
RNAi沉默藏猪TGF-β1对PRRSV病毒复制和免疫的效应研究
2018-08-01 02:19王晔
畜牧兽医科技信息 2018年7期
王 晔
(四川大学生命科学学院生物资源与生态环境教育部重点实验室,四川 成都 610064;四川省动物疫病预防与食品安全重点实验室,四川 成都 610064)
猪生殖和呼吸综合症病毒(PRRSV)是动脉病毒科、动脉病毒属的高致病性单正链RNA病毒,突变率高。猪群感染该病毒后,主要表现为呼吸系统疾病、流产、以及继发性病毒和/或细菌感染,而且因为PRRSV会抑制宿主的免疫系统,故会使宿主处于长期患病状态,给全球的养猪产业带来巨大的经济损失。
PRRSV表达的蛋白是多功能型的,能有效弥补RNA病毒有限的编码能力。结构蛋白和非结构蛋白几乎都参与调控宿主免疫系统,躲避宿主的攻击。目前发现的调控机制已有很多,包括:改变免疫相关细胞数量和功能;促进IL-10和TGF-β1表达使其发挥免疫抑制功能,进而破坏早期炎症因子与抗炎症因子的平衡;抑制中和抗体的生成,同时促进非中和抗体合成。
在免疫系统中,TGF-β有维持免疫系统稳定的作用,其主要作用方式是通过依赖于以及不依赖于Smad信号通路调节多种白细胞系的增殖、分化和活性,抑制免疫炎症因子的功能。已有研究发现,I型和II型PRRSV均可诱导TGF-β过表达,使宿主免疫系统失衡、瘫痪,病毒大量繁殖。TGF-β有三个亚型——TGF-β1~3,其中在免疫系统发挥主要作用的是 TGF-β1,同IL-10一起发挥免疫抑制功能。而TGF-β2和TGF-β3则主要在间叶组织和骨骼中大量表达。Gómez-Laguna等已证实 TGF-β1可以通过抑制IFN-γ合成和促进IL-10的表达来抑制巨噬细胞的生物活性。
藏猪是一种中国原始高原猪种,具有抗逆性强、适应高寒气候的特点,与其它猪种在表型和生理特征方面差异明显。但是因地域原因,目前国内外对藏猪免疫遗传及其免疫应答特性的研究较少。PRRSV在四川、西藏地区流传十分广泛,相关免疫预防成果不明显。故本次实验欲通过RNAi体外转染已被PRRSV感染的藏猪的外周血单核粒细胞,沉默细胞中TGF-β1基因的表达,通过RT-PCR的方法检测细胞中PRRSV病毒的复制情况以及抗病毒免疫应答激活情况。
1 材料与方法
1.1 准备藏猪外周血单个核细胞(Tp-PBMCs)
