
镉胁迫对尾穗苋种子萌发及幼苗生理特性的影响
2018-08-14 12:59,,,,
种子 2018年7期
, , , ,
(贵州大学 生命科学学院, 贵阳 550025)
近年来,我国工业发展非常猛速,对自然环境的污染也随之加剧。相关调查显示,目前我国重金属超标点位占全部超标点位的82.8%,其中最为严重的是镉污染,超标率达7.0%[1]。重金属污染具有长期性、隐蔽性和不可逆性等特点,寻求环保、高效、经济等解决方案已迫在眉睫[2]。在重金属胁迫下衡量植物耐受性的一个重要指标是植物长势。同时植物吸收重金属主要取决于重金属在培养液中存在的形态,其中容易被植物吸收的形态会对植物的生长产生一定影响[3]。重金属污染会降低生物多样性并影响多种重要经济作物和农作物的产量和质量,已成为全球面临的重要环境问题[4]。重金属污染土壤修复治理是近年来国际上兴起的植物修复新技术[5]。利用某些植物对重金属具有超富集能力,能将重金属富集到自身体内[6],达到修复土壤的目的。植物修复具有三个明显优势:一是适用范围广[7];二是可实施原位修复;三是投入成本较低。因而,对重金属污染的治理需要植物修复技术,植物修复技术可美化环境,又可使被污染的土壤得以再利用。
尾穗苋(Amaranthuscaudatus)是苋科(Amaranthaceae)苋属(AmaranthusL.)一年生草本植物,抗性强、易生长、耐旱、耐湿、耐高温,病虫害很少发生,叶杂有各种颜色可供观赏。研究表明,尾穗苋为超积累镉特征植物,对镉等重金属阳离子具有很强的抗性,可作为当今重金属污染植物修复的修复植物。本试验以尾穗苋为材料,采用不同浓度的镉胁迫处理,通过观察镉对植物生长的影响和生理指标的变化,研究镉胁迫对尾穗苋生理生化的影响,为重金属污染植物复合修复研究提供参考。
1 试验材料与方法
1.1 材 料
尾穗苋种子来源于贵州省贵阳市花溪区生态园,胁迫试剂氯化镉(CdCl22.5 H2O)为分析纯。试验在贵州大学生物技术试验室进行。
1.2 方 法
1.2.1 水培法
挑选大小均一的尾穗苋种子,用70%酒精消毒3~5 min后用蒸馏水反复冲洗干净,在已灭菌培养皿中进行播种,选取直径为9 cm的培养皿,并在皿内垫2张直径为9 cm的滤纸作为发芽床,每皿放入50粒种子。等量的Cd2+溶液(浓度分别为10,50,100,200,500 mg/L)(按氯化镉中Cd2+计算)按编号加入培养基中。将种子均匀地放入培养基,种子间要有一定的间距,用蒸馏水作空白对照,每个浓度设置3个重复,处理完后将培养皿放在(25±2)℃光照恒温培养箱中培养。每天补充等量相应浓度的溶液,空气湿度要保持在一定水平。定期记录每个培养皿中种子的生长发芽状况。
1.2.2 土培法
挑选大小均一的尾穗苋种子,用70%酒精消毒3~5 min后用蒸馏水反复冲洗干净,取口径为21 cm的塑料盆,盆中铺上滤纸,再放入等量的营养土;将配好的各浓度镉溶液等量依次加入对应编号的塑料盆中搅拌。将处理后的种子播种在土壤里,每盆放入50粒种子。播种后用少许土将种子掩盖,用蒸馏水作空白对照,每个浓度设置3个重复,用保鲜膜封住盆口,置入(25±2)℃光照恒温培养箱中培养,保持一定的空气湿度。定期观察记录每个塑料盆中种子的生长发芽状况。每隔1~2 d浇1次蒸馏水。定期观察尾穗苋出苗情况。
1.3 种子萌发及幼苗生理指标的测定
1.3.1 种子萌发指标的测定
尾穗苋种子水培萌发后,第7天开始测定其根长和苗长,并统计种子的发芽率[9],发芽率(%)=试验种子有效的发芽数/试验种子总数×100%,用数显游标卡尺测量根长和苗长,每个处理分别测量20株。
1.3.2 幼苗生理指标的测定
第40天测量尾穗苋幼苗的各项生理指标,叶绿素含量采用95%的酒精抽提[10];丙二醛(MDA)含量采用硫代巴比妥酸法测定[11];过氧化物酶(POD)活性采用愈创木酚法测定[12];超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)光化还原法测定[13];可溶性蛋白含量测定采用考马斯亮蓝G 250染色法[14];可溶性糖含量和淀粉含量测定采用蒽酮比色法[14];脯氨酸含量采用茚三酮法测定[14]。
1.4 试验数据分析
本次试验数据采用Microsoft Excel 2010进行处理,并通过SPSS 21.0统计软件对数据进行方差分析和相关分析。
2 结果与分析
2.1 镉胁迫对尾穗苋种子发芽率的影响
由图1可见,尾穗苋种子的发芽率在90%~98.90%范围内;镉浓度为0~50 mg/L时,随着镉浓度的增加,发芽率逐渐增大,表明在此浓度范围内镉胁迫对种子的萌发有促进作用,并且当镉浓度为50 mg/L时种子发芽率最高,达98.90%;当镉浓度为50~100 mg/L时,发芽率急剧下降,在镉胁迫浓度为100,200,500 mg/L时,发芽率分别为93.33%、91.10%、90.00%,均抑制尾穗苋种子的萌发。表明低浓度镉胁迫处理促进了尾穗苋种子的萌发,而高浓度镉胁迫处理则抑制了尾穗苋种子萌发。
2.2 镉胁迫对尾穗苋幼苗长势的影响
由图2可见,尾穗苋的根长在2.05~46 mm之间;无镉胁迫下尾穗苋根长势最好,根长达46 mm;随着镉浓度的增大,根长逐渐变短,在镉浓度为500 mg/L时根长最短,为2.0 mm;当镉浓度为10 mg/L以后,其镉浓度处理的根长与对照组(0 mg/L)相比有显著差异的抑制作用(p<0.05);但镉浓度超过200 mg/L后,各浓度所对应的根长差异不显著。表明镉胁迫对尾穗苋幼苗根的生长具有明显的抑制作用。

