
营巢树修枝对鹭类繁殖活动的影响
2018-07-05 08:22沈建新李如雪张淑霞
大理大学学报 2018年6期
沈建新,李如雪,徐 芳,张淑霞
(大理大学东喜玛拉雅研究院,云南大理 671003)
人类干扰的出现使野生动物对本来可以利用的食物食源、巢址、庇护地不再加以利用,因此人类对野生动物的干扰是生物多样性保护中的主要问题,而准确识别哪些人类活动对野生动物保护造成威胁也就尤为关键〔1〕。
对集群繁殖鹭类的人为干扰多数来自于科研调查和生态旅游活动,例如调查研究人员对巢内卵或幼雏的频繁接触会导致亲鸟弃巢或繁殖成功率下降,以及观鸟者闯入营巢地对亲鸟孵卵行为的干扰〔2〕。人类对种植林的强化管理(例如频繁修枝破坏树冠层)导致了种植林中鸟类多样性的下降〔3〕。人类对鹭类繁殖林的管理少有研究,营巢树的修枝给鹭类繁殖带来的影响尚不明确。
云南大理古城为世界著名旅游景点,虽为闹市区却为鹭类的集群繁殖地,该现象与其靠近洱海这一大型淡水湖泊湿地和周边发达的水稻农业密切相关。大理古城的鹭类繁殖地由夜鹭Nycticorax nycti⁃corax、白鹭Egretta garzetta、牛背鹭Bubulcus ibis混群形成〔4〕。当适宜的湿地营巢地资源有限时,鹭类选择靠近湿地、居民区的残留林斑营巢,可能是为了减少来自地面捕食者的干扰〔5〕。在2014年1月,大理古城原繁殖地的鹭类营巢树被人为修枝,营巢树树冠直径大幅缩减,繁殖鹭群被迫转移至距离该繁殖地400 m的新繁殖地进行繁殖活动。
在波斯湾沿海红树林中,树高和树冠直径为影响黄喉岩鹭Egretta gularis选择营巢树的主要因素,且树冠直径的影响更大〔6〕;繁殖鹭类的巢密度也取决于营巢树的结构和高度〔7〕。因此,营巢树结构的改变可能对鹭类繁殖活动产生较大的影响。大理古城鹭类原繁殖地的营巢树被修枝,将对鹭类繁殖产生怎样的影响。本研究对新繁殖地3种鹭类整个2016年繁殖季的活动进行了观察,试图通过比较原繁殖地和新繁殖地之间鹭类繁殖活动的时间和空间变异特点,来揭示繁殖鹭类对人为改变营巢树结构的适应机制。
1 研究方法
