
亲和人体与非亲和人体双歧杆菌之间的生理特征差异
2018-08-29 07:59黄勤雯陈铁涛肖金忠
中国乳品工业 2018年6期
黄勤雯,陈铁涛,肖金忠
(1.森永乳业株式会社基础研究所,座间市神奈川县228-8583,日本;2.百施(上海)生物科技有限公司上海,200436)
0 引 言
双歧杆菌是人类肠道菌群中主要的有益菌种之一[1]。大量的研究证实双歧杆菌在肠道内可起到抑制有害菌的增值、刺激人体免疫系统,从而提高抗病能力等功能[2-3]。有鉴于此,近年来一部分双歧杆菌的菌种(比如长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、两歧双歧杆菌、乳双歧杆菌等)被广泛地应用于医药品以及各种食品,其中包括发酵乳、婴儿配方奶粉和营养保健食品[4-5]等。
本文旨在深入了解不同寄宿起源的双歧杆菌的基因型和生理特征的差异。双歧杆菌的比较基因组学研究揭示了不同寄宿起源的双歧杆菌在人体肠道的适应性的不同。此外,生理学方面以及临床研究也显示不同寄宿起源的双歧杆菌对人体的健康促进作用的差异。
1 双歧杆菌种类及分布
双歧杆菌是一种革兰阳性、专性厌氧、非运动性、无芽孢的杆菌。它们最初是1890年由H enry Tissier从母乳喂养的婴儿粪便分离得到,根据其独有的不规则Y型形态,Tissier将其命名为分叉杆菌[6]。尽管1924年O rla-Jensen提议将其独立分类为双歧杆菌属[7],但由于它们与乳酸杆菌的相识性,在相当长的期间里双歧杆菌常被归类为乳酸杆菌属(Lactobacillus)。直到20世纪中期,双歧杆菌才被分类为双歧杆菌属(Bifidobacterium)。在分类学上双歧杆菌归属于细菌界、放线菌门、放线菌纲、双歧杆菌目、双歧杆菌科、双歧杆菌属。双歧杆菌的形态多变,常因种及生长环境不同而呈现弯曲杆形、Y或V形等多种形态。
双歧杆菌广泛分布于不同的生态环境,其中包括人体肠道、口腔、部分热血动物的消化道、粪便、血液和食物中[8]。目前,随着分子生物学的发展和各项技术的应用,对已发现的双歧杆菌属分类成58个种和10个亚种(LPSN,http://www.bacterio.net)[9]。一般来说,定殖在人体肠道和口腔内的双歧杆菌被归类为亲和人体双歧杆菌(人类寄宿性双歧杆菌,Human-Residential Bifidobacteria,HRB)(图 1)[10-11]。双歧杆菌作为人类肠道菌群的重要组成部分,随着婴儿出生后迅速增殖并成为优势菌种,特别是在母乳哺育的婴儿肠道里双歧杆菌占有很大的比率。断奶以后肠道中的双歧杆菌的数量开始下降,特别是在高龄者的肠道中其数量更加减少[12-13]。
HRB可分为婴儿型HRB和成人型HRB,其中普遍分布于婴儿肠道的婴儿双歧杆菌(B.longumsubsp.infantis)、长双歧杆菌(B.longumsubsp.longum)、短双歧杆菌(B.breve)和两歧双歧杆菌(B.bifidum)被归类为婴儿型HRB[14,15],而主要分布于成人肠道的菌种如青春双歧杆菌(B.adolescentis)、假链双歧杆菌(B.pseudocatenulatum)、链状双歧杆菌(B.catenulatum)、长双歧杆菌(B.longumsubsp.longum)等则被归类为成人型HRB[16]。值得一提的是,长双歧杆菌是唯一普遍分布于婴儿及成人的肠道内的菌种[17]。
