
温度胁迫对意大利蜜蜂工蜂封盖子能量消耗的影响
2018-06-06 06:05郝振帮徐新建周姝婧周冰峰朱翔杰
福建农林大学学报(自然科学版) 2018年3期
郝振帮, 徐新建, 周姝婧, 周冰峰, 朱翔杰
(福建农林大学蜂学学院,福建 福州 350002)
温度是影响昆虫生命活动最重要的因素之一[1],昆虫的生命过程,如发育、生长、繁殖和衰老,均受到环境温度的制约[2].蜂子(蜜蜂的卵、幼虫、蛹的统称)的发育具有狭温性,工蜂可以通过产热和扇风等行为来调节蜂群的温度[3-5],使蜂子在33~36 ℃发育[6-8],在蜂子分布的中心区域,温度可以维持在(35±0.5)℃范围内[9-11].
封盖子(封盖巢房内蜜蜂的大幼虫、预蛹、蛹)发育对温度变化敏感[12],偏离正常发育温度范围,封盖子的羽化、发育历期,以及羽化后的蜜蜂器官形态和行为等均会受到影响.封盖子的发育温区在29~37 ℃,低于31 ℃或高于37 ℃封盖子的死亡率将显著升高[9,13].低温会引起封盖子发育历期延长,发育温区内,温度越低发育历期越长[9].封盖子偏离正常发育温度会造成羽化蜜蜂翅和足出现畸形[13-14],翅、吻和背板的长度缩短[13],大脑突触的数量降低[9].低温32 ℃培育工蜂封盖子,羽化后学习和短期记忆能力[15]、首次采集日龄、舞蹈行为、清巢行为以及出巢飞行之后回巢的比例均受影响[16].
能量消耗可能是温度胁迫影响蜜蜂发育的因素之一.昆虫在蛹期不取食,变态发育所需能量依赖幼虫期储存的脂肪、糖类和蛋白等能量物质[17].发育温度环境会直接影响到昆虫蛹期发育时间、能量利用,进而影响到蛹的存活和羽化成虫的生殖能力.越冬环境温度升高4 ℃导致蝴蝶(Erynnispropertius)增加了越冬蛹期的能量消耗量[18].角额壁蜂蛹在13 ℃越冬比4 ℃下的脂肪耗速率加快,存活率降低,且羽化成蜂后的寿命缩短[19].低温胁迫对蜜蜂封盖子期能量利用影响的规律尚不清楚.研究温度胁迫对蜜蜂封盖子能量消耗的影响,能够深入了解温度因子对蜜蜂发育影响的机理.通过发育温度对狭温性蜜蜂发育的敏感性的研究,对认识温度与昆虫发育关系的影响规律具有重要的意义.
1 材料与方法
1.1 样本获取与温度处理
选择蜂子健康、蜂王产卵力强的意大利蜜蜂(Apismelliferaligustica)3群[20-21].将蜂王限制在空脾上产卵,取4 h内封盖的工蜂封盖子脾,其日龄记为0 d[22].样本在恒温恒湿箱(±0.1 ℃,CTHI-250B,施都凯仪器设备有限公司)中培育至羽化,温度分别设置为29、31、35、37、38 ℃,相对湿度设置为75%±5%.
1.2 测定指标及方法
分别测定0 d封盖子、不同温度处理1、5、10 d封盖子和刚羽化工蜂的总糖、总脂肪和总蛋白三类物质的含量.使用总糖含量试剂盒(苏州科铭生物技术有限公司)对干燥后的单只封盖子或羽化蜜蜂进行测定.使用BCA试剂盒(北京全式金生物技术有限公司)分别对单只封盖子、刚羽化工蜂总蛋白进行测定.参考索氏抽提法测定脂肪含量[23-24],单只蜜蜂放入离心管中在60 ℃条件下干燥48 h至恒重时称取质量,然后加入1.5 mL氯仿与甲醛的混合液(2∶1,体积比)进行抽提.24 h后更换新的抽提液,重复3次后在60 ℃条件下干燥48 h至恒重时称取质量,脂肪含量即为2次质量的差值.能量物质测定每个温度3~15只·群-1,3群重复.封盖子的能量消耗量为0 d能量含量与对应日龄能量含量的差(mg·只-1).