1.1 研究地概况 研究区域位于云南省大理市古城解放军第六十医院北围墙林带中,由原繁殖地和新繁殖地两片林区组成(图1)。原繁殖地的树木构成为整齐的圆柏同质林,详细介绍见文献〔4〕。新繁殖地为圆柏、桉树、合欢和大青树的混合林,树高不一致,为异质林。新繁殖地距离原繁殖地以东400 m处,北邻中和路,南至六十医院内,东邻叶榆路,西至六十医院北门口,东西长254 m,南北长16 m,面积共达2 814 m2。原繁殖地紧邻医院职工居民楼,由于大理地处风力较大区域,鹭类营巢的树木较高大,已经出现个别树木被大风吹倒的现象,出于保护居民楼安全考虑,医院管理方在2014年春季对原繁殖地鹭类营巢树进行了大幅修枝,多数营巢树的大侧枝被锯掉,仅留少量树冠和主干。人为对营巢树的修枝导致繁殖鹭类放弃了对原繁殖地的利用,2014年开始利用新繁殖地进行繁殖。
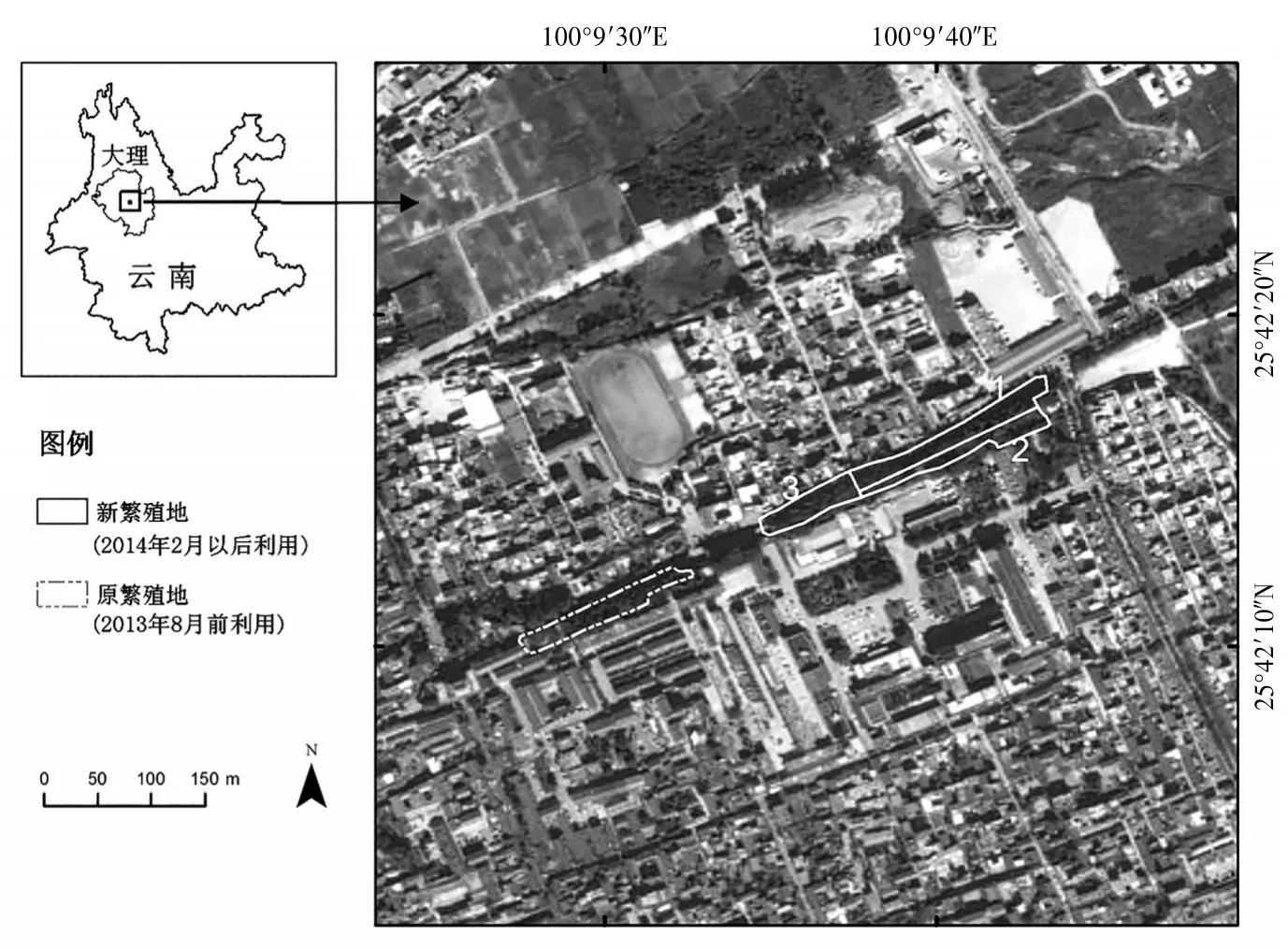
图1 大理古城的鹭类繁殖地
根据2016年进入该繁殖地鹭类的繁殖活动情况,将东西水平方向上分为3个片区:1区和2区为3种鹭类的混合营巢区,3区为开始繁殖较晚的夜鹭繁殖区。1区面积为1 206 m2,以中和路与叶榆路交点为坐标原点,东西向长度近140 m,南北方向为不规则边缘,最近距中和路4 m,最远距中和路15 m,平均宽度为9 m,是多数鹭类的繁殖区域;区内树种类型丰富,主要为圆柏、桉树,少量合欢和大青树。2区面积为536 m2,位于1区以南(六十医院内),东西长度约为75 m,南北宽度约为7 m;1区和2区之间有围墙相隔,2区为封闭区域,无法进入,区域内植株主要为圆柏及藤蔓类。3区面积为1 080 m2,位于1区以西方向,东西向长近100 m,南北向窄,平均宽约9 m,为夜鹭繁殖区,区域内营巢树全为桉树。