此外,定殖于动物肠道或其他生态环境的双歧杆菌被归类为非亲和人体双歧杆菌(非人类寄宿性双歧杆菌,N on-Human-Residential Bifidobacteria,非HRB),其中包括动物双歧杆菌(B.animalissubsp.animalis)、乳双歧杆菌(B.animalissubsp.lactis)、嗜热双歧杆菌(B.thermophilum)、假长双歧杆菌(B.pseudolongum)等(图 1)[10-11]。其中,有报道表明普遍应用于各种乳制品的乳双歧杆菌亦可从人的粪便中分离得到。但很多研究表明其分离与人们在饮食中摄取该菌有关系,因此乳双歧杆菌被视为非自然寄宿于人体的菌种[18-19]。

图1 亲和人体双歧杆菌(HRB)与非亲和人体双歧杆菌(非HRB)的分布
2 HRB与非HRB之间生理特征差异
基因组学的研究发现HRB与非HRB之间在基因构成上有着很多差异,揭示了双歧杆菌的起源与其生理特征方面的关系[20-21]。
2.1 叶酸合成
叶酸是一种对人体有着重要的生理功能的水溶性维生素,其备受关注的最早原因是叶酸缺乏对胎儿及婴幼儿神经系统发育的影响;随着研究深入,发现叶酸还具备抗肿瘤,防治心脑血管疾病及肠道疾病的功能[22-23]。然而人类等高等动物却由于缺乏相应的叶酸合成基因而不能合成自身所需的叶酸,而必须通过饮食摄入或依靠肠道菌群所产生的叶酸来供给。研究证实肠道菌群中的双歧杆菌可以产生多种维生素包括叶酸,是宿主维生素的主要来源之一,然而不同的菌种的叶酸合成能力有着显着的差异[24-26]。
基因组学的研究发现双歧杆菌的叶酸代谢基因与菌株起源性有相关性[20,21]。双歧杆菌的叶酸合成代谢途径包括蝶呤代谢途径(Pterin branches)及氨基苯甲酸代谢途径(pABA branches)(见图2),这二条代谢途径只有同时发挥作用才能促成叶酸合成。基因组学研究发现HRB菌株普遍具备完整的从头合成叶酸的途径,然而非HRB菌株如动物双歧杆菌和乳双歧杆菌则缺乏合成6-羟甲基-7,8-二氢喋呤焦磷酸(DHPPP)的关键基因以及结合DHPPP和氨基苯甲酸(pABA)的二氢叶酸合成酶(dihydroptetroate synthase)的基因,从而导致非HRB不能有效地从头合成叶酸[20-25](图2)。

图2 基于基因组学所预测的亲和人体双歧杆菌(HRB)和非亲和人体双歧杆菌(非HRB)的叶酸合成途径
图2中,部分非HRB菌株如动物双歧杆菌和乳双歧杆菌缺乏合成6-羟甲基-7,8-二氢喋呤焦磷酸(DHPPP)的6-羟甲基-7,8-二氢叶酸激酶(HPPK),以及结合DHPPP和氨基苯甲酸(pABA)的二氢叶酸合成酶(dihydropteroate synthase),从而导致该类双歧杆菌不能有效地从头合成叶酸。
