的添加有助于增加开窖过程中玉米青贮中乳酸菌数量。因为开窖后同型和异型发酵乳酸菌利用WSC等营养物质,维持生长繁殖,由于添加的乳酸菌数量在青贮中占主导地位,因此即使开窖后有氧环境不适于乳酸菌继续生长,但其数量仍然缓慢增加。Tran等[38]研究发现青贮饲料中固有的布氏乳杆菌暴露于空气后,其活性不足以维持青贮饲料的安全稳定。而布氏乳杆菌添加剂的使用可进一步促进乙酸产生,从而抑制好氧微生物的生长,降低霉菌数量。本研究中单一添加异型发酵乳酸菌(布氏乳杆菌)可在开窖后抑制玉米青贮中霉菌的增长,但添加同型发酵乳酸菌的效果不明显,且复合添加2种发酵类型的乳酸菌反而使开窖后霉菌数量增加。可能由于开窖后玉米青贮中的厌氧环境被打破,且单一添加同型发酵乳酸菌及复合添加2种发酵类型乳酸菌的玉米青贮中含有较高的乳酸,为霉菌的生长繁殖创造了有利条件,虽然含有部分乙酸但其对霉菌的抑制作用弱于霉菌数量的增长。
Jungbluth等[30]研究发现微生物开始改变它们的新陈代谢,并利用氧作为对厌氧阶段的直接反应。一旦它们的新陈代谢从厌氧变成有氧,氧气被微生物呼吸所消耗,伴随着温度上升。试验过程中各处理在开窖后好氧细菌均有不同程度的增加,青贮pH环境的变化促使好氧微生物生长,造成青贮营养成分的进一步的损失[41]。本研究中添加乳酸菌的玉米青贮中好氧细菌数量较对照组少,同时异型发酵乳酸菌的单一添加更易于减少好氧细菌的产生。这些抗菌性能是通过有机酸穿过细胞膜在细胞质内解离并释放H+的结果。其结果使细胞内的pH迅速降低,产生大量ATP抵抗pH值下降以维持细胞内稳态,最终导致细胞减少或停止生长[27]。在Kharazian等[4]对生物测定的结果中反映出,异型发酵乳酸菌相对于同型发酵菌具备较高抑菌性能。本研究发现单一添加同型或异型发酵乳酸菌可在开窖后大部分时间内抑制玉米青贮中酵母菌的增长,但复合添加 2种发酵类型的乳酸菌反而效果不佳。是由于开窖后复合添加同型或异型发酵乳酸菌的玉米青贮中,其pH值高于添加同型发酵乳酸菌的玉米青贮处理,乙酸质量分数低于单一添加异型发酵乳酸菌的玉米青贮中的玉米青贮处理。
Baek等[42]报道,明串珠菌和魏斯氏乳酸菌所产生的乳酸和乙酸的是主要抗菌物质。已有的研究表明乳酸在抑菌中起重要作用,但不是唯一的一种。在 Kharazian等[4]的研究中,一些乳酸菌菌株没有高质量分数的乳酸生产但显示出较高的抑菌率,因此乳酸菌可能会产生一些其他的抗真菌物质。异型发酵乳酸菌能抑制好氧变质,其产生的乙酸被看作是主要的抑制物质[18]。但Akihisa等[43]研究发现添加异型发酵乳酸菌 TM2的黑麦草青贮饲料发酵品质好,乙酸和丙酸质量分数也较少但有氧稳定性比乙酸质量分数较多的乙酸添加青贮饲料好,说明乳酸菌TM2能产生乙酸以外的抗酵母菌和霉菌的物质。最近的研究已经证实,一些乳酸菌生产其他抗真菌化合物,如环二肽、蛋白质类化合物、有机酸、脂肪酸和抑菌素[44-45]。因此,添加这类不仅能提高青贮发酵品质,而且能改善青贮饲料有氧稳定性的乳酸菌值得研究与利用。
4 结 论
综上所述,玉米青贮中添加不同乳酸菌对开窖后青贮饲料发酵品质、微生物数量、有氧稳定性影响显著。综合评价表明,单一添加异型发酵乳酸菌(Lactobacillus buchneri,添加量为 1×105cfu/g)的处理评分最高(0.790)。因此,单一添加异型发酵乳酸菌的玉米青贮,开窖后在改善发酵品质、降低有害微生物数量、提高有氧稳定性方面均优于单一添加同型或复合添加乳酸菌的玉米青贮。
[1] Ferreira G, Alfonso M, Depino S, et al. Effect of planting density on nutritional quality of green-chopped corn for silage[J]. Journal of Dairy Science, 2014, 97(9): 5918-5921.
