
云雾山典型草原群落空间异质性对不同干扰方式的响应
2018-08-10 02:55何晴波赵凌平谭世图王占彬
西北农林科技大学学报(自然科学版) 2018年9期
何晴波,赵凌平,2,谭世图,2,王占彬,2
(1 河南科技大学 动物科技学院,河南 洛阳 471023;2 河南省饲草饲料资源开发与畜禽健康养殖院士工作站,河南 洛阳 471023)
草地不仅是发展畜牧业的重要经济资源,还是牧区、半牧区人们赖以生存的环境基础。草地的可持续发展利用关系到生态文明建设问题,也关系到牧区、半牧区经济的发展问题。对草地生态环境的了解,有助于草地的可持续发展利用[1-2]。在草地生态系统中,施肥、放牧、火烧、刈割和封育等都是常见的干扰方式,同时也是影响草地演替方向和演替进程的关键因子。许多研究表明干扰方式不仅可以改变群落的外貌特征、物种组成、物种多样性和空间分布格局,还对优势种群的形态结构、生态系统功能和繁殖策略有着直接的影响[2]。研究干扰对草地生态系统的影响对草地资源的合理利用、退化草地的植被恢复和物种多样性的保护具有重要意义。
空间异质性是指生态学过程的不稳定性和空间分布上的格局存在的不均匀性及其复杂性[1]。空间异质性影响生态系统的功能和进程,是生态学研究的核心。空间格局一方面是空间自相关和异质性的具体表现,另一方面又是干扰在各种尺度域上作用的结果[2-3]。在群落中,种群格局的形成,一方面与物种自身的特征相关,另一方面则取决于群落环境。群落环境包括生物因子和非生物因子,而干扰则是群落中最常见的非生物因子之一。可见,干扰是引起空间异质性的重要因素之一。干扰可以改变草地的物种组成、空间结构,提高或降低草地的空间异质性,从而影响草地的生态功能和进程[4]。研究不同干扰方式对草地植物群落空间异质性和对物种空间分布格局的影响,有助于揭示空间分布格局与其相关联的生态学过程的相互关系[5]。
针对不同类型数据的空间异质性,有着不同的定量分析方法[4]。从分维分析到分布拟合法、小波分析、分型形析和地统计学,都是十分有效且被许多学者常用的空间异质性研究方法[6-7]。幂函数法最早是由Taylor[8]于1960年提出,后被Shiyomi等[9]、盐见正卫等[10]运用到草地种群的分析与研究中,该方法以后就被研究者广泛地应用到草原群落空间异质性领域。霍光伟等[11]采用幂函数法则探讨了轻度、中度和重度放牧对草原群落空间异质性的影响;黄琛等[12]基于幂函数法则研究了克氏针茅草原群落空间异质性对封育和放牧干扰的响应特征;霍光伟等[13]于2012年又研究了封育和放牧对克氏针茅草原植被空间异质性的影响。运用幂函数法则研究空间异质性,可以有效弥补传统调查分析方法耗时长、难度大的不足,同时避免了盖度调查法中测定群落特征所带来的人为误差[11],还可准确客观地体现不同物种的空间异质性特征[14]。
目前,关于草地生态系统物种空间异质性对同一干扰方式不同梯度的响应前人已开展了较多的研究。在科尔沁沙地,蒋德明等[15]探讨了不同封育年限对植被群落种群小尺度空间异质性的影响;在内蒙古荒漠草原,刘红梅等[16]采用GS+软件和地统计学分析方法对短花针茅植物的种群空间异质性分布进行了研究;在宁夏荒漠草原,魏乐等[5]研究了不同坡位的植物群落物种空间分布格局。但有关不同干扰因素对物种空间异质性影响的研究却鲜有报道,因此深入探讨天然草地群落空间异质性对不同干扰方式的响应,对于退化草地的恢复重建和物种多样性的保护具有重要意义。本研究以云雾山典型草原群落的主要植物种群为对象,结合幂函数法则、物种多样性和丰富度,研究了封育、放牧、火烧干扰对优势种群出现频率和空间异质性以及群落空间异质性特征的影响,以期为云雾山天然草地的管理利用与恢复重建提供依据。
1 研究区概况
研究区位于宁夏回族自治区固原市东北部45 km处的云雾山国家草原自然保护区(N 36°13′-36°19′,E 106°24′-106°28′),海拔为1 800~2 180 m。该区是黄土高原典型草原带的代表性区域,为温带大陆性半干旱季风气候,年平均温度5 ℃,≥10 ℃的年积温为2 370~2 882 ℃,气温最低月为1月(平均最低气温约-14 ℃),气温最高月为7月(22~25 ℃)。该区年平均降雨量400~450 mm,其中70%以上集中在7~9月;年平均蒸发量1 330~1 640 mm;年平均无霜期为112~140 d;植物以多年生草本植物为主,主要为大针茅(Stipagrandis)、本氏针茅(Stipabungeana)、甘青针茅(Stipaprzewalskyi)、干生苔草(Carexaridula)和百里香(Thymusmongolicus)等。
