
脾气虚大鼠海马神经元线粒体呼吸链功能及Lon蛋白酶表达变化及意义
2018-07-27 03:35马丹刘文俊于化新王凌志刘旭东刘慧慧王路柴纪严单德红
山东医药 2018年26期
马丹,刘文俊,于化新,王凌志,刘旭东,刘慧慧,王路,柴纪严,单德红
(辽宁中医药大学,沈阳110847)
中医认为,脾主运化,在志为思,而海马能够调控消化吸收,又是情绪、思维和学习记忆等形成的重要部位,因此脾与海马关系密切,实验也证实二者确有关联[1,2]。线粒体是细胞动力工厂,线粒体DNA(mtDNA)与核DNA共同编码呼吸链复合体(C)Ⅰ、Ⅲ~Ⅴ。线粒体通过维持其蛋白质的正确折叠组装和错误折叠蛋白降解的平衡来达到线粒体稳态,即线粒体蛋白质控。与核DNA相比,mtDNA极易突变而致上述蛋白错误折叠,发生未折叠蛋白反应,如不及时干预则抑制ATP合成,甚至促进反应性氧簇(ROS)产生,引起细胞凋亡[3~5]。线粒体内存在Lon蛋白酶,能够分解这类异常蛋白,发挥蛋白质控作用[6,7]。研究发现,脾气虚大鼠海马线粒体损伤、产能减少[2,8],但是否发生了未折叠蛋白反应还不清楚。2015年1月~2017年12月,我们观察了脾气虚大鼠海马神经元线粒体内蛋白积聚情况,并检测线粒体呼吸链功能变化及Lon蛋白酶表达情况,从线粒体蛋白质控方面探讨脾气虚的发生机制。
1 材料与方法
1.1 动物、试剂与仪器 SPF级雄性SD大鼠16只,体质量(200±10)g,购于辽宁省本溪巿实验动物中心。ATP测定试剂盒(南京建成生物工程研究所),大鼠ROS检测ELISA试剂盒(Tsz Biosciences),线粒体呼吸链复合物活性检测试剂盒(北京索莱宝生物技术有限公司),抗Lon蛋白酶1多克隆抗体(Proteintech)。日本电子JEM-1011透射电镜,iMark酶标仪(美国Bio-Rad),电泳仪(美国Bio-Rad)。
1.2 动物分组与模型制备 大鼠置于室温18~23 ℃,相对湿度45%~55%,自由饮水进食,适应性饲养1周后开始实验。将大鼠按照随机数字表法分为正常组和模型组各8只。正常组正常饮食水。模型组参照文献[9]方法复制脾气虚证模型,即15 d内给予饮食失节(饱食1 d,禁食2 d,不禁水)和每日劳倦刺激(水温35~37 ℃游泳至力竭),当大鼠出现消瘦、食少、神疲和乏力时为模型制作成功。
1.3 海马神经元线粒体蛋白积聚情况观察 大鼠腹腔注射10%水合氯醛麻醉,快速开颅取海马。将大部分置于-80 ℃冻存备用,少部分在4 ℃下切成约1 mm3小块,2.5%戊二醛固定,PBS漂洗,1%锇酸固定,乙醇、丙酮梯度脱水,环氧树脂包埋聚合,LKB-1型超薄切片机切片,3%醋酸铀-枸橼酸铅双染,透射电镜观察线粒体内高密度斑块情况并拍片。
1.4 海马线粒体呼吸链CⅠ~CⅣ活性、ATP和ROS水平检测 CⅠ~CⅣ活性、ATP检测采用比色法,ROS检测采用ELISA法。取冻存的海马组织,制备组织匀浆,应用Bio-Rad美国伯乐iMark酶标仪检测,严格按照说明书操作。
1.5 海马线粒体呼吸链CⅤ和Lon蛋白酶表达检测 采用Western blotting法。取冻存保存的海马组织20~40 mg,加入200 μL裂解液,冰上研磨后置于EP管中4 ℃离心10 min,吸取上清液。BCA法测定蛋白浓度后,每管加入2×SDS-PAGE蛋白上样缓冲液100 μL,100 ℃变性,按50 μg总蛋白上样量计算加样体积,SDS-PAGE凝胶电泳并转膜和切膜,37 ℃水浴5%脱脂奶粉/TBST封闭2 h,加入1∶1 000和1∶2 000的抗Lon蛋白酶1多克隆抗体和CⅤ抗体稀释液4 mL,4 ℃摇床过夜。TBST洗膜,加入4 mL HRP标记的山羊抗兔二抗(1∶5 000),GAPDH盒内加入4 mL HRP标记山羊抗鼠二抗(1∶5 000),室温摇床孵育1.5 h。TBST洗膜,加入ECL发光液,曝光显影。应用AlphaView SA软件检测蛋白的相对表达量。
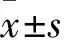
2 结果
2.1 两组海马神经元线粒体内异常蛋白积聚情况比较 正常组线粒体内部高密度斑块较少,而模型组线粒体内部高密度斑块则较多。