1.3 数据统计分析
蜜蜂总糖、总脂肪和总蛋白的消耗量是0 d封盖子的能量含量与对应日龄的封盖子能量含量的差值,取各处理组的平均值,并计算标准差SD.所有均值使用SPSS 21软件进行单因素方差分析,多重比较采用SNK法.
2 结果与分析
封盖子在29 ℃低温条件下发育会增加其能量的消耗,而在38 ℃条件下会导致其发育先加快后受阻,最终无法完成发育,造成能量堆积.在31、35、37 ℃条件下,蜜蜂只是发育速度不同,完成发育羽化时的能量消耗相同.
2.1 不同温度下蜜蜂封盖子糖的消耗
发育温度影响蜜蜂封盖子的总糖消耗量.在封盖期低温29 ℃导致工蜂发育迟缓,发育期延长,虽然10 d的总糖消耗较低,但整个发育过程总糖消耗总量是增加的.高温38 ℃能够提高早期封盖子的大幼虫总糖消耗,相对于35、37 ℃条件下发育的封盖子,化蛹后逐渐减少了总糖的消耗.
与35 ℃相比,低温29 ℃条件下,封盖子在1、5、10 d的总糖消耗量显著降低;羽化时即整个封盖期间的总糖消耗量显著增加.低温31 ℃发育到5 和10 d时,总糖消耗量显著降低;在羽化时总糖消耗与35 ℃无差异.35、37 ℃培育的封盖子各个阶段的能量消耗量均无显著影响.与35 ℃相比,38 ℃下1 d封盖子的总糖消耗量显著升高;5 d总糖消耗量与35 ℃差异不显著;10 d时总糖消耗量显著降低(表1).
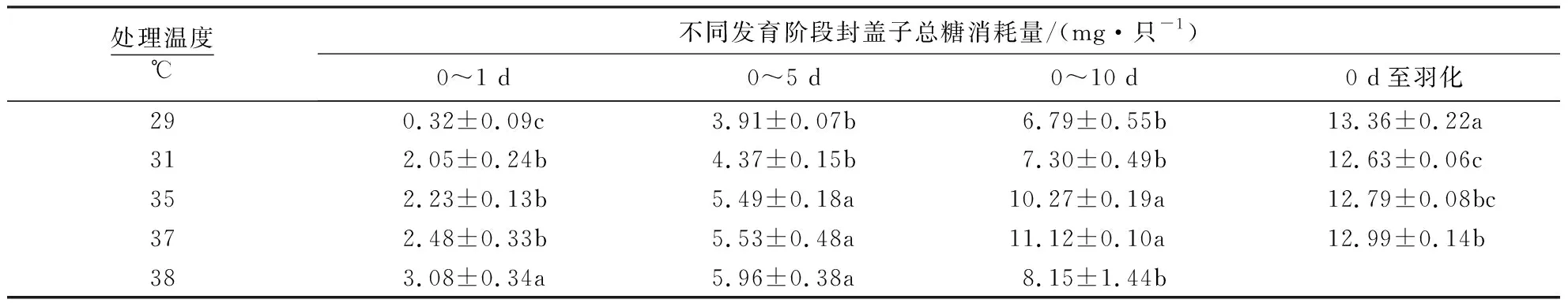
表1 不同发育温度下蜜蜂封盖子总糖消耗1)Table 1 The carbohydrate cost of capped brood honeybee at different temperatures
1)同列不同字母表示不同温度之间差异显著(P<0.05),相同字母表示差异不显著(P>0.05).38 ℃封盖子羽化前全部死亡.