图1 镉胁迫对尾穗苋种子发芽率的影响
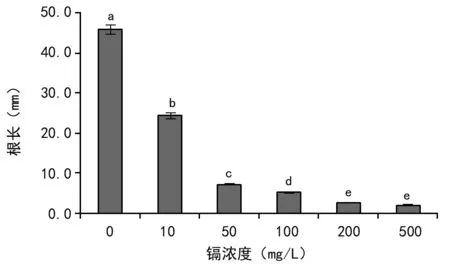
注:不同字母表示0.05水平差异显著。下同。图2 镉胁迫对尾穗苋幼苗根长的影响
由图3可知,尾穗苋的苗长为4.23~62.2 mm;无镉胁迫下尾穗苋苗长长势最好,苗长为62.2 mm;随着镉浓度的升高,对尾穗苋苗长的抑制作用逐渐增强,当镉浓度为500 mg/L时苗长最短,为4.23 mm。不同镉浓度处理之间的差异极显著(p<0.01)。表明镉胁迫对尾穗苋幼苗苗长均具有显著的抑制作用。

图3 镉胁迫对尾穗苋幼苗苗长的影响
2.3 镉胁迫对尾穗苋幼苗SOD活性的影响
超氧化物歧化酶(SOD)是以氧自由基为底物的一种酶,其在活性氧代谢中具有重要地位,为生物体内最重要的清除活性氧自由基的酶类[16]。由图4可知,尾穗苋的SOD活性范围在41.704~382.479 U/g(FW)之间;无镉胁迫(对照组)时SOD活性最高,为382.479 U/g(FW);当镉浓度为10~500 mg/L时,SOD活性均低于对照组,而镉浓度为10 mg/L时,尾穗苋幼苗SOD活性最低,为41.704 U/g(FW),除了浓度为50 mg/L时的SOD活性与对照组差异不显著外,其他均呈现显著性差异。表明镉胁迫对尾穗苋幼苗SOD活性均具有抑制作用。

图4 镉胁迫对尾穗苋幼苗SOD活性的影响
2.4 镉胁迫对尾穗苋幼苗POD活性的影响
过氧化物酶(POD)是植物体内保护酶,其在抗逆境中起着关键作用[17]。由图5可以看出,尾穗苋的POD活性范围在851.185~2 666.667 U/g(FW)之间;镉浓度为10 mg/L时,POD活性最高,达2 666.667 U/g(FW);而镉浓度超过50 mg/L后,对POD有显著的抑制作用(p<0.05)。显示低浓度(10 mg/L)镉胁迫增加了尾穗苋幼苗的POD活性,而高浓度的镉胁迫降低了尾穗苋幼苗的POD活性。