1.2 调查方法 在1区和2区内,2016年2月下旬开始每隔3~4 d于16:00~19:00到繁殖地观察有无鹭类在此栖息筑巢,判断鹭类迁入该繁殖地的时间,共监测3次。3~6月为筑巢时间,持续监测17次,每次监测均分种计数巢数;期间,通过多次观察营巢鸟种来确定巢主。6月,在1区内使用BOSCH(GLM 7000)红外激光测距仪陆续测量每个确定物种巢的垂直高度和巢距主干的距离。由于2区无法进入,因此未对其进行巢的完全计数和准确定位。3区为夜鹭繁殖区域,仅对巢数进行了完全计数,未进行巢空间特征调查。7~8月期间每隔7 d对1区和2区繁殖鹭类调查一次,记录其离巢情况,至8月9日所有鹭类离巢而去,共监测5次。对3区夜鹭群体的监测自6月初开始,直至8月20日所有夜鹭离去,共监测8次。自2016年2月至8月,整个鹭类繁殖期共监测25次。
为与原繁殖地的巢空间特征进行对比,采用与原繁殖地相同的方法分析了1区混群繁殖鹭类的巢空间特征〔4〕。调查以中和路与叶榆路东北角交点处为原点,记为(0,0);人为划分以东西方向为x轴、南北方向为y轴,向西最远记为(250,0),向南最远记为(0,16)。2016年2月初,在鹭类进入繁殖地之前,使用Bushnell-Elite1500红外激光测距仪以10 m为单位划分水平坐标,并进行标记。东西方向上以20 m为单位,将0~140 m划分为7个资源等级;南北方向上以3 m为单位,将其划分为4个资源等级;垂直高度以5 m为单位,将0~20 m划分为4个资源等级;巢距主干的距离以0.5 m为单位,将0~4 m划分为9个资源等级(含0 m)。
1.3 数据分析方法 3种鹭在东西水平方向、南北水平方向、巢高和巢距主干距离的生态位宽度计算方法见文献〔4〕。上述计算是基于1区巢的空间定位数据进行的。
使用Pearson's Chi-squared test分别检验3种鹭类在东西水平方向、南北水平方向、垂直方向上、巢距主干距离上不同资源等级间比例分布是否有显著差异,检验在统计软件R 2.15.0中完成。
2 结果
2.1 群落组成 与原繁殖地一样,2016年新繁殖地的繁殖鹭群依然由夜鹭、白鹭和牛背鹭组成。据不完全统计,巢数分别为163、105和126个,见表1。每种鹭类个体总数按每巢1对鸟计算,2016年在大理古城混合林繁殖地栖息的鹭类总数量为788只,密度0.280只∕m2,夜鹭为优势种。