试管实验发现大部分HRB菌株能够在不含有叶酸的培养基或含有一定叶酸水平的培养基中增殖并产生叶酸[26-27]。然而,只有少数的非HRB菌株能产生少量的叶酸[20,27]。Sugahara等研究了44个双歧杆菌菌株的叶酸合成能力,发现所有HRB菌株培养后均含有高于培养基所含有的初始水平的叶酸,与之相反,非HRB菌株的叶酸浓度则相对地低于培养基所含有叶酸的初始水平[26](图3)。另有研究发现一些从非人类的灵长类动物中分离到的HRB菌株,如青春双歧杆菌和齿双歧杆菌(B.dentium)亦能产生高量的叶酸[24]。相比之下,非HRB,如动物双歧杆菌、乳双歧杆菌、假长双歧杆菌、星状双歧杆菌(B.asteroides)、棒状双歧杆菌(B.coryneforme)和蜜蜂双歧杆菌(B.indicum)均不能合成叶酸[24]。
图3中,显示双歧杆菌培养后的上清液中的叶酸浓度,虚线为培养基中的初始叶酸水平。
动物实验也证实了HRB菌株不需要与其他肠道菌群相互作用即能直接合成叶酸[26]。将HRB或非HRB的单菌株移植到无菌小白鼠的肠道中发现,长双歧杆菌单独定植的无菌小白鼠粪便中的叶酸浓度高于乳双歧杆菌单独定植的小白鼠[26]。因为叶酸与贫血有关,该研究还发现定植有长双歧杆菌的无菌小白鼠的血中的血红蛋白浓度高于无菌小白鼠或定植有非HRB菌株的小白鼠。有鉴于此,具备叶酸合成能力的HRB还可能在预防贫血方面给宿主带来健康益处。
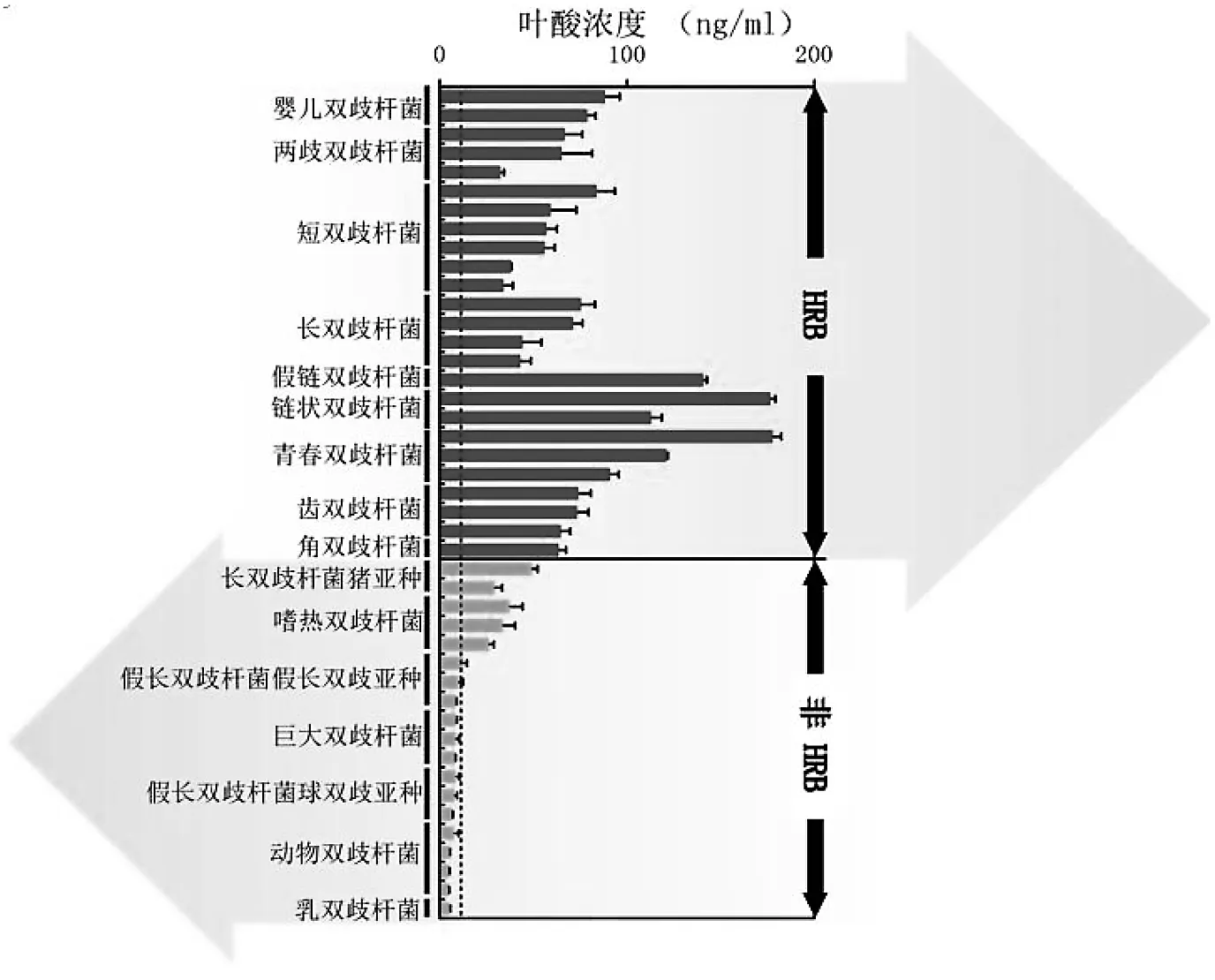
图3 亲和人体双歧杆菌(HRB)和非亲和人体双歧杆菌(非HRB)的叶酸合成能力
另外,Pompei等研究了在缺乏叶酸饮食中添加双歧杆菌对改善Wistar大白鼠叶酸水平的效果。研究表明添加HRB菌株,青春双歧杆菌DSM Z 18352、青春双歧杆菌DSM Z 18350和假链双歧杆菌DSM Z 18353均能有效地改善大白鼠的叶酸水平[28]。为此,Pompei等还进行了一项临床试验,让23名健康志愿者每人每天连续30天摄取上述的HRB菌株。试验结果表明该类HRB菌株显著地增加了参试者粪便中的叶酸含量,从而表明该类HRB菌株能够在人的肠道中合成和分泌叶酸[29]。事实上,结肠是叶酸的主要贮藏库,而由双歧杆菌合成的叶酸还能起到维持粘膜肠细胞中维生素的动态平衡并预防结肠癌发生的作用[21]。因此,相比于非HRB,具有叶酸合成能力的HRB菌株在维持肠内维生素动态平衡以及改善宿主的健康应该更为优越。
2.2 碳水化合物代谢能力
双歧杆菌具有各种外切性糖苷酶和内切性糖苷酶等独特的代谢途径,以代谢利用肠道环境中的多种碳水化合物,而同时其碳水化合物代谢能力有助于双歧杆菌菌种在特定生态环境中定殖。值得注意的是,不同起源的双歧杆菌菌种之间在碳水化合物代谢能力方面有着显著的差异。其中,HRB菌株具有代谢各种多聚糖的重要基因簇和糖苷酶,可有效地代谢多种糖类碳水化合物如人乳低聚糖(human milk oligosaccharides,HMO s),黏液素(mucin)以及膳食来源的碳水化合物,从而反映出它们与人体宿主共同进化的关系。相比之下,非HRB菌株则缺乏可代谢如上所述的人类肠胃道中的各种碳水化合物的能力。