2.2 不同温度下蜜蜂封盖子脂肪的消耗
与35 ℃相比,蜜蜂封盖子的脂肪消耗量仅在38 ℃条件下5和10 d显著降低和29 ℃下1 d显著降低,其余温度条件下蜜蜂封盖子的脂肪消耗量均无显著差异(表2).

表2 不同发育温度下蜜蜂封盖子脂肪消耗1)Table 2 The lipids cost of capped brood honeybee at different temperatures
1)同列不同字母表示不同温度之间差异显著(P<0.05),相同字母表示差异不显著(P>0.05).38 ℃封盖子羽化前全部死亡.
2.3 不同温度下蜜蜂封盖子总蛋白的消耗
发育温度会影响封盖子的总蛋白消耗量.与35 ℃相比,低温29 ℃胁迫封盖子在1、5和10 d的总蛋白消耗量显著降低,但羽化时即整个封盖期间的总蛋白消耗量显著增加.在31、38 ℃胁迫下,封盖子仅在10 d时的蛋白消耗量显著降低.而其他发育阶段与35、37 ℃的总蛋白消耗均无显著差异(表3).
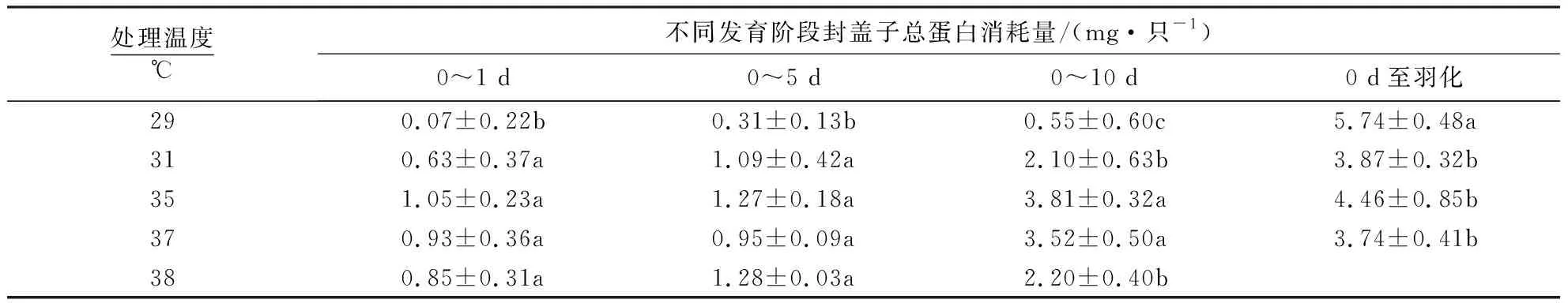
表3 不同发育温度下蜜蜂封盖子总蛋白消耗1)Table 3 The protein cost of capped brood honeybee at different temperatures
1)同列不同字母表示不同温度之间差异显著(P<0.05),相同字母表示差异不显著(P>0.05).38 ℃封盖子羽化前全部死亡.
3 讨论
3.1 低温影响封盖子的能量消耗
低温29 ℃条件下蜜蜂封盖子死亡的主要原因有两个:能量过度消耗和低温对封盖子造成组织损伤.在29 ℃中的蜜蜂封盖子羽化率低于20%,死亡封盖子虫态主要有两种:临近羽化虫态(图1A)和蛹后期(图1B),比例分别为25.22%和58.59%.本研究对这部分封盖子的脂肪含量和总糖含量进行了测定,发现临近羽化虫态的个体脂肪含量和总糖含量均低于羽化蜜蜂.因此这部分蜜蜂死亡可能是由于在低温29 ℃胁迫下的蜜蜂封盖子消耗了过多的能量,造成封盖子在羽化前无法完成发育或无力咬开蜡盖;在蛹后期死亡的封盖子的脂肪含量高于羽化蜜蜂,可能是低温29 ℃造成了封盖子组织损伤而未完成发育.