以EDTA-K2抗凝采血管(康健,江苏)收集藏猪上腔静脉血。将血液置于猪淋巴细胞分离液(灏阳,天津)上层,离心(550g,20min)分离得到 Tp-PBMCs。用 Hank’s平衡液(Hyclone,北京)清洗细胞,再用培养基重悬细胞,于37℃,5%CO2的培养箱中培养。
1.2 构建靶向TGF-β1的 shRNA表达载体
通过GeneBank获得猪 TGF-β1 mRNA序列(NM_214015),利用在线 SiRNA 设计软件(Http://jura.wi.mit.edu/bioc/siRNA;Http://www.ambion.com/techlib/misc/siRNA_finder.HtmL)设计4条靶向猪TGF-β1 mRNA的干扰序列(表1)。这些序列由Invitrogen公司(上海)合成,并构建到shRNA(短发夹RNA)表达载体pSilencer 3.1-H1neo中,得到4个重组质粒(shTGFβ1-1~4)。将它们分别转化到大肠杆菌DH5α中,在LB培养基中扩增,并经酶切和测序(华大基因,北京)验证质粒正确性。空载体在猪mRNA数据库(Taxid:9823,NCBI)中没有靶点,作为空白对照。所有用于转染的质粒均通过试剂盒纯化,并经鳌试剂内毒素检测试剂盒(A&C Biological Ltd,湛江)检测,内毒素含量少于0.125 EU/mL。
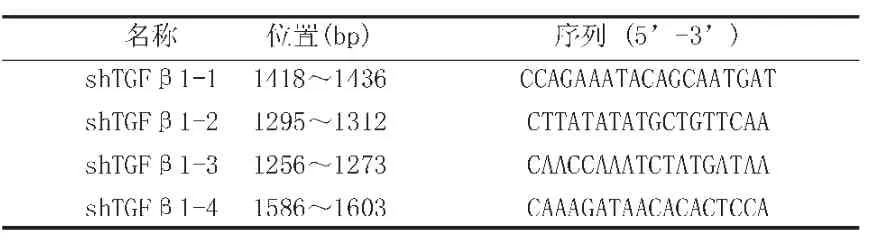
表1 干扰TGFβ-1的寡核苷酸序列 shTGFβ1-1~4
1.3 重组干扰质粒转染Tp-PBMCs
阳离子脂质体DMRIE-C(Invitrogen,USA)作为转染试剂。按照说明书准备转染复合物。混合物混匀后室温静置5min,将稀释后的质粒分别加入稀释的DMRIE-C中,室温静置30min。将Tp-PBMCs 550g离心15min,细胞经PBS清洗后以RPMI 1640(无血清无抗生素)重悬,密度为1.5×107个/mL。将0.2mL细胞悬液(3×106个细胞)添加到6孔细胞培养板中,然后分别加入转染复合物并混匀。在培养箱培养4.5h后,加入2mL RPMI 1640培养基和30μL PRRSV,继续培养。在设定的时间点将Tp-PBMCs以10000g离心30s,上清液冻存于-80℃。除质粒和病毒外,空白对照组经过上述相同的处理。