[2] Neumann M, Horts E H, Figueira D N, et al. Potential of corn silage production in different sowing times in the Paraná Midwest region[J]. 2016, 9(1): 37-44.
[3] Keller L A M, Pereyra M L G, Keller K M, et al. Fungal and mycotoxins contamination in corn silage: monitoring risk before and after fermentation[J]. Journal of Stored Products Research, 2013, 52(1): 42-47.
[4] Kharazian Z A, Jouzani G S, Aghdasi M, et al. Biocontrol potential of Lactobacillus, strains isolated from corn silages against some plant pathogenic fungi[J]. Biological Control,2017, 110(4): 33-43.
[5] Yuan X J, Guo G, Wen A Y, et al. The effect of different additives on the fermentation quality, in vitro, digestibility and aerobic stability of a total mixed ration silage[J]. Animal Feed Science & Technology, 2015, 207(6): 41-50.
[6] Ni K, Wang Y, Li D, et al. Characterization, identification and application of lactic acid bacteria isolated from forage paddy rice silage[J]. Plos One, 2015, 10(3): 1-14.
[7] Jr K L, Robinson J R, Ranjit N K, et al. Microbial populations, fermentation end-products, and aerobic stability of corn silage treated with ammonia or a propionic acid-based preservative.[J]. Journal of Dairy Science, 2000, 83(7): 1479-1486.
[8] Muck R E. Silage microbiology and its control through additives.[J]. Revista Brasileira De Zootecnia, 2010, 39(39):183-191.
[9] Junior V H B, Fortaleza A P D S, Junior F L M, et al.Aerobic stability in corn silage (Zea mays L.) ensiled with different microbial additives[J]. Acta Scientiarum Animal Sciences, 2017, 39(4): 357-362.
[10] Rabelo C H S, Rezende A V D, Nogueira D A, et al. Perdas fermentativas e estabilidade aeróbia de silagens de milho inoculadas com bactérias ácido-láticas em diferentes estádios de maturidade[J]. Revista Brasileira De Saúde E Produção Animal, 2012, 13(3): 656-668.
[11] Silva N C D, Dos Santos J P, Ávila, Carla L. S, et al.Evaluation of the effects of two Lactobacillus buchneri strains and sodium benzoate on the characteristics of corn silage in a hot-climate environment[J]. Grassland Science,2015, 60(3): 169-177.
[12] Denek N, Can A, Avci M, et al. The effect of molasses-based pre-fermented juice on the fermentation quality of first-cut lucerne silage[J]. Grass & Forage Science, 2011, 66(2): 243-250.
[13] Chen L, Guo G, Yuan X, et al. Effects of applying molasses,lactic acid bacteria and propionic acid on fermentation quality, aerobic stability and in vitro gas production of total mixed ration silage prepared with oat–common vetch intercrop on the Tibetan Plateau[J]. Journal of the Science of Food & Agriculture, 2016, 96(5): 1678-1685.
[14] Trabi E B, Yuan X, Li J, et al. Effect of glucose and lactic acid bacteria on the fermentation quality, chemical compositions and in vitro digestibility of mulberry (Morus Alba) leaf silage[J]. Pakistan Journal of Zoology, 2017, 49(6):2271-2277.
[15] Koc F, Aksoy S O, Okur A A, et al. Effect of pre-fermentated juice, Lactobacillus plantarum and Lactobacillus buchneri on the fermentation characteristics and aerobic stability of high dry matter alfalfa bale silage[J]. Journal of Animal & Plant Sciences, 2017, 27(5): 1426-1431.