2.2 两组海马神经元线粒体呼吸链功能比较 见表1。模型组CⅣ活性、CⅤ表达和ATP水平均低于正常组(P<0.05或<0.01),其他指标比较差异无统计学意义(P均>0.05)。

表1 两组海马神经元线粒体呼吸链功能比较
注:与正常组比较,*P<0.05,**P<0.01。
2.3 两组海马神经元线粒体Lon蛋白酶表达比较 正常组与模型组Lon蛋白酶的相对表达量分别为0.249±0.080、0.169±0.057,模型组低于正常组(P<0.05)。
3 讨论
传统中医理论指出,脾能够将食物转化为精微物质,生成气血后为机体利用。现代医学认为,食物中的营养物质在胃肠道消化吸收后,要经过线粒体的氧化磷酸化才能产生ATP,进而供给细胞。因此认为,中医的脾与线粒体存在密切联系。脾气虚是临床常见证型,以食少、消瘦、腹胀和神疲乏力等为主要症状,关于其发生机制的研究虽然较多,但涉及海马神经元线粒体的报道较少[1,8]。海马是整合内脏功能与情绪、思维和学习记忆的高级中枢结构,能量供给不足将引起其功能紊乱[10,11]。线粒体产能主要依赖呼吸链CⅠ、Ⅲ~Ⅴ的协同作用,而这些蛋白异常可导致ROS等过度产生,使线粒体由动力工厂变成毒物工厂。研究表明,mtDNA存在上述复合物的7、1、3和2个编码基因[12,13],但由于缺乏组蛋白保护且靠近氧化磷酸化场所,mtDNA极易突变,从而导致包括上述成分在内的许多蛋白发生错误折叠,如不及时清除而大量积聚,就会损伤线粒体功能,甚至诱发氧化应激等严重反应[14~17]。
研究表明,Lon蛋白酶能够清除线粒体内异常蛋白,修复mtDNA,维持线粒体稳态[6,7],但其在脾气虚中的作用却少有研究。本研究首先进行形态学观察,以验证脾气虚时线粒体内是否存在异常蛋白积聚现象。电镜结果显示,正常组与模型组内均存在此种现象,但模型组要更严重。正常组海马神经元线粒体内出现少量蛋白积聚,表明即使在正常情况下,海马神经元的mtDNA也有突变现象,只是比较轻微,其原因可能是部分线粒体衰老或mtDNA受到了某些外来刺激;模型组出现大量蛋白积聚,提示脾气虚时海马神经元mtDNA突变严重,导致异常蛋白过多。其次在呼吸链方面,本研究显示模型组CⅣ活性和CⅤ表达显著下降,同时ATP含量明显减少,但ROS水平升高不明显,表明脾气虚时海马神经元线粒体呼吸链只是部分受损。由于CⅣ和CⅤ是ATP合成的关键环节,其表达下降对于ATP生成影响较大,这也可能是模型组ATP含量明显低于正常组的重要原因。
基于以上形态学与功能两方面的结果,可以推测脾气虚时海马神经元mtDNA发生了突变,导致了相关蛋白错误折叠,影响了呼吸链功能,但还未出现氧化应激反应,因此及时清除异常蛋白就显得极为必要。在线粒体的蛋白质控体系中,Lon蛋白酶较受关注。研究发现,Lon蛋白酶属于AAA+超家族,是一种同质寡聚环状的蛋白酶,在古生菌、原核生物和真核生物线粒体中高度保守,其突变可引起异常蛋白质降解缺陷[6,18]。Lon蛋白酶含量N端结构域、ATP酶结构域和P结构域,主要通过前二者识别并结合底物,后者完成蛋白降解。脾气虚时Lon蛋白酶表达的研究目前还未见报道。
本研究发现,正常组Lon蛋白酶表达较高,而模型组则明显降低,表明正常情况下,海马神经元线粒体内存在较多的Lon蛋白酶,可以应对异常蛋白错误折叠并及时清除,以保证线粒体产生充足ATP,而脾气虚时Lon蛋白酶不足,导致异常蛋白降解延迟,从而影响了呼吸链的产能。因此本研究认为,海马神经元内线粒体蛋白质控水平下降与脾气虚的发生有关。
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
趣味(作文与阅读)(2021年12期)2021-04-19
创新作文(小学版)(2019年10期)2019-09-25
文苑(2018年22期)2018-11-19
小学生学习指导(低年级)(2017年5期)2017-05-04
发明与创新·中学生(2016年4期)2016-05-14
中国粮油学报(2016年5期)2016-01-23
西南军医(2016年6期)2016-01-23
西南军医(2014年5期)2014-04-25
红蜻蜓·低年级(2014年1期)2014-02-19