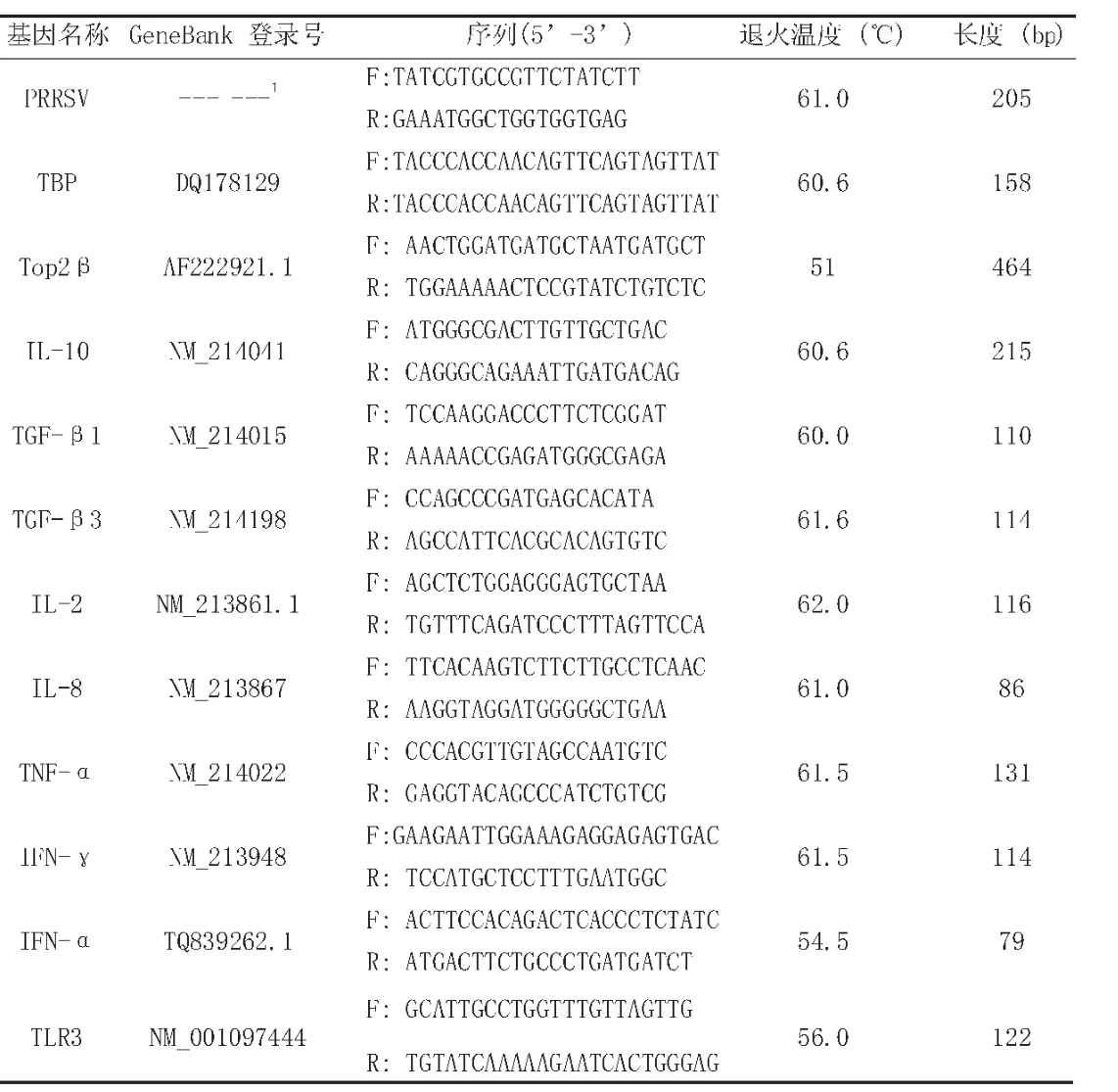
表2 实时荧光定量RT-PCR引物
Tp-PBMCs细胞活性由CCK-8试剂盒(7-sea,上海)测定。将3×105经处理过的细胞(PRRSV感染,转染等)加入96孔板,随后加入CCK8,混匀后培养4h,在酶标仪上读取480nm的吸光值,重复3次。每个处理设置五个重复孔。每24h检测1次。
1.4 实时荧光定量PCR(real-time qPCR)分析mRNA表达
将冻存的细胞从-80℃取出后,按照RNAiso plus说明书提取总RNA,用反转录试剂盒(Transgen,北京)合成cDNA。使用iQ5(BIO-RAD)进行real-time qPCR,分析每个基因的表达水平。反应程序如下:94℃预变形3min,40个扩增循环(94℃ 6s;退火 6s,72℃ 10s),随后进入溶解曲线循环(从55℃开始,每6秒增加0.5℃,直至95℃)。重复三次检测。TBP(TATA box binding protein)和DNA拓扑异构酶2-beta(Top2-β)作为内参基因。基因的相对表达量由2-ΔΔCT计算得出。所有引物的信息列于(表2)中。
1.5 数据分析
所有试验均重复3次,对照组的值都设为1。数据以平均值±标准差的形式表示。
2 结果
2.1 shTGFβ1-1能抑制感染 PRRSV的 Tp-PBMCs转录T G F-β1
Tp-PBMCs经处理后培养36h(转染结束时为0h),收集细胞,然后通过real-time qPCR的方法检测TGFβ-1的mRNA水平,筛选4条质粒抑制TGF-β1的情况,发现shTFGβ1-1对TGF-β1的沉默效果最佳,相较PRRSV组,其mRNA转录水平下降了74.47%(表3),且抑制时效长达72h(表4),因此被选中用于后续。另外,除空白对照组外,还设有PRRSV组为阳性对照组,DMRIE-C组和pNeg组均为阴性对照组,前者仅加入转染试剂,后者则向感染PRRSV的细胞转染空质粒载体。
另外,为排除假阳性情况,分别用空载体和转染试剂DMRIE-C转染Tp-PBMCs,培养36h后检测免疫相关因子的mRNA水平,结果表明除IFN-α略有升高外,不影响其余免疫相关因子的转录(表5)。