2 研究方法
2.1 野外调查
野外取样于2016年8月中旬进行。选取封育23年的草地、放牧地和火烧后5年的草地为试验地,在这3类试验地的典型地段分别设一条50 m的样线,然后在每个样地沿样线选择植被生长均匀、地形情况基本一致的地段依次设置50 cm×50 cm(简称“L”型样方)的样方100个,再将每个“L”型样方划分为25 cm×25 cm(简称“S”型样方)的4个小样方,因此共计400个“S”型样方。最后统计“S”型样方内的物种组成、频度、高度、多度、盖度和地上生物量等指标,并计算物种的重要值、出现频率、种群空间异质性指数和群落的空间异质性指数。
2.2 群落结构特征分析
2.2.1 种群的空间异质性 用幂函数法则分析种群的空间分布格局。幂函数法则是以lg [p(1-p)/n]作为x轴(其中p为物种在“S”型样方中的出现频率;n为一个“L”型样方中的“S”型样方数,n=4;p(1-p)/n为某物种随机分布时的方差),以lg [v/n2]作为y轴(其中v为实际观察中某物种出现的方差,v/n2为实际观察中物种出现频度的方差),做散点图,将y用x的单元回归方程来表示的一种经验法则[10]。
本研究中,定义δi为物种i的空间异质性指数,代表物种i的空间异质性程度(不均一性),δi=yi-xi,即δi为物种i的yi与随机分布直线(y=x)之间的距离差值[14]。
2.2.2 群落空间异质性 群落的空间异质性指数(δc)表示群落的不均一性或异质性程度,为群落中各个种群的空间异质性指数和该种群出现频度乘积的平均值,用下式计算:
δc=∑(pi×δi)∑pi。
式中:pi为物种i在“S”型样方中的出现频度。
2.2.3 物种的重要值 重要值用下式计算:
重要值=(相对高度+相对盖度+相对多度+相对频度+相对生物量)/5
式中:相对高度为某物种的高度(m)与样方中所有植物高度之和的百分比,相对盖度为某物种的盖度(%)与所有物种盖度之和的百分比,相对多度为某物种的丰富度与所有物种丰富度之和的百分比,相对频度为某物种频度与所有物种总频度的百分比,相对生物量为某物种的地上生物量(g/m2)与样方中所有物种地上生物量的百分比。
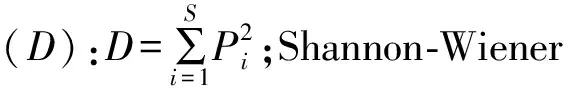
式中:N为样本中所有物种的个体总数,S为物种总数,Pi为物种i的个体数占所有物种个体总数的比例,既重要值。
2.3 数据统计与分析
用Excel 2003对数据进行预处理,运用SPSS 16.0软件进行单因素方差(ANOVA)分析。
3 结果与分析
3.1 不同干扰方式下物种出现频率的变化
本研究通过各个物种出现的频率来确定不同群落的优势种。由表1可知,在云雾山典型草原,放牧地植物群落由29个物种组成,其优势种为干生苔草、百里香、猪毛蒿和散穗早熟禾;封育地由28种植物组成,其优势种为甘青针茅、白莲蒿和甘菊;火烧地出现30种植物,其优势种为白莲蒿、甘青针茅和干生苔草。可见,不同的干扰方式明显改变了草地群落的优势种,在放牧地干生苔草、百里香和猪毛蒿表现出相对较好的优势性,出现频率最高;在封育地,干生苔草和百里香的出现频率降低,猪毛蒿不再出现,而甘青针茅和白莲蒿的优势地位突显,草地群落已恢复成以多年生禾草为优势种的群落;火烧干扰降低了甘青针茅的出现频率,提高了白莲蒿的出现频率。因此,封育、放牧和火烧干扰对种群的存在状况以及群落的结构产生了一定的影响。结果表明,群落的组成结构和种群的优势地位均会随着干扰方式的不同而发生一定程度地改变。

表1 不同干扰方式下云雾山典型草原群落的物种组成特征Table 1 Characteristics of community species composition in typical steppe of Yunwu Mountain under different disturbance regimes
注:“-”代表物种不存在
Note: No species represented by“-”.