2.2.1 人乳低聚糖代谢能力
人类的母乳是婴儿的天然食物,它含有丰富的营养和能量,可促进新生儿的健康生长和发育[30]。人乳中含有乳糖、脂肪酸、低聚糖和分泌型IgA(sIgA),其中人乳低聚糖是母乳中的重要成分,它是一种低度聚合糖,不能被人的肠胃道消化吸收,但可以被一部分肠内细菌特别是双歧杆菌利用代谢产生短链脂肪酸等产物,起到①降低结肠pH值、促进钙、镁、铁等矿物质吸收;②给肠细胞提供营养、维持肠黏膜屏障和抵御病原微生物的侵袭等作用。因此,HMO s可促进肠内双歧杆菌,特别是婴儿型HRB菌株的增殖[31-32]。HMO s的基本单元包括半乳糖(Gal)、葡萄糖(Glu)、岩藻糖(Fuc)、唾液酸(Sia)和N-乙酰葡糖胺(Glu-NAc)。几乎所有的HMO s都包含一个乳糖核心基团(Gal-β-1,4-G lu)作为还原端,以通过β-1,3-或β-1,6-反复连接乳-N-生物糖(lacto-N-biose,LNB)或N-乙酰乳糖胺(N-acetyllactosam ine)而延伸[33]。目前研究发现,人乳中含有大约200多种HMO s,其中乳-N-四糖(lacto-N-tetraose,LN T)、乳糖-N-岩藻戊糖I(lacto-N-fucopentaose I,LN FP I)、乳糖-N-二岩藻己糖I(lacto-N-difucohexaose I,LNDFH I)和2’-岩藻糖(2’-fucosyllactose,2’-FL)的含量最为丰富(图4)。值得注意的是,这四种主要的HMO s中有三种是具有LN T结构的I型链寡糖[34],因此,LNT及其降解产物,LNB,是HMO s代谢中的关键结构。
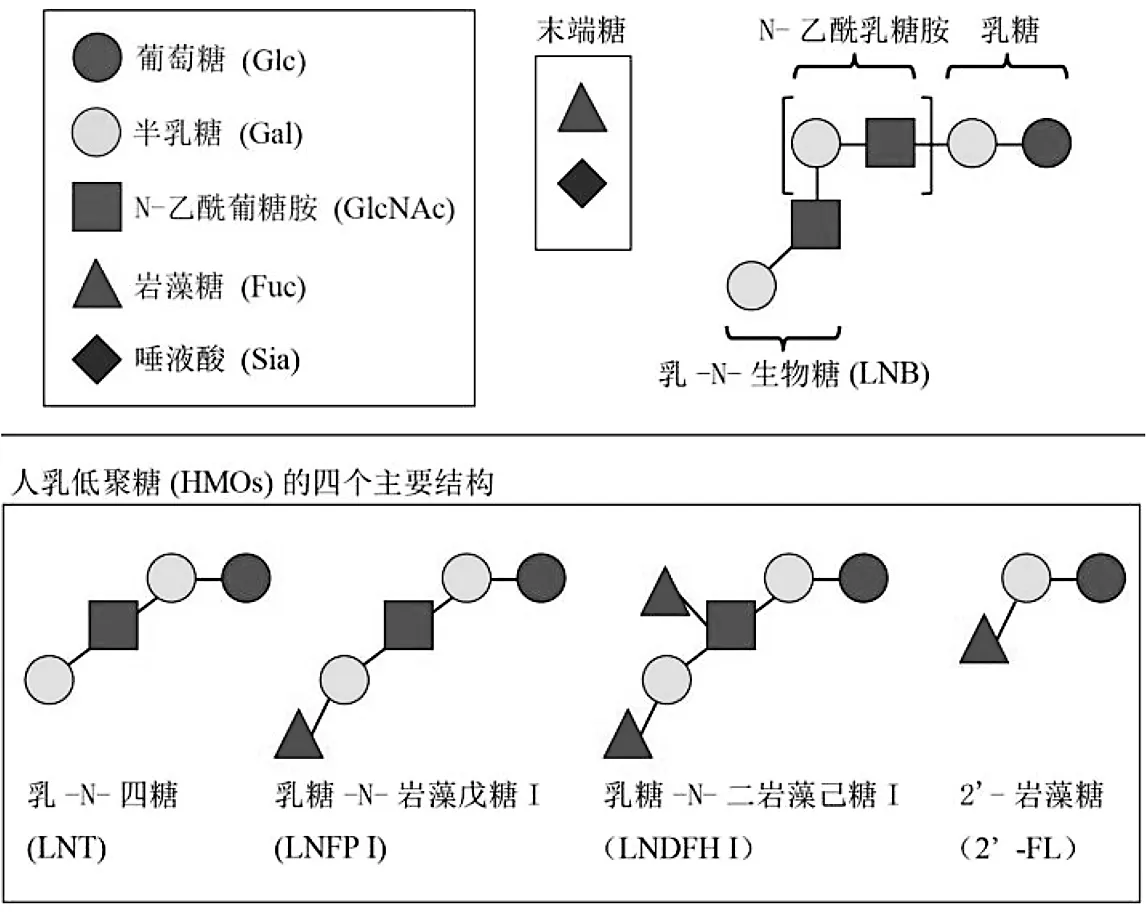
图4 人乳低聚糖(HMOs)的结构
图4中,HMO s的基本单元包括半乳糖(Gal)、葡萄糖(G lu)、岩澡糖(Fuc)、唾液酸(Sia)和N-乙酰葡糖胺(GluNAc)。HMO s包含一个乳糖核心基团(Gal-β-1,4-G lu)作为还原端,以通过β-1,3-或β-1,6-反复连接乳-N-生物糖(lacto-N-biose,LNB)或N-乙酰乳糖胺(N-acetyllactosam ine)而延伸。HMO s的四个主要结构包括乳-N-四糖(lacto-N-tetraose,LNT)、乳糖-N-岩藻戊糖I(lacto-N-fucopentaose I,LNFP I)、乳糖-N-二岩藻己糖 I(lacto-N-difucohexaose I,LNDFH I)和2’-岩藻糖(2’-fucosyllactose,2’-FL)。