2.2 shTGFβ1-1抑制PRRSV的复制并增强感染PRRSV的Tp-PBMCs的细胞活性
PRRSV的mRNA水平在感染Tp-PBMCs后的72hpi达到峰值,在转染shTGFβ1-1后其mRNA水平明显受到抑制,抑制率高达99.0%(表4)。
另外,从(表6)可以看出,PRRSV的感染会降低Tp-PBMCs的细胞活性,而抑制TGF-β1的表达可以一定程度上恢复细胞活性,使活性升高约150%~160%。

表3 重组干扰质粒shRNA及对照组对TGF-β1的沉默效果
表4 重组干扰质粒shTGFβ1-1对PRRSV复制和TGF-β1、TGF-β3的mRNA水平的影响
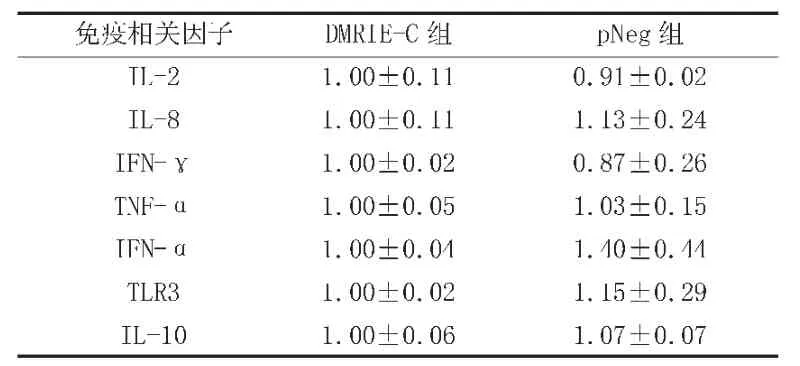
表5 空载体和转染试剂对Tp-PBMCs免疫相关细胞因子mRNA水平的影响
表6 重组干扰质粒shTGFβ1-1对细胞活性的影响
2.3 shTGFβ1-1对PRRSV感染的Tp-PBMCs中免疫相关基因表达的影响
由于免疫相关基因的表达情况可以反应细胞的免疫应答的状态,因此本实验用RT-PCR分析了一些代表性的免疫相关基因的mRNA水平以推断shTGFβ1-1对Tp-PBMCs免疫应答的调控情况。
如表4所示,PRRSV感染后会持续抑制Tp-PBMCs中TGF-β3的表达,但当转染干扰质粒后的前48h其mRNA水平就会有所升高,这可能是其在体外对TGF-β1缺失的一种补偿效应。
如表7所示,PRRSV感染Tp-PBMCs后会下调TLR-3的表达,但转染shTGFβ1-1后72h,其mRNA水平有快速、明显升高。IFN-α和TNF-α的mRNA水平在PRRSV感染后的前24h处于抑制状态,随即又恢复到正常水平,转染shTGFβ1-1 72h后,其mRNA转录就被上调。伴随TGF-β1基因表达的抑制,被PRRSV波动性调控的IFN-γ的mRNA水平快速升高。与Ctrl组相比,shTGFβ1-1的转录会明显上调IL-2的转录,但PRRSV感染Tp-PBMCs后对其表达影响较小。PRRSV感染后会快速诱导IL-8转录,相反shTGFβ1-1会降低IL-8的mRNA水平。另外,与TGFβ-1同为免疫抑制因子的IL-10在细胞被PRRSV感染后,转录水平明显升高并在感染后72h达到峰值,但是当细胞中TGF-β1表达被沉默后,其mRNA水平相比Ctrl组而言并没有明显变化,表达没有受到影响。