3.2 不同干扰方式下种群空间异质性的变化
3.2.1 不同干扰方式下种群空间分布的幂函数规律 放牧、封育和火烧干扰下云雾山典型草原群落物种随机分布与实际分布的变化趋势如图1所示。图1显示,放牧、封育和火烧样地群落的实际分布曲线均位于随机分布曲线的上方,可见这3类草地群落物种均呈集群分布,说明相对于随机分布而言,3类样地的群落在实际分布中都偏离了随机分布,且在整体上具有较强的空间异质性。放牧、封育和火烧3类样地的决定系数R2依次是0.960 8,0.896 8和0.916 9,均接近于1,表明这3类样地中并没有不稳定种群使群落的结构产生混乱,即种群的分布与幂函数法则基本吻合。

直线y=x为种群的随机分布曲线,直线y=ax+b(a,b均为常数)为实际分布情况The straight line y=x is the random distribution curve of population,and the straight line y=ax+b(a,b is constant) is the actual distribution图1 不同干扰方式下云雾山典型草原群落物种随机分布与实际分布的变化趋势Fig.1 Change of species random and actual distribution in typical steppe of Yunwu Mountain under different disturbance regimes
3.2.2 不同干扰方式下物种出现频率与种群空间异质性指数的比较 物种出现频率p和种群空间异质性指数δi共同决定着种群分布的密集程度,也就是指种群在群落整体上所起的作用。出现频率较高且空间异质性指数也高的种群,对于群落的空间异质性具有正向促进作用;相反,出现频率较低且空间异质性指数也较低的种群,则会在一定程度上降低群落的空间异质性。不同干扰方式下云雾山典型草原物种出现频率与种群空间异质性指数之间的关系如图2所示。

物种编号参见表1 Species No.refer to Table 1图2 不同干扰方式下云雾山典型草原物种出现频率与种群空间异质性指数之间的关系Fig.2 Relationship between occurrence frequency and spatial heterogeneity index in typical steppe of Yunwu Mountain under different disturbance regimes
由图2可以看出,放牧和火烧地的物种在群落整体中的分布情况较封育样地而言相对集中,植物大多呈现“密集型”分布,如出现频率相对较高的物种干生苔草呈现“密集型”分布;此外,出现频率相对较低的物种翼茎凤毛菊则呈“疏散型”分布。因此,研究不同干扰方式下各群落物种的出现频率与空间异质性指数的关系,可以体现出其不同功能特性的异质性。群落整体的空间异质性指数由大到小依次为放牧地(1.258 6)>火烧地(1.002 1)>封育地(0.485 3)(表2),结果表明,放牧和火烧干扰提高了群落的空间异质性,而封育措施使群落的均匀性较好。
许多物种在不同的干扰方式下出现频率发生明显变化,并表现出各自的空间特性。在放牧地,物种甘青针茅、干生苔草、百里香、猪毛蒿、阿尔泰狗娃花、星毛委陵菜、散穗早熟禾和鳞叶龙胆的空间异质性指数均高于群落空间异质性指数,说明这些物种对群落的空间异质性指数影响较大;物种百里香和干生苔草的出现频率和空间异质性指数均较大,在小尺度内容易形成斑块化或小型群落,对群落空间异质性也起着非常重要的影响;物种甘青针茅、多叶棘豆和野韭在放牧地出现频率较小,但空间异质性指数较大;出现频率与空间异质性指数均较小的多毛并头黄芩仅仅增加了物种多样性,而对群落的空间异质性影响可以忽略。在封育地,有11个物种的空间异质性指数高于群落空间异质性指数,分别为星毛委陵菜、甘菊、百里香、柴胡、阿尔泰狗娃花、蓬子菜、西山委陵菜、干生苔草、多毛并头黄岑、茅香和披碱草,其中甘菊的出现频率和空间异质性指数均较大;出现频率较小而空间异质性指数较大的物种有星毛委陵菜和柴胡;出现频率较大而空间异质性指数较小的物种为白莲蒿和甘青针茅;出现频率和空间异质性指数均较小的物种有翼茎凤毛菊、大针茅、散穗早熟禾和野韭,其对群落空间异质性影响作用不明显。