[16] Liu Q H, Shao T, Bai Y F. The effect of fibrolytic enzyme;Lactobacillus plantarum, and two food antioxidants on the fermentation quality; alpha-tocopherol and beta-carotene of high moisture napier grass silage ensiled at different temperatures[J]. Animal Feed Science & Technology, 2016,221(8): 1-11.
[17] Dunière L, Sindou J, Chaucheyras-Durand F, et al. Silage processing and strategies to prevent persistence of undesirable microorganisms[J]. Animal Feed Science &Technology, 2013, 182(1-4): 1-15.
[18] Rabelo C H S, Basso F C, Lara E C, et al. Effects of Lactobacillus buchneri as a silage inoculant and as a probiotic on feed intake, apparent digestibility and ruminal fermentation and microbiology in wethers fed low-dry-matter whole-crop maize silage[J]. Grass & Forage Science, 2018,73(1): 534-544.
[19] Rabelo C, Basso F, Mcallister T A, et al. Influence of Lactobacillus buchneri and forage: Concentrate ratio on the growth performance, fatty acid profile in longissimus muscle and meat quality of beef cattle[J]. Canadian Journal of Animal Science, 2016, 96(4): 550-562.
[20] Li X, Xu W, Yang J, et al. Effects of applying lactic acid bacteria to the fermentation on a mixture of corn steep liquor and air-dried rice straw[J]. Animal Nutrition, 2016, 2(3): 229-233.
[21] Abdul R N, Abd Halim M R, Mahawi N, et al. Determination of the use of Lactobacillus plantarum and Propionibacterium freudenreichii application on fermentation profile and chemical composition of corn silage[J]. BioMed Research International, 2017(1): 1155-1162.
[22] Huisden C M, Adesogan A T, Kim S C, et al. Effect of applying molasses or inoculants containing homofermentative or heterofermentative bacteria at two rates on the fermentation and aerobic stability of corn silage[J].Journal of Dairy Science, 2009, 92(2): 690-697.
[23] Weinberg Z G, Ashbell G, Hen Y, et al. Ensiling whole-crop wheat and corn in large containers with Lactobacillus plantarum and Lactobacillus buchneri[J]. Journal of Industrial Microbiology & Biotechnology, 2002, 28(1): 7-11.
[24] Adesogan A T, Salawu M B. Effect of applying formic acid,heterolactic bacteria or homolactic and heterolactic bacteria on the fermentation of bi-crops of peas and wheat[J]. Journal of the Science of Food & Agriculture, 2004, 84(9): 983-992.
[25] 张丽英. 饲料分析及饲料质量检测技术(第 2版)[M]. 北京:中国农业出版社, 2002.
[26] 高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006.Gao Junfeng. Plant Physiology Experimental Guidance [M].Beijing: Higher Education Press, 2006. (in Chinese with English abstract)
[27] Wang M, Yu Z, Wu Z, et al. Effect of Lactobacillus plantarum‘KR107070’ and a propionic acid-based preservative on the fermentation characteristics, nutritive value and aerobic stability of alfalfa-corn mixed silage ensiled with four ratios[J]. Grassland Science, 2017, 64: 51-60.
[28] Ashbell G, Weinberg Z G, Azrieli A, et al. A simple system to study the aerobic deterioration of silages[J]. El Día Médico,1991, 30(64): 391-393.
[29] Wang M, Yang C, Jia L, et al. Effect of Lactobacillus buchneri and Lactobacillus plantarum on the fermentation characteristics and aerobic stability of whipgrass silage in laboratory silos[J]. Grassland Science, 2015, 60(4): 233-239.
[30] Jungbluth K H, Trimborn M, Maack G C, et al. Effects of three different additives and two different bulk densities on maize silage characteristics, temperature profiles, CO2and O2-dynamics in small scale silos during aerobic exposure[J].Applied Sciences, 2017, 7(6): 545-556.
[31] Kristensen N B, Sloth K H, Højberg O, et al. Effects of microbial inoculants on corn silage fermentation, microbial contents, aerobic stability, and milk production under field conditions[J]. Journal of Dairy Science, 2010, 93(8): 3764-3774.