HRB菌株和非HRB菌株之间的比较基因组学研究揭示了HRB菌株,特别是婴儿型HRB菌株在代谢利用LNT的能力[35]。其中,婴儿双歧杆菌的基因簇包含一系列降解HMO s所必要的糖苷水解酶(GH)和ABC转运器(ABC transporter)[36]。相比于其他菌种婴儿双歧杆菌可为利用HMO s的顶级好手,能降解不同类型的HMO s并能够利用HMO s作为唯一的碳源而增殖[37-38]。婴儿双歧杆菌具备细胞内降解各种完整的HMO s的途径,以将HMO s水解成单糖并用于生产能量[39](表1)。相对于婴儿双歧杆菌能够完整的利用各种HMO s分子,其他婴儿型HRB菌株如长双歧杆菌、两歧双歧杆菌和短双歧杆菌一般则拥有不完整的HMO s降解途径[40](表1)。两歧双歧杆菌与长双歧杆菌利用细胞外LNBase将大分子HMO s部分降解以后输入细胞内降解,而大部分短双歧杆菌菌株则需要依靠其他菌种在细胞外部分降解以后才能利用[34,37,41-42]。有趣的是,最近的一项研究中发现短双歧杆菌UCC 2003菌株也具有与婴儿双歧杆菌类似的LNT降解途径[43]。与之相反,成人型HRB菌株,如青春双歧杆菌和非HRB菌株,如动物双歧杆菌则不能降解LNT或其他可检测的HMO s[40](表1)。这些研究表明普遍定植于成年人和动物肠道以及其他来源的成人型HRB菌株和非HRB菌株可能不存在着或已经失去降解HMOs的能力。
Xiao等的研究显示,部分成人型HRB菌株,包括青春双歧杆菌、链状双歧杆菌、齿双歧杆菌和角双歧杆菌(B.angulatum)与非HRB菌株,包括乳双歧杆菌和嗜热双歧杆菌都不能利用LNB[44]。同时,Wada等也强调了双歧杆菌的LNBase活性功能的菌株特异性[41],其中一些婴儿型HRB菌株,如长双歧杆菌和两歧双歧杆菌拥有活性功能的LNBase以利用I型HMO s,反之,非HRB菌株,如动物双歧杆菌和假长双歧杆菌则缺乏LNBase[38,45]。总的来说,这些研究反映出婴儿型HRB菌株在适应婴儿肠道环境方面的共同进化,从而阐明了婴儿型HRB在代谢HMO s的优越性。

表1 亲和人体双歧杆菌(HRB)和非亲和人体双歧杆菌(非HRB)不同的人乳低聚糖(HMOs)降解途径
2.2.2 人类黏液素代谢能力
HRB和非HRB菌株具有各种不同遗传基因特征以适应并定植于人类肠胃道中。它们能够降解和吸收黏液素作为营养源,以便在肠道黏膜层内定植与存活[46]。其中,大部分的长双歧杆菌和两歧双歧杆菌的菌株均具备内切α-N-乙酰半乳糖胺酶(endo-α-N-acetylgalactosaminidase)的基因同源体以释放在黏液素寡糖核心结构中的Galβ1-3GalNAc[47]。肠道中含有极度有限可利用的碳源,因此具备该类基因的婴儿型HRB菌株具有可降解黏液素的能力,从而有利于它们在婴儿肠道中的适应和竞争。此外,O dam aki等发现大部分的HRB菌株,包括婴儿双歧杆菌、长双歧杆菌和短双歧杆菌具备α-N-乙酰半乳糖胺酶(α-N-acetylgalactosaminidase,N agBb)的基因同源物体以降解粘液素的核心糖链,然而,这些基因组并不存在于非HRB菌株[48]。
另外,有文献表明HRB菌株中的两歧双歧杆菌能够代谢黏液素[49-50]。此外,一项关于婴儿型HRB菌株降解黏液素能力的研究发现,两歧双歧杆菌D 119和L22、短双歧杆菌NC IM B8807和长双歧杆菌NCIM B8809在含有黏液素的培养基中生长良好,从而表明了该类婴儿型HRB菌株能够利用黏液素作为碳源以增殖[51]。相比之下,动物双歧杆菌和假链双歧杆菌的菌种无法生长在含有粘液素的培养基。因此,具有可代谢黏液素的能力的HRB菌株,特别是婴儿型HRB菌株,能够在人类肠胃道中存活并定植,从而阐明了HRB菌株的优越性。