在火烧地,种群空间异质性指数高于群落空间异质性的物种有12种,分别为百里香、狼毒、星毛委陵菜、白莲蒿、甘菊、二裂委陵菜、蓬子菜、披碱草、阿尔泰狗娃花、茅香、多叶棘豆和干生苔草,其中干生苔草和白莲蒿的出现频率与空间异质性指数均较高;物种多叶棘豆、星毛委陵菜、蓬子菜、狼毒和阿尔泰狗娃花出现频率较小,空间异质性指数较大;甘青针茅出现频率较大,空间异质性指数较小;空间异质性较小且出现频率较低的物种为中华委陵菜、散穗早熟禾、翼茎凤毛菊和鳞叶龙胆,其降低了群落空间异质性,使群落具有良好的空间结构,进而能够更好地维持群落各物种相对稳定的生长态势。
3.3 干扰方式对物种多样性的影响
方差分析结果表明,封育、放牧和火烧干扰对物种丰富度、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(E)影响显著(P<0.05),而对Simpson多样性指数(D)影响不显著(P>0.05)(表2)。物种丰富度和Shannon-Wiener多样性指数在放牧地和火烧地之间差异不显著,但在封育地显著降低;Pielou均匀度指数在放牧地和火烧地显著低于封育地,说明火烧和放牧干扰将导致物种均匀度下降,这可能是火烧和放牧使地表长时间裸露导致土壤水分损失较多,对植物的生长产生不利影响。

表2 放牧、封育和火烧不同干扰方式下云雾山典型草原植物群落的多样性Table 2 Diversities of plant communities in grazed,fenced and burned grasslands in typical steppe of Yunwu Mountain
4 讨 论
空间格局始终是生态学研究的核心问题,而在生态系统中,空间异质性则是形成格局的主要原因[14]。许多生物因子(如竞争、牲畜采食、人为采伐等)和非生物因子(地貌、温度、水分等)分布的不规则性和复杂性,导致了植被分布的空间异质性[15]。目前,植被空间异质性与环境因子之间关系的研究是国内外学者对空间异质性研究的主要内容之一。了解不同干扰因素下群落植被的空间分布格局,能够更好地对其在不同干扰因素下的空间分布规律以及各群落种群的消长过程进行分析[13]。本研究表明,在云雾山典型草原,封育地、放牧地和火烧地群落中的种群空间分布格局均偏离随机分布曲线y=x,物种都呈集中分布,表现出较强的空间异质性。在草地生态系统中,种群的空间分布格局几乎都偏离随机分布。在内蒙古克氏针茅(Stipakrylovii)草原,物种在围栏内外均偏离随机分布而呈集中分布[11]。在内蒙古典型草原,星毛委陵菜在不同放牧强度下小尺度空间格局均呈种内聚集分布[17]。在不同干扰条件下,群落均呈现集中分布,这可能与植物自身的生物学特性和个体的自我保护补偿机制有关[11]。
本研究表明,不同干扰方式显著影响草原植被群落物种的空间异质性。在该区,基于幂函数规律的回归曲线y=ax+b的相关性系数在放牧地、封育地和火烧地均较高,依次为0.960 8,0.896 8和0.916 9,与在其他草地类型的研究结果一致[6,10],说明幂函数法可以运用到典型草原群落中。另外,与封育措施相比,放牧和火烧干扰均明显提高了草地群落的空间异质性指数,说明封育地的空间异质性低于放牧地和火烧地,物种在无外界干扰因素影响下均匀分布较强,而集群生长能力相对于放牧地和火烧地弱,这与蒋德明[15]的研究结果一致,其原因可能与封育地缺乏牲畜的采食、动物的践踏和人为干扰有关。本研究中封育草地的围封时间为23年,长期排除了人为干扰和放牧牲畜的践踏与采食,形成了以多年生禾草为优势的群落,群落枯落物大量积聚,土壤结构在一定程度上也发生了变化,生境更加均匀一致,因此封育地群落植被的随机分布增强,共存物种减少。放牧干扰增强了群落的空间异质性,这可能与放牧牲畜的采食和践踏有关。