A:羽化前死亡;B:蛹后期死亡.图1 在29 ℃死亡蜜蜂的状态Fig.1 The two developmental stages of dead honeybee at 29 ℃
3.2 高温影响封盖子发育
较高的发育温度会加快发育速率、降低昆虫的适合度[18,25],但温度升高到发育极限温度时,会阻碍昆虫发育、抑制昆虫能量消耗,造成机体损伤[26-27].前人研究中发现37 ℃会加快封盖子的发育速率、翅和吻长等形态指标变小.本研究中37 ℃可能处于蜜蜂封盖子的耐受范围内,相同处理时间内封盖子能量消耗量和羽化蜜蜂的能量物质含量均与35 ℃无显著差异.封盖子在38 ℃短时间会加速总糖消耗,长时间的高温抑制能量消耗,导致封盖子发育受阻最终死亡.1 d后脂肪消耗受到明显抑制,5 d后蜜蜂封盖子的总糖和总蛋白消耗均受到抑制.高温下封盖子的能量消耗直接影响到封盖子的发育进程.我们观察到5 d后封盖子的虫态慢于35 ℃(图2A,2B),随着38 ℃处理时间的延长封盖子均不能完成发育,死亡在复眼着深褐色状态,身体着色未加深,迟于正常发育的蜜蜂(图2C,2D).
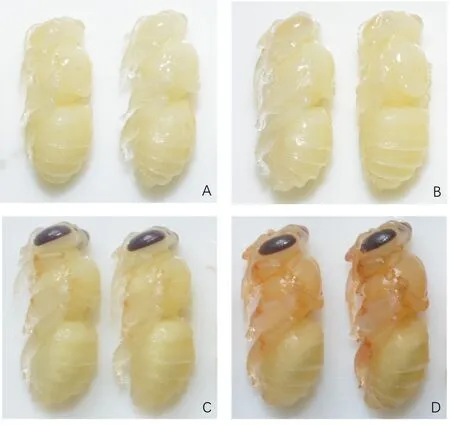
A:38 ℃胁迫5 d意蜂封盖子;B:35 ℃发育5 d的意蜂封盖子;C:38 ℃胁迫10 d意蜂封盖子;D:35 ℃发育10 d的意蜂封盖子.图2 分别在38和35 ℃条件下5和10 d的封盖子体色比较Fig.2 The morphs of capped brood at 38 ℃ vs 35 ℃ for 5 and 10 days
[1] COSSINS A R, BOWLER K. Temperature Biology of Animals[M].Dordrecht: Springer, 1987.
[2] BECK S D. Insect thermoperiodism[J]. Annual Review of Entomology, 2003,28(28):91-108.
[3] KLEINHENZ M, BUJOK B, FUCHS S H. Hot bees in empty broodnest cells: heating from within[J]. Journal of Experimental Biology, 2003,206(23):4 217-4 231.
[4] SEELEY T D. The wisdom of the hive: the social physiology of honey bee colonies[J]. Quarterly Review of Biology, 1995,89(6):907-908.
[5] COOK C N, KASPAR R E, FLAXMAN S M, et al. Rapidly changing environment modulates the thermoregulatory fanning response in honeybee groups[J]. Animal Behaviour, 2016,115:237-243.
[6] STABENTHEINER A, KOVAC H, BRODSCHNEIDER R. Honeybee colony thermoregulation-regulatory mechanisms and contribution of individuals in dependence on age, location and thermal stress[J]. PLoS ONE, 2010,5(1):e8967.
[7] HEINRICH B. The hot-blooded insects: strategies and mechanisms of thermoregulation[J]. Bioscience, 1993,44(11):184-193.
[8] KRONENBERG F, HELLER H C. Colonial thermoregulation in honey bees (Apismellifera)[J]. Journal of Comparative Physiology, 1982,148(1):65-76.
[9] GROH C, TAUTZ J, RÖSSLER W. Synaptic organization in the adult honey bee brain is influenced by brood-temperature control during pupal development[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004,101(12):4 268-4 273.
[10] TAUTZ J, MAIER S, GROH C, et al. Behavioral performance in adult honey bees is influenced by the temperature experienced during their pupal development[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003,100(12):7 343-7 347.