图5 镉胁迫对尾穗苋幼苗POD活性的影响
2.5 镉胁迫对尾穗苋幼苗可溶性蛋白含量的影响
由图6可见,随镉胁迫浓度的增大尾穗苋幼苗可溶性蛋白质的含量先增后减,尾穗苋的可溶性蛋白质含量范围在0.600~7.75 0 mg/L之间,在镉胁迫浓度为10 mg/g时,尾穗苋幼苗可溶性蛋白质的含量最高,为7.75 mg/g。在10~100 mg/L浓度范围内尾穗苋幼苗可溶性蛋白质的含量明显下降,从7.750 mg/g下降到0.625 mg/g;而在100~500 mg/L浓度范围内可溶性蛋白质的含量变化不大;当镉浓度为500 mg/L时,可溶性蛋白质含量最低,为0.600 mg/g。试验结果表明,在镉胁迫浓度小于50 mg/L时,尾穗苋幼苗可溶性蛋白质含量保持在较高的水平,而镉胁迫浓度大于100 mg/L以后,可溶性蛋白质含量保持在较低的水平。

图6 镉胁迫对尾穗苋幼苗可溶性蛋白质含量的影响
2.6 镉胁迫对尾穗苋幼苗可溶性糖含量的影响
由图7可知,尾穗苋幼苗可溶性糖含量范围在0.014 3%~0.022 9%之间,镉浓度为100 mg/L时尾穗苋幼苗可溶性糖含量最大,为0.022 9%,但镉浓度为10 mg/L时达到最小值,为0.0143%;在0~10 mg/L和100~200 mg/L浓度范围内,尾穗苋幼苗的可溶性糖含量都在减少,在10~100 mg/L和200~500 mg/L浓度范围内可溶性糖含量都在增多。

图7 镉胁迫对尾穗苋幼苗可溶性糖含量的影响
2.7 镉胁迫对尾穗苋幼苗淀粉含量的影响
由图8可知,尾穗苋幼苗淀粉的含量范围在21.857%~45.000%之间,无镉胁迫时淀粉含量达到最大值,为45%;随着镉胁迫浓度的增大,可溶性淀粉的含量逐渐降低,在镉浓度为200 mg/L时达到最小值,为21.857%。

图8 镉胁迫对尾穗苋幼苗淀粉含量的影响
2.8 镉胁迫对尾穗苋幼苗叶绿素含量的影响
由图9可见,尾穗苋幼苗叶绿素含量范围在0.52~0.81 mg/g(FW)之间,镉胁迫对尾穗苋幼苗叶绿素含量影响不明显,它们之间的差异不显著。试验结果表明,镉浓度为10~500 mg/L时对尾穗苋幼苗叶绿素含量影响不明显。

图9 镉胁迫对尾穗苋幼苗叶绿素含量的影响
2.9 镉胁迫对尾穗苋幼苗MDA含量的影响
脂质氧化终产物丙二醛(MDA)是膜脂过氧化最重要的产物之一[18];通过MDA了解膜脂过氧化的程度,间接测定膜系统受损程度以及植物的抗逆性[19]。由图10可知,尾穗苋幼苗MDA含量范围在1.610~2.603 mol/g(FW)之间;镉浓度为10 mg/L时MDA含量最高,为2.603 mol/g(FW);但在镉浓度为50 mg/L时MDA含量最低,为1.610 mol/g(FW)。表明在镉浓度为0~50 mg/L区间内,MDA含量先增加后降低;50~100 mg/L区间内MDA含量缓慢升高。
2.10 镉胁迫对尾穗苋幼苗游离脯氨酸含量的影响
脯氨酸(Pro)是植物蛋白质的重要组分之一,Pro含量在一定程度上能反映植物在逆境中的抗逆性。由图11可知,尾穗苋幼苗Pro含量范围在63.08~97.59μg/g(FW)之间;镉浓度为10 mg/L时Pro含量达到最高,为97.59μg/g(FW);在镉浓度为500 mg/L时Pro含量最低,为63.08μg/g(FW)。表明0~500 mg/L浓度镉胁迫对尾穗苋幼苗Pro含量的影响不明显,差异不显著。
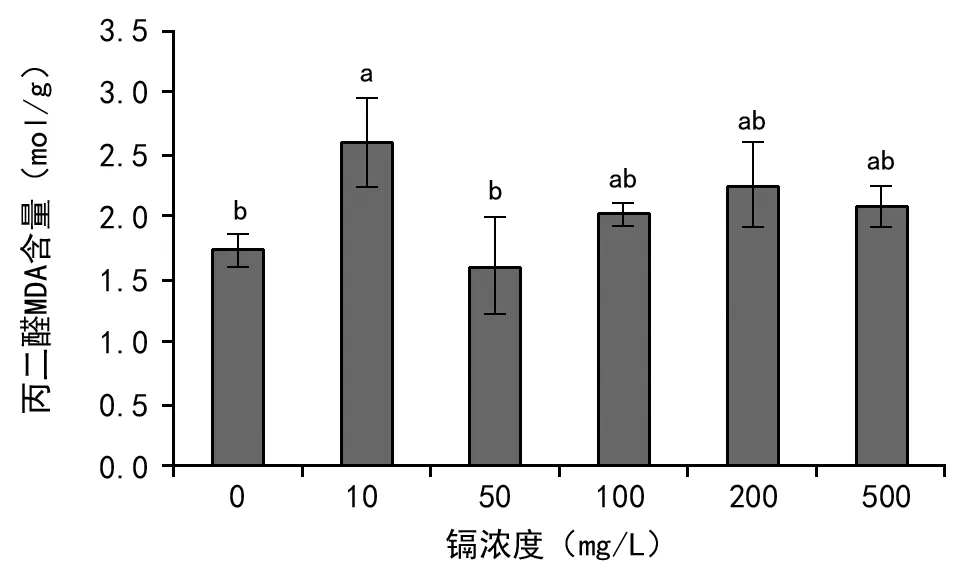
图10 镉胁迫对尾穗苋幼苗MDA含量的影响