表1 2016年大理古城鹭类繁殖地巢数量分区计数
1区完全计数3种鹭巢共258个,繁殖个体密度0.428只∕m2,占新繁殖地总巢数的65.5%。与原繁殖地不同的是,新繁殖地出现了单一夜鹭繁殖种群,出现在3区,据完全计数,共统计繁殖夜鹭巢42个,但繁殖个体密度较低,仅有0.078只∕m2。
2.2 繁殖活动时间变化 与原繁殖地不同,新繁殖地的鹭类繁殖活动时间持续更长。2016年大理古城新繁殖地的鹭类繁殖活动从2月末开始,结束于8月(图2)。1区和2区的混群鹭类繁殖活动规律为:2月末,有少量夜鹭首先迁入;紧接着3月初白鹭迁入,然后夜鹭和白鹭分批迁入;牛背鹭4月初才迁入;5月下旬至6月上旬,夜鹭幼鸟基本长成并大部分离去;7月下旬牛背鹭开始逐步离开;8月初,白鹭离开;而3区的单一夜鹭繁殖群的繁殖活动始于5月底,离开也最晚,直至8月20日才完全离开。
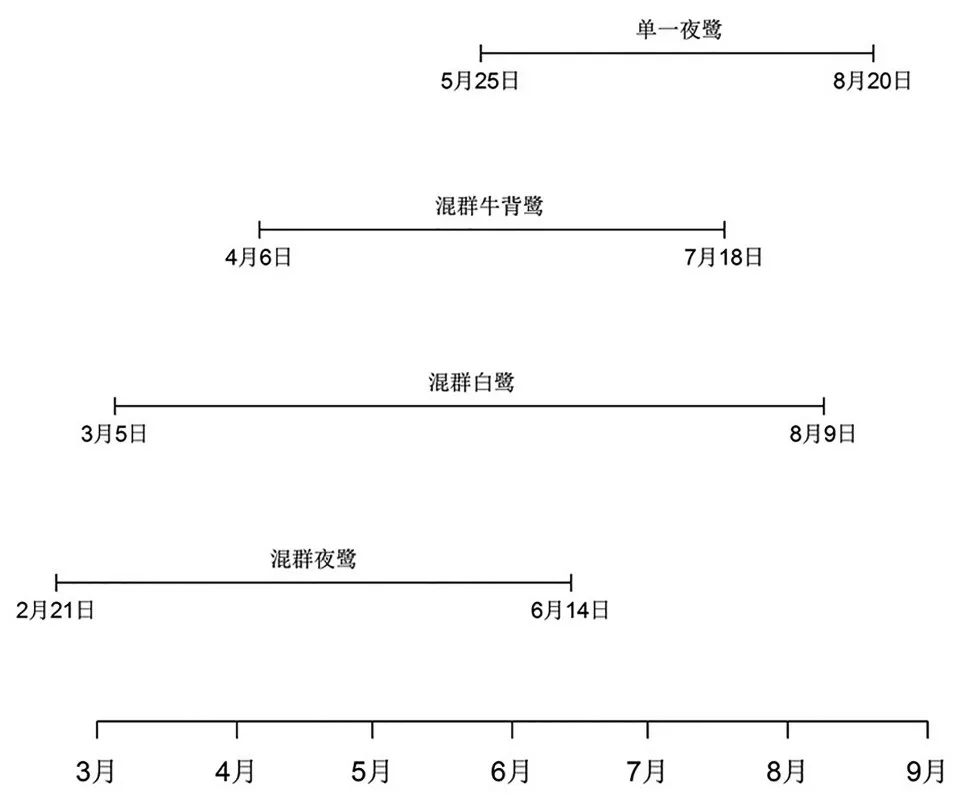
图2 3种鹭类在大理古城新繁殖地不同繁殖群体中的停留时间(2016年)
2.3 新繁殖地混群鹭巢的生态位宽度与原繁殖地的比较
2.3.1 东西水平方向上的分布 3种鹭类在东西水平方向上的分化显著(χ2=86.893,df=12,P<0.01)。在生态位宽度上,夜鹭最大,为0.944;牛背鹭最小,为0.592。在距离叶榆路最近的0~20 m上,夜鹭占有的巢数最多,有18个,而牛背鹭和白鹭的较少。与原繁殖地3种鹭类的生态位值相比,在鹭巢东西水平方向分布上,夜鹭的生态位宽度值变大,而牛背鹭的变小。见表2。2.3.2 南北水平方向上的分布 3种鹭类在南北水平方向上的分化显著(χ2=45.599,df=6,P<0.01)。在生态位宽度上,夜鹭最大,为0.971;牛背鹭最小,为0.745。在距离中和路最远的13.1~16.0 m上,夜鹭占有的巢数最多,有35个,白鹭的较少,牛背鹭则没有。与原繁殖地相比,在鹭巢东西水平方向分布上,夜鹭的生态位宽度值变大,而牛背鹭的变小。见表3。
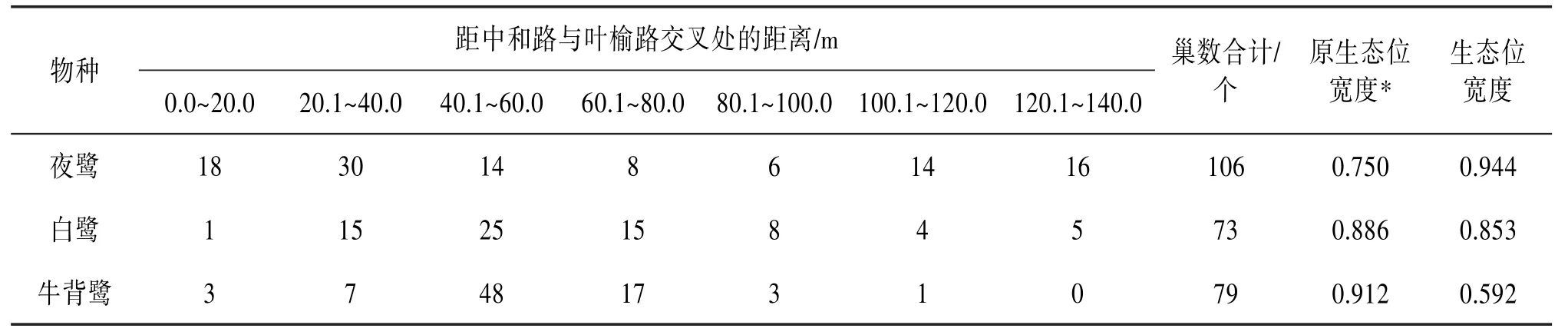