[11] LI Z, HUANG Z Y, SHARMA D B, et al. Drone and worker brood microclimates are regulated differentially in honey bees,Apismellifera[J]. PLoS ONE, 2016,11(2):e0148740.
[12] SEELEY T, HEINRICH B. Regulation of temperature in the nests of social insects[J]. Insect Thermoregulation, 1981:159-234.
[13] 朱翔杰,周冰峰,罗其花,等.温度对蜜蜂工蜂封盖期发育的影响[C]//2006年全国蜂产品市场信息交流会论文集.重庆:中国养蜂学会,2006:57-60.
[14] HIMMER A. Die temperaturverhältnisse bei den sozialen hymenopteren[J]. Biological Reviews, 1932,7(3):224-253.
[15] JONES J C, HELLIWELL P, BEEKMAN M, et al. The effects of rearing temperature on developmental stability and learning and memory in the honey bee,Apismellifera[J]. Journal of Comparative Physiology A, 2005,191(12):1 121-1 129.
[16] BECHER M A, SCHARPENBERG H, MORITZ R F A. Pupal developmental temperature and behavioral specialization of honeybee workers (Apismellifera, L.)[J]. Journal of Comparative Physiology A Neuroethology Sensory Neural & Behavioral Physiology, 2009,195(7):673-679.
[17] SINCLAIR B J. Linking energetics and overwintering in temperate insects[J]. Journal of Thermal Biology, 2015,54:5-11.
[18] WILLIAMS C M, HELLMANN J, SINCLAIR B J. Lepidopteran species differ in susceptibility to winter warming[J]. Climate Research, 2012,53(2):119-130.
[19] BOSCH J, KEMP W P. Effect of wintering duration and temperature on survival and emergence time in males of the orchard pollinatorOsmialignaria(Hymenoptera: Megachilidae)[J]. Environmental Entomology, 2003,32(4):711-716.
[20] 袁安,郭亚惠,黄晓,等.发育温度对蜜蜂归巢能力及记忆相关基因表达的影响[J].应用昆虫学报,2016,53(6):1 261-1 266.
[21] 陈琳,徐新建,王青,等.低温20 ℃对意大利蜜蜂未受精卵发育的影响[J].应用昆虫学报,2016,53(3):574-580.
[22] WANG Q, XU X, ZHU X, et al. Low-temperature stress during capped brood stage increases pupal mortality, misorientation and adult mortality in honey bees[J]. PLoS ONE, 2016,11(5):e0154547.
[23] 刘兆良,袁忠林,罗兰.日本龟蜡蚧雌成虫越冬前后耐寒性及相关生化物质含量的变化[J].昆虫学报,2017,60(1):120-126.
[24] 李娜,周晓榕,庞保平,等.轮纹异痂蝗卵的过冷却能力与其体内水分和生化物质含量的关系[J].昆虫学报,2014,57(7):745-753.
[25] VESTERLUND S R, LILLEY T M, OOIK T V, et al. The effect of overwintering temperature on the body energy reserves and phenoloxidase activity of bumblebeeBombuslucorum, queens[J]. Insectes Sociaux, 2014,61(3):265-272.
[26] 张青,涂永勤,刘怀,等.温度胁迫对小金蝠蛾幼虫抗氧化酶活性的影响[J].环境昆虫学报,2016,38(1):47-53.
[27] 杨丽红.柑橘全爪螨Panonychuscitri(McGregor)对热胁迫的响应机制研究[D].重庆:西南大学,2011.
猜你喜欢
孩子(2020年4期)2020-06-08
小学科学(学生版)(2019年11期)2019-12-09
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
科技与创新(2016年22期)2017-03-30
三联生活周刊(2017年9期)2017-03-03
科技视界(2016年20期)2016-09-29
特产研究(2016年3期)2016-04-12
作文评点报·中考版(2015年15期)2015-12-17
广西蚕业(2015年3期)2015-11-30