2.2.3 植物来源碳水化合物代谢能力
除了宿主衍生的聚糖之外,双歧杆菌还能代谢多种不可被人类宿主消化的植物来源碳水化合物,特别是宿主膳食中的植物细胞壁碎片和淀粉颗粒。然而,HRB和非HRB菌种之间具有不同的植物来源碳水化合物的代谢能力。值得注意的是,HRB菌株含有编码代谢植物来源碳水化合物的各种基因组,它们可利用聚糖作为其主要的碳源来生存。其中,主要寄宿于婴儿和成人肠道中的长双歧杆菌富含糖基水解酶GH 43和51家族中的同源基因,可水解阿拉伯呋喃糖苷和木聚糖[48]。这些在长双歧杆菌中的基因组被视为有助于婴儿在断奶期间的消化,尤其是当摄取含有复合碳水化合物的非乳饮食的时候。此外,该类戊糖也存在于典型的植物性成人饮食中,从而解释了长双歧杆菌为何可生长和定殖于婴儿和成人肠道中的原因[17]。
此外,成人型HRB菌株中的青春双歧杆菌的基因组富含编码α-基葡糖苷酶,淀粉酶,支链淀粉酶和环麦芽糖糊精酶等活性酶的基因。青春双歧杆菌具有可广泛地代谢植物来源的碳水化合物的能力,其中包括在成人饮食中丰富含有的淀粉和淀粉样寡糖/多糖如支链淀粉(amylopectin,pullulan),麦芽三糖和麦芽糖糊精[52-53]。这些基因组结构反映出该类成人型HRB菌株在含有丰富的植物来源的聚糖的成年人肠胃道中的适应能力。
淀粉是膳食中的主要碳水化合物,可以被人类的消化酶消化吸收。有研究指出,尽管大部分动物来源的非HRB菌株具有广泛的淀粉降解的能力,而在HRB菌株中,只有青春双歧杆菌、角双歧杆菌和齿双歧杆菌能够降解和利用淀粉[54]。然而,一些非HRB菌株,如乳双歧杆菌、假长双歧杆菌和嗜热双歧杆菌等则缺乏编码代谢阿拉伯木聚糖寡糖的酶的基因[55]。这些研究结果表明了非HRB菌株代谢植物来源碳水化合物的遗传性基因组结构与其相应的宿主有着息息相关的关系,从而揭示了非HRB菌株与其宿主共同进化的过程[56]。有鉴于此,HRB菌株似乎具有优于非HRB菌株在人类宿主的适应性优势。显而易见,HRB菌株具备着可代谢多样化人类宿主不可消化的植物来源碳水化合物,从而为宿主提供能量并有助于宿主消化和营养摄入。
3 双歧杆菌与母乳的亲和性
研究证实母乳是影响婴儿肠道菌群的组成的关键因素,而尽管双歧杆菌可优先定殖于婴儿肠道,不同寄宿起源的双歧杆菌之间的母乳亲和性有着显着的差异[14,57]。M inam i等在一项评估37种双歧杆菌菌株在人类母乳中增殖能力的体外研究中表明婴儿型HRB菌株中的短双歧杆菌和婴儿双歧杆菌均在人类母乳中增殖良好,而长双歧杆菌和两歧双歧杆菌则显示出菌株间的差异但大部分菌株能够增殖[14](图5)。与之相反,大部分的成人型HRB菌株和非HRB菌株在人类母乳中不但不能增殖反而会失去活性[14](图5)。其他文献也阐明不同双歧杆菌菌种在人类母乳中增殖性的不同,其中可代谢人乳低聚糖的婴儿型HRB菌株如两歧双歧杆菌和短双歧杆菌的增殖优于成年型HRB菌株如青春双歧杆菌[57]。