图11 镉胁迫对尾穗苋幼苗游离脯氨酸含量的影响
3 讨 论
3.1 不同浓度镉胁迫对尾穗苋种子萌发的影响
有报道显示,重金属对植物种子具有低促高抑的作用[20]。在姚焱等[21]研究中,低浓度铊胁迫并未对白菜种子发芽率和发芽势产生显著影响,但高浓度铊胁迫下植物生长受到显著抑制,表现为随培养时间延长胚根伸长减缓。Kaplan等[22]研究了铊毒胁迫大豆生长情况。本研究得到相似结果,随着重金属镉浓度的增加,尾穗苋种子萌发率呈现出先增后降的趋势,可能是由于较高浓度的镉影响了尾穗苋植物正常代谢,从而抑制了尾穗苋的萌发。
3.2 不同浓度镉胁迫对尾穗苋幼苗生长的影响
研究表明,重金属物质进入植物体后,与植物体内的大分子结合,大部分累积在植物的根部,仅少量向地上部分迁移[23]。孙圆圆等研究表明,高浓度胁迫下Cd2+明显抑制一年生黑麦草根和芽的生长[24]。杨鹏等认为,镉胁迫抑制了紫英幼苗的生长[25]。殷欣研究表明,各处理浓度的镉胁迫均导致大豆幼苗出现不同程度的毒害症征,抑制了大豆的生长[26]。本研究得到相似结果,镉胁迫对尾穗苋幼苗的根和苗的生长都有抑制作用,尾穗苋幼苗的长势随着镉浓度的升高而被抑制越明显。不同镉浓度对尾穗苋幼苗处理后,苗长差异极显著。
3.3 不同浓度镉胁迫对尾穗苋幼苗生理生化的影响
植物本身能够使体内ROS的产生与清除维持动态平衡,但在受到重金属胁迫后这种平衡会被打破[27]。重金属胁迫导致植株组织的氧化胁迫和膜损伤,改变酶系活性,这种损伤对植株生长产生一定的影响[28]。植物通过POD和SOD的抗氧化酶系统来提高植株对逆境的耐抗性作用[29]。本试验对SOD、POD、可溶性蛋白质、可溶性糖、淀粉、叶绿素、丙二醛(MDA)和游离脯氨酸含量进行测定,结果发现,不同镉浓度对尾穗苋幼苗处理后,叶绿素含量和脯氨酸含量差异不显著,这可能与本试验取材时期有关,有关各时期的具体变化情况,有待进一步研究证明;可溶性蛋白质含量、丙二醛(MDA)含量及POD活性随镉浓度的增加呈现出先增后降,由此表明植物仅在一定范围的逆境中具有抵抗能力,这一范围一旦被超过,植物自身的抵御能力就会失去,这与徐玲玲等[30]的研究结果一致;可溶性糖含量、淀粉含量和SOD活性随镉浓度的增加呈现出先降后增,可见植物在逆境中有一定的适应时间段,而后植株通过自身的调剂机制进行综合调控,以减轻逆境对本身带来的伤害。由于植物对重金属的吸收和转运机理很复杂,本试验不同镉浓度胁迫尾穗苋种子萌发和幼苗初期生理特性的影响表明,对镉耐受性存在差异,其原因有待进一步分析。
猜你喜欢
农业灾害研究(2022年2期)2022-05-31
今日农业(2021年10期)2021-11-27
昆明医科大学学报(2021年8期)2021-08-13
矿产综合利用(2020年1期)2020-07-24
天然产物研究与开发(2019年1期)2019-03-01
中成药(2017年6期)2017-06-13
中国火炬(2015年12期)2015-07-31
医学研究杂志(2015年9期)2015-07-01
食品工业科技(2014年23期)2014-03-11
表面工程与再制造(2014年2期)2014-02-27