表2 大理古城鹭类新繁殖地鹭巢东西水平方向上的巢数分布

表3 大理古城鹭类新繁殖地鹭巢南北水平方向上的巢数分布
2.3.3 垂直方向上的分布 3种鹭类在垂直方向上的分化显著(χ2=30.186,df=6,P<0.01)。在生态位宽度上,夜鹭最大,为0.669;牛背鹭最小,为0.404。夜鹭对林冠层(15.1~20.0 m)占领最多,有26个巢;而牛背鹭对林冠层的巢位占领最少,仅有2个。与原繁殖地相比,在鹭巢垂直分布上,夜鹭的生态位宽度在新繁殖地变大,牛背鹭的生态位宽度则变小。见表4。
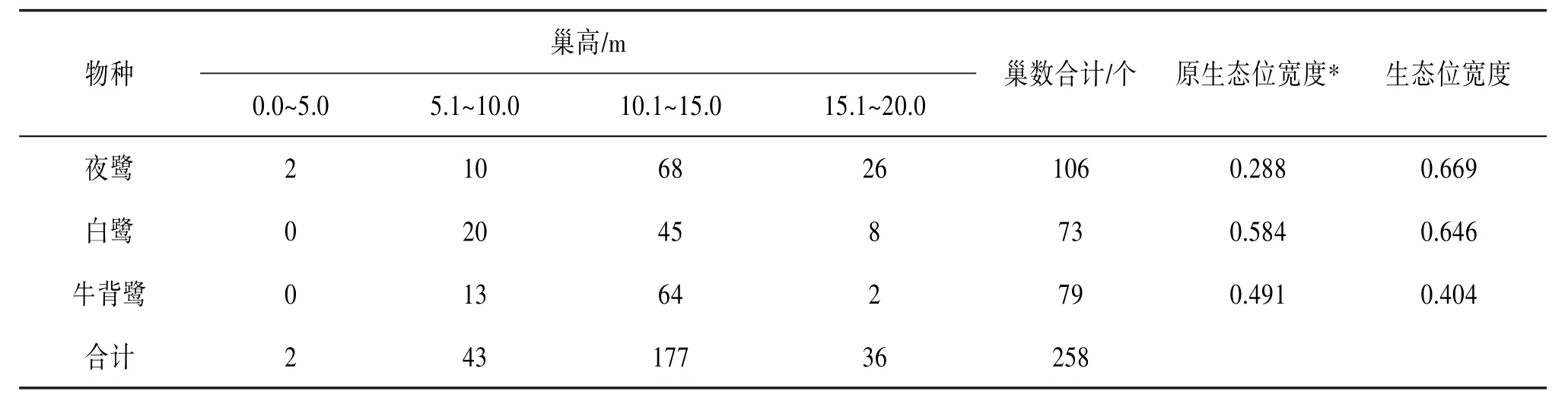
表4 大理古城鹭类新繁殖地鹭巢垂直方向上的巢数分布
2.3.4 巢距主干距离上的分布 3种鹭类在巢距主干距离上的分化显著(χ2=26.752,df=6,P<0.01)。在生态位宽度上,夜鹭最大,为0.878;白鹭最小,为0.580。夜鹭和白鹭位于巢主干上的巢数较多,分别有35和33个;而牛背鹭位于巢主干上的巢较少,有22个。与原繁殖地相比,在巢距主干的距离上,夜鹭在新繁殖地的生态位变大,牛背鹭变小。见表5。
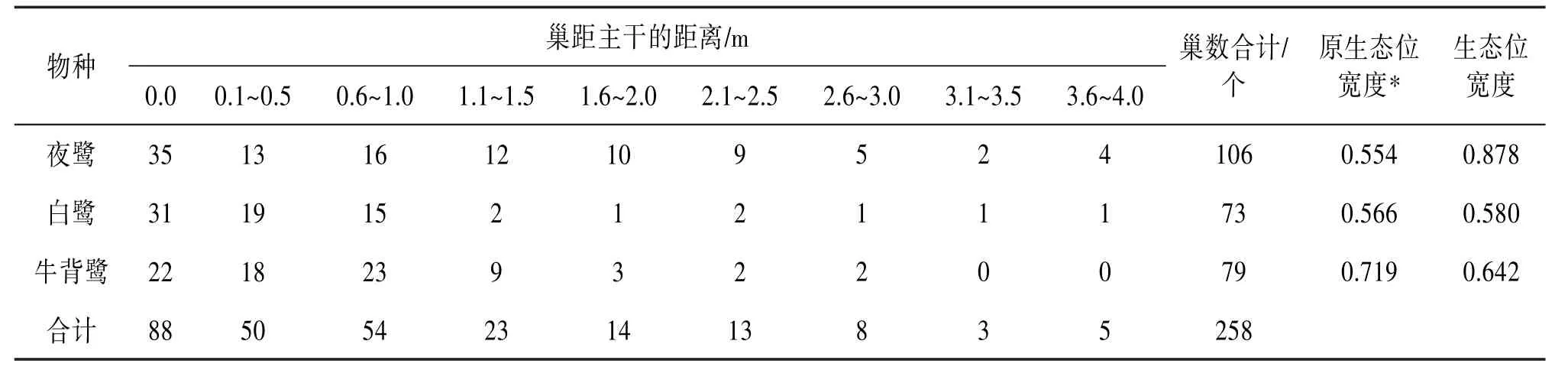
表5 大理古城鹭类新繁殖地鹭巢距树主干距离上的巢数分布
3 讨论
3.1 繁殖鹭群组成的新变化 与原繁殖地相比,新繁殖地鹭类组成上多出了活动于3区的单一夜鹭繁殖群。这群繁殖夜鹭的开始时间较晚,在5月底才开始筑巢活动,离开也晚于混群繁殖的鹭类;在空间上也与混群繁殖的鹭类重叠,单独占据3区进行繁殖活动。目前这一单一夜鹭群的形成有两种可能,一种可能为混群繁殖夜鹭失败者开始的二次繁殖,因为1区混群繁殖鹭类中,夜鹭到来时间是2月底,为混群繁殖的3种鹭类中最早开始繁殖的物种,繁殖失败时尚有时间开始二次繁殖。另一种可能是来自本地留鸟的繁殖群体,不同于混群繁殖的夜鹭群体,后者可能为夏候鸟繁殖群体,因为夏候鸟的繁殖时间通常早于本地留鸟〔8-9〕。这两种可能有待于未来对单一夜鹭群繁殖个体进行卫星跟踪的调查。