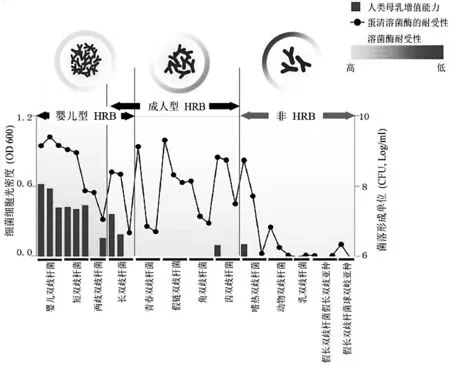
图5 亲和人体双歧杆菌(HRB)和非亲和人体双歧杆菌(非HRB)在人类母乳中的增殖能力与对溶菌酶的耐受性
图5中,红色柱子为双歧杆菌在人乳中的存活量,黑圈为双歧杆菌在含有蛋清溶菌酶的培养基中的存活量(OD值)。
应该注意的是,人乳中除了人乳低聚糖以外还含有大量的双歧杆菌可利用的乳糖(约70 m g/m L)[58],因此缺乏代谢人乳低聚糖的能力似乎不是决定成人型HRB菌株与非HRB菌株在母乳中失活的关键。研究显示母乳含有各种抗菌因子如免疫球蛋白、乳铁蛋白、溶菌酶、乳过氧化物酶、游离脂肪酸、抗菌肽等,可选择性地抑制有害菌的增殖[59]。其中,作为人类母乳中主要的抗菌因子之一的溶菌酶是影响肠道菌群定殖的最重要的因素之一。溶菌酶大量存在于人类母乳中(高达400μg/m L),但牛奶等动物乳汁中的含量极低,牛奶中的浓度为人母乳的千分之一左右[60]。它可选择性抑制细菌在人体生长,从而帮助建立适当的肠道菌群。
近期研究证实了溶菌酶可影响双歧杆菌菌种在母乳中的增殖,并且表明人乳溶菌酶可能是影响婴儿肠道中的双歧杆菌组成的关键因素[11,14,57]。M inam i等进行的体外研究显示,大部分婴儿型HRB菌株对高浓度的溶菌酶具有良好的耐受性,而成人型HRB菌株具有中等耐受性,反之,非HRB菌株则对溶菌酶具有较差的耐受性[14](图5)。在另一项类似的研究中,相比于非HRB菌株如乳双歧杆菌、假长双歧杆菌和瘤胃双歧杆菌,HRB菌株包括短双歧杆菌、两歧双歧杆菌和长双歧杆菌更能抵抗蛋清溶菌酶(400μg/m L)[61]。同时,耐溶菌酶的HRB菌株(两歧双歧杆菌)在含有15~58μg/m L人乳溶菌酶的母乳中增殖良好,而非HRB菌株(乳双歧杆菌)则失活[61]。
除了母乳之外,溶菌酶天然存在于几乎所有人体组织、体液和分泌物如泪液、鼻腔排出物、唾液、汗液和血清中[59]。因此,溶菌酶可能影响双歧杆菌在人体的生存能力,同时决定了具有溶菌酶耐受性的HRB菌株能够更好地定殖于人类肠道。此外,比较人类母乳或配方奶粉喂养婴儿的大便中的微生物构成的研究发现,母乳喂养婴儿的肠道中具有大量的HRB菌种如长双歧杆菌、短双歧杆菌和两歧双歧杆菌,而不存在非HRB菌种如动物双歧杆菌[62]。有鉴于人类母乳中含有人乳低聚糖和溶菌酶,而婴儿配方奶粉并没有,人乳低聚糖和溶菌酶可能是决定婴儿肠道中的双歧杆菌构成的主要因素(图6)[62]。

图6 人乳低聚糖和溶菌酶可能是影响亲和人体双歧杆菌(HRB)与非亲和人体双歧杆菌(非HRB)在婴儿肠道中组成的关键因素
图6中,婴儿型HRB菌株具有可降解人乳低聚糖的能力和较高的人乳溶菌酶的耐受性,可存活于母乳喂养的婴儿肠道中。成人型HRB菌株虽对溶菌酶有一定的耐受性但不能降解人乳低聚糖。非HRB菌株则欠缺降解人乳低聚糖的能力以及对溶菌酶的耐受性。
4 HRB对人体健康的优越性