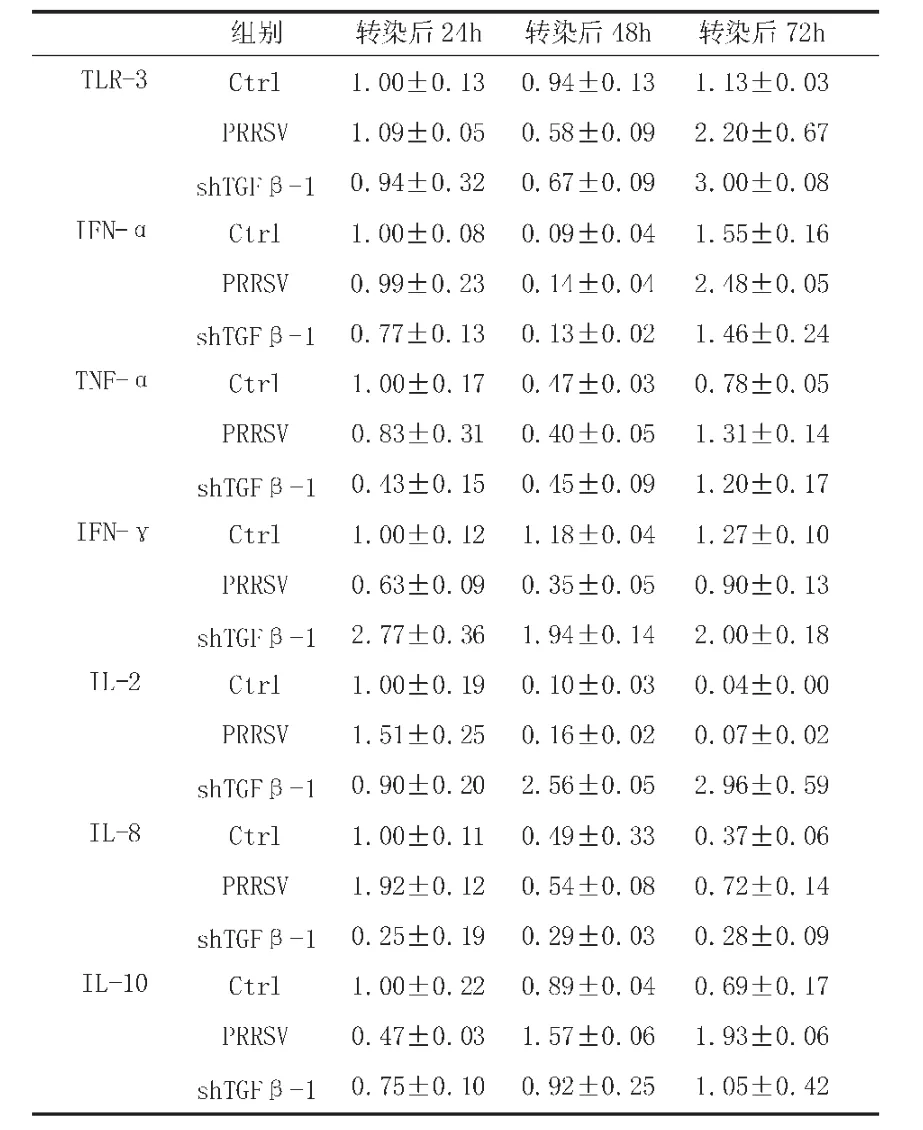
表7 重组干扰质粒shTGFβ1-1对免疫相关细胞因子mRNA水平的影响
3 讨论
现在已有报道证实PRRSV的复制能够诱导TGF-β的表达,PRRSV会通过激活Tregs(即Th3亚群)来抑制宿主免疫应答,TGF-β是后者的主要效应分子。过量表达的TGF-β1是PRRSV抑制宿主免疫系统的基础,考虑到它在PRRSV感染宿主细胞方面的重要地位,本篇欲利用RNAi探究是否可以通过沉默TGF-β1表达达到抑制PRRSV复制和激活被PRRSV抑制的免疫应答反应。结果表明,干扰TGF-β1可以成功抑制PRRSV在藏猪外周血单个核细胞中的复制,且抑制率高达99.0%,同时使细胞活性升高约150%~160%,另外成功激活多个抗病毒免疫相关基因转录。这表明TGF-β1在PRRSV复制过程中发挥重要作用,更重要的是,这有可能成为抵抗PRRSV感染的新方法。
TGF-β1是多功能性细胞因子,破坏其表达可能会严重影响细胞功能。已有研究发现,在体外TGF-β3对TGF-β1的缺失有一定的补偿作用。本次试验也进一步验证了这个说法,转染shTGFβ1-1后,细胞中TGF-β3的mRNA水平明显升高。但是升高的TGF-β3是否弥补了免疫抑制功能还需进一步试验确定。
IL-8在介导白细胞迁移至感染部位过程中起重要作用。本篇研究结果表明,PRRSV感染会快速诱导IL-8表达升高,这与Reeth等人的研究结果一致,参与病毒感染更多白细胞。沉默TFG-β1所引起的IL-8 mRNA水平降低可能与PRRSV复制被抑制相关。
研究表明,PRRSV会破坏TLR3的抗病毒能力,以此降低宿主的免疫应答能力,本篇研究结果与之一致。感染PRRSV病毒后,宿主细胞中TLR3的mRNA水平先降后生,抑制TFG-β1后72h,其mRNA水平快速、明显升高,表明此时细胞开始启动固有免疫应答。
IFN-γ水平宿主与抵抗和清除PRRSV有关。向已感染病毒的细胞转录shTFGβ1-1后IFN-γ和TNF-α的mRNA水平都有所升高,这可能与效应TH1细胞的抗PRRSV免疫应答反应被激活有关。另外,干扰细胞中TFGβ-1的表达后,IL-2的mRNA水平明显升高,这说明TH2免疫应答也被激活了。