3.2 新繁殖地繁殖活动持续时间发生的变化 在混群鹭类繁殖活动持续时间上,新繁殖地的比原繁殖地的更长。在新繁殖地,混群繁殖的鹭类繁殖活动从2月底持续至8月,而原繁殖地混群鹭类的繁殖时间为2月底至5月底〔3〕。激烈的竞争压力,例如对巢位的抢夺,或食物资源的不足均可能使得混群繁殖鹭类的繁殖效率降低。
3.3 新繁殖地混群繁殖鹭类的生态位宽度发生的变化 通过对比新旧繁殖地混群鹭类的巢空间分布数据变化,发现夜鹭在巢东西水平方向、南北水平方向、垂直方向和巢距主干的距离等4个方向的生态位宽度上均明显变大,而牛背鹭的均明显变小,而白鹭的仅在东西水平方向上变小,其他3个方向上均变大,且变化幅度不如牛背鹭和夜鹭的大。作为最早开始繁殖的混群夜鹭,其到达新繁殖地后首先占据了大量的有利巢位,其生态位宽度值在4个方向上均变大,且均为3种鹭类中的最大值。紧接着到来的白鹭,占据了夜鹭部分未利用的巢位,因此生态位也有一定的扩展,其生态位宽度在3个方向上均变大。最后到达的牛背鹭个体能利用的巢位已经比较紧张,因此其生态位宽度被前两个物种严重压缩,4个方向上的生态位宽度值均已变小。
已知高位巢可以显著提高鹭类的繁殖成功率〔10-11〕,因为高位巢可以提供更好的庇护以躲避天敌〔12-13〕。牛背鹭的巢址生态位宽度被压缩后,其高位巢的数量为3种鹭类中最少的,因此,人为修枝活动可能对最后进入繁殖地的牛背鹭繁殖造成了较大的负面影响。
3.4 对鹭类营巢树进行修枝须谨慎 通过本研究对大理古城新旧繁殖地的鹭类繁殖活动和生态位宽度值变化的比较,发现新繁殖地的繁殖鹭类面临更大的繁殖压力;与原繁殖地相比,表现为繁殖活动持续时间的延长,牛背鹭的生态位宽度明显降低。基于本研究结果,建议在城市绿化管理中,对鹭类营巢树的修枝须谨慎进行,不宜大幅修枝。
致谢:感谢中国三江并流区域生物多样性协同创新中心和云南省高校洱海流域保护与可持续发展研究重点实验室对本文提供的帮助。
〔1〕GILL J A.Approaches to measuring the effects of human disturbance on birds〔J〕.Ibis,2007,149(S1):9-14.