刘红梅等[16]研究指出,植物通过株丛矮化与扩大种群所占据的空间来适应各种外界因素的干扰。一方面,放牧行为可为一些耐牧的牧草提供拓殖的空间,它们经过自身可塑性特征的调节,更倾向于通过无性繁殖来进行种群更新[18]。已有许多研究表明,克隆生长可以促进植物获取环境异质性资源[19-22],植物的无性生长通过觅食(foraging)行为适应资源异质性[23]。另一方面,放牧活动产生的空地斑块的空间分布方式进一步决定了牧草的空间格局,耐牧牧草在小范围内密集呈小斑块以增强种群竞争力。另外,同一物种个体之间以及与其他植物之间的种内种间关系(竞争),在一定程度上决定了物种种群集中分布的空间格局。耐牧牧草为了避免动物的啃食,植株个体矮化和小型化,种内竞争减弱,个体间距缩小,易于形成集中分布来分担风险。同样在火烧地,一些不耐火植物经历高温之后灭绝,而生存下来的耐火的植物更倾向于通过无性繁殖来形成聚集的群体,这种集群分布更有利于提高个体的生存和竞争力 , 有利于种群的迅速扩散和传播。可见,对长期封育的草地进行适度的放牧和火烧干扰,有利于增强群落的空间异质性,形成更加多样的小生境。群落物种的异质化分布,易于形成有利于对抗不利影响因素干扰的空间生态位,能允许更多的物种共存,提高种群的存活率,维持物种多样性。另外,不同干扰方式也会改变种群在群落中的贡献地位和作用。放牧干扰下的群落优势种为干生苔草、散穗早熟禾、猪毛蒿和百里香;封育干扰下的群落优势种为甘青针茅、甘菊、白莲蒿和披碱草;火烧干扰下的群落优势种为白莲蒿、甘青针茅和干生苔草。群落的空间异质性状况还受干扰频度和强度的影响,它们决定着各群落动态变化过程中群落在空间序列的位移程度[14]。
受不同干扰因素的影响,草地群落中的种群空间分布也表现出不同程度的异质性[24]。在本研究区,决定群落空间分布异质性的物种可分为种群空间异质性指数低于、接近和高于群落空间异质性指数的3类物种。不同物种在不同干扰因素下表现出的空间差异性具有物种特异性。一些物种如白莲蒿种群的空间异质性指数在封育地小于群落空间异质性指数,在放牧地接近群落空间异质性指数,而在火烧地却明显高于群落空间异质性指数,表明白莲蒿的空间异质性在不同干扰方式下具有物种特异性,这可能与其自身的生物学特性和对生态环境的适应有关[25]。此外,一些物种如干生苔草在放牧地和火烧地的出现频率和空间异质性指数均较高,表明干生苔草种群在群落中分布较密集,具有相对较大的斑块分布;而另外一些物种如散穗早熟禾在封育和火烧地的出现频率和空间异质性指数均较低,说明这些物种在封育和火烧草地群落中分布较为稀疏,其斑块分布较小。尽管斑块大小对于提高种群空间异质性的贡献和作用有所不同,但植被的出现频率和空间异质性指数均能够明确地体现出种群在群落中所起的意义与作用[13]。
物种多样性与物种的空间分布特征密切相关,是物种分布对环境的依赖性和群落功能变异性的综合反映[26]。在该区,封育显著降低了群落的Pielou均匀度指数。封育措施排除了家畜的啃食行为,提高了甘青针茅等多年生高大禾草的有性繁殖能力,缩小了空地斑块,从而降低了植物对生境的分割和群落的空间异质性。
5 结 论
在封育、放牧和火烧草地,基于幂函数规律的种群实际分布曲线都位于理想曲线y=x的上方,植被呈现集中分布的特点,表现出比随机分布更强的空间异质性。在黄土高原典型草原,基于幂函数规律的种群分布与幂函数法则具有良好的吻合性,可见幂函数法则可以运用到典型草原植被空间异质性的研究中。封育措施降低了群落的空间异质性,而放牧和火烧干扰增强了草地群落的空间异质性。
猜你喜欢
当代水产(2022年8期)2022-09-20
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
绿色科技(2021年10期)2021-06-23
林业科技(2020年3期)2021-01-21
河南科学(2020年3期)2020-06-02
南方周末(2019-06-27)2019-06-27
小学生作文(低年级适用)(2018年12期)2018-03-23