[32] Tabacco E, Piano S, Revellochion A, et al. Effect of Lactobacillus buchneri LN4637 and Lactobacillus buchneri LN40177 on the aerobic stability, fermentation products, and microbial populations of corn silage under farm conditions.[J]. Journal of Dairy Science, 2011, 94(11): 5589-5598.
[33] Filya I. The Effect of Lactobacillus buchneri, and Lactobacillus plantarum, on the fermentation, aerobic stability, and ruminal degradability of low dry matter corn and sorghum silages[J]. Journal of Dairy Science, 2003,86(11): 3575-3581.
[34] Jungbluth K, Maack C, Büscher W, et al. A new ex-situ method to investigate aerobic stability of maize silage faces[J]. Journal of Agricultural Science and Technology.2016, 2, 49-54.
[35] Yuan X J, Wang J, Guo G, et al. Effects of ethanol, molasses and Lactobacillus plantarum on fermentation characteristics and aerobic stability of total mixed ration silages[J]. Grass &Forage Science, 2016, 71(2): 328-338.
[36] Romero J J, Zhao Y, Balseca-Paredes M A, et al. Laboratory silo type and inoculation effects on nutritional composition,fermentation, and bacterial and fungal communities of oat silage[J]. Journal of Dairy Science, 2017, 100(3): 1812-1828.
[37] Ávila C L S, Pinto J C, Sugawara M S, et al. Effect of addition of a bacteria isolated from sugarcane silage on the quality of the silage[J]. Acta Scientiarum Animal Sciences,2008, 30(3): 255-261.
[38] Tran T M T, Nguyen M T, Nguyen H V, et al. Effects of wilting and lactic acid bacteria inoculation on fermentation and microbial community of elephant grass silage produced in Vietnam[J]. Grassland Science, 2017, 10: 1-5.
[39] Kleinschmit D H, Jr K L. A meta-analysis of the effects of Lactobacillus buchneri on the fermentation and aerobic stability of corn and grass and small-grain silages[J]. Journal of Dairy Science, 2006, 89(10): 4005-4013.
[40] Hashemzadeh-Cigari F, Khorvash M, Ghorbani G R, et al.Interactive effects of molasses by homofermentative and heterofermentative inoculants on fermentation quality,nitrogen fractionation, nutritive value and aerobic stability of wilted alfalfa (Medicago sativa L) silage[J]. Journal of Animal Physiology & Animal Nutrition, 2013, 98(2): 290-299.
[41] Borreani G, Piano S, Tabacco E. Aerobic stability of maize silage stored under plastic films with different oxygen permeability. Journal of the Science of Food & Agriculture,2014, 94(13): 2684-2690.
[42] Baek E, Kim H, Choi H, et al. Antifungal activity of Leuconostoc citreum, and Weissella confusa, in rice cakes[J].Journal of Microbiology, 2012, 50(5): 842-848.
[43] Akihisa Y, Hidenori K, Yasuhiro A, et al. Effects of addition of acetic acid, 1,2-propanedol and lactic acid bacteria on the aerobic stability of Italian ryegrass silage[C]// Grassland Agriculture: Balancing Production and Environmental Protection----Proceedings of the, China-Japan-Korea Grassland Conference, 2006: 93-95.
[44] Crowley S, Mahony J, Sinderen D V. Current perspectives on antifungal lactic acid bacteria as natural bio-preservatives[J].Trends in Food Science & Technology, 2013, 33(2): 93-109.
[45] Gajbhiye M H, Kapadnis B P. Antifungal-activity-producing lactic acid bacteria as biocontrol agents in plants[J].Biocontrol Science & Technology, 2016, 26(11): 1451-1470.
猜你喜欢
畜牧兽医科技信息(2022年8期)2022-12-06
野生动物学报(2022年2期)2022-05-16
中学生数理化·高一版(2022年4期)2022-05-09
兽类学报(2022年1期)2022-03-03
中学化学(2021年11期)2021-12-09
电脑迷(2020年10期)2020-12-17
湖南饲料(2019年5期)2019-10-15
今日农业(2018年23期)2018-01-16
中成药(2017年9期)2017-12-19
中国医药指南(2017年30期)2017-01-15