沉默TFG-β1后,Tp-PBMCs中IL-10的mRNA水平也明显降低,这可能与细胞中IFN-γ,IFN-α和TNF-α的大量表达有关。
4 结论
本次实验首次筛选出可以高效、稳定地抑制猪外周血单个核细胞中TGF-β1基因表达的重组质粒。体外实验表明,已感染PRRSV的Tp-PBMCS在转染该干扰质粒后细胞活性明显增强,抗病毒免疫应答被激活,病毒复制明显受到抑制。为增强藏猪抵抗PRRSV感染提供了新的研究思路。
中国农业科学院重大产出科研选题“重大动物疫病新型标记疫苗创制”启动会在哈尔滨召开
2018年6月27日,中国农业科学院重大产出科研选题“重大动物疫病新型标记疫苗创制”启动会在哈尔滨兽医研究所顺利召开。
应邀出席会议的有中国农业科学院科技局熊明民副局长、项目处干部解沛;中国农业科学院财务局张士安副局长、姜成钢副处长。项目参与单位中国农业科学院哈尔滨兽医研究所、兰州兽医研究所、上海兽医研究所的相关团队首席科学家及主要研究人员近20人参加了本次会议。
项目主持人哈尔滨兽医研究所所长步志高研究员从重大动物疫病新型标记疫苗创制的重要意义入手,对重大选题研究内容、研究目标、研究进展、实施方案、实施机制、经费执行要求等内容进行了详细的汇报。熊明民副局长强调了院重大产出科研选题的立项背景和重要性,并对项目的组织实施提出具体的指导意见,强调了项目执行过程中要坚持产出导向,希望项目成员认真完成考核指标,产生重大标志性成果。
启动会上,与会专家针对各课题取得的重要进展、存在的问题以及项目任务落实和组织管理等方面进行了充分的讨论。
猜你喜欢
心理学报(2022年8期)2022-08-09
数学物理学报(2021年6期)2021-12-21
江西农业学报(2021年4期)2021-04-20
水生生物学报(2021年1期)2021-02-04
科学(2020年3期)2020-11-26
基层中医药(2020年3期)2020-09-11
当代水产(2020年3期)2020-06-15
食品科学(2020年4期)2020-03-11
疯狂英语·新读写(2018年3期)2018-11-29
小星星·阅读100分(高年级)(2015年11期)2015-11-28