随着益生菌领域的不断挖掘,目前双歧杆菌被广泛地应用于医疗、保健和食品等领域。在市面上我们可以看到的益生双歧杆菌包括HRB菌株,如长双歧杆菌,短双歧杆菌,婴儿双歧杆菌和两歧双歧杆菌以及非HRB菌株,如乳双歧杆菌[63-66]。尽管有人认为益生菌的功效归因于菌株水平的特异性的生理特征而不是其分离源,但如本文所表明的,我们也不能否认菌株的起源与其特性有很多关联,如双歧杆菌的生理特征中的叶酸合成、碳水化合物代谢能力和母乳亲和性等是取决于菌种菌株的寄宿起源。因此很有必要从临床角度评估HRB和非HRB菌株对人体中的功效与安全性。
然而,临床评估HRB和非HRB菌株对促进人类健康作用的研究并不多。有研究表明,给配方奶粉和人乳喂养的早产儿补充双歧杆菌时,HRB菌株如婴儿双歧杆菌的定殖优于非HRB菌株如乳双歧杆菌[67]。在人乳喂养的婴儿中,相比于乳双歧杆菌,服用婴儿双歧杆菌时会显著地增加婴儿大便中双歧杆菌的数量,并降低了有害的γ变形杆菌的数量[67]。这些研究结果表明,相比于非HRB菌株,HRB菌株能更有效地定殖于婴儿肠道中。另外一项涉及108位健康新生儿的研究发现,婴儿型HRB菌株,如短双歧杆菌或婴儿双歧杆菌为最优先定殖于婴儿肠道的菌种,而且其定殖能力不受分娩方式和喂养方法的影响,从而表明婴儿型HRB菌株在婴儿肠道中的优越性[68]。相比之下,非HRB菌株,如乳双歧杆菌的定殖能力则受喂养方法的影响而更容易定殖于配方奶粉喂养的婴儿肠道[68]。因此,正如其一般不存在于母乳喂养的婴儿肠道里一样,乳双歧杆菌可能不是常见的婴儿肠道菌群。
总的来说,HRB菌株将会是更理想的人类益生菌,特别是对于于婴幼儿,且对宿主会有更多的功效(表2)。毫无疑问,我们还需进一步地探讨HRB与非HRB的特征,特别是从临床角度研究HRB与非HRB菌株在人体中功能性的差异。

表2 亲和人体双歧杆菌(HRB)与非亲和人体双歧杆菌(非HRB)的不同生理特征
5 结束语
尽管双歧杆菌具有促进人类健康的功效,但并非所有菌株都具有相同的功能特征。本文显示了HRB与非HRB菌株之间的一些不同的生理特征。比较基因组学研究提供了HRB和非HRB菌株之间的功能差异的有力证据,并且显示了HRB和非HRB菌株在合成叶酸和代谢人乳低聚糖、粘液素和膳食聚糖的基因结构的不同。另外,针对双歧杆菌的生理特征所进行的大量体外研究以及临床评估指出,一些HRB菌株,特别是婴儿型HRB菌株比非HRB菌株更适合人类的肠道环境。综合HRB菌株可合成叶酸和代谢人类肠道中可用的各种聚糖的能力,以及它们在人类母乳中的亲和性,HRB菌株尤其是婴儿型HRB菌株可能更为适合用于婴幼儿。
猜你喜欢
右江医学(2022年10期)2022-11-03
中国乳品工业(2022年3期)2022-04-11
齐鲁工业大学学报(2021年1期)2021-12-30
世界最新医学信息文摘(2021年77期)2021-01-07
中国生殖健康(2019年11期)2019-01-07
消费导刊(2018年8期)2018-05-25
环球时报(2017-12-20)2017-12-20
中国药业(2014年19期)2014-05-17
食品工业科技(2014年13期)2014-03-11
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