〔2〕CARNEY K M,SYDEMAN W J.A review of human distur⁃bance effects on nesting colonial waterbirds〔J〕.Waterbirds,1999,22(1):68-79.
〔3〕PHIPOTT S M,ARENDT W J,ARMBRECHT I,et al.Bio⁃diversity loss in Latin American coffee landscapes:review of the evidence on ants,birds,and trees〔J〕.Conservation Biology,2008,22(5):1093-1105.
〔4〕WERSCHKUL D F,MCMAHON E,LEITSCHUH M.Some effects of human activities on the Great Blue Heron in Ore⁃gon〔J〕.Wilson Bulletin,1976,88:660-662.
〔5〕赵俊松,李廷野,房以好,等.云南大理古城3种繁殖鹭类的巢空间隔离〔J〕.四川动物,2015,34(3):357-363.
〔6〕PARKES M L,MORA M A,FEAGIN R A.Using scale,cov⁃er type and GIS to evaluate nuisance cattle egret colony site selection〔J〕.Waterbirds,2012,35(1):56-63.
〔7〕ETEZADIFAR F,BARATI A.Nest-site selection of west⁃ern reef heron(Egretta gularis)in relation to mangrove(Avicennia marina)structure in the Persian Gulf:implica⁃tion for management〔J〕.Forest Ecology and Management,2013,310:74-79.
〔8〕AYAŞ Z.Nest site characteristics and nest densities of Ar⁃deids(Night Heron:Nycticorax nycticorax,Grey Heron:Ar⁃dea cinerea,and Little Egret:Egretta garzetta)in the Nali⁃han Bird Sanctuary (Sariyar Reservoir,Ankara,Tur⁃key)〔J〕.Turkish Journal of Zoology,2008,32:167-174.
〔9〕BIBBY C,BURGESS N D,HILL D A,et al.Bird census techniques〔M〕.London:Academic Press,1992.
〔10〕BACHIR A S,BARBRAUD C,DOUMANDJI S,et al.Nest site selection and breeding success in an expanding species,the cattle egretBubulcus ibis〔J〕.Ardea,2008,96(1):99-107.
〔11〕BARATI A,ASHOORI A.Breeding success of blackcrown night heron(Nycticorax nycticorax),little egret(Egretta garzetta)and cattle egret(Bubulcus ibis)(Aves:Ardeidae)in relation to nest height in the South Caspian Sea〔J〕.Italian Journal of Zoology,2013,80(1):149-154.
〔12〕FASOLA M,ALIERI R.Nest site characteristics in rela⁃tion to body size in herons in Italy〔J〕.Colonial Water⁃birds,1992,15:185-192.
〔13〕ETTERSON M,NAGY L R,ROBINSON T R.Partitioning risk among different causes of nest failure〔J〕.Auk,2007,124:432-443.
猜你喜欢
文萃报·周二版(2022年4期)2022-02-14
甘肃林业科技(2021年3期)2021-03-30
青年歌声(2021年2期)2021-03-05
音乐世界(2020年4期)2020-09-10
东坡赤壁诗词(2019年3期)2019-07-05
民族音乐(2018年5期)2018-11-17
山西林业科技(2018年4期)2018-03-19
野生动物学报(2017年1期)2017-09-18
Coco薇(2017年8期)2017-08-03
中南林业科技大学学报(2015年9期)2015-12-21
