
胃癌相关miRNA-21靶位点多态性分析
2018-09-07 10:28王任翔
西安交通大学学报(医学版) 2018年5期
王任翔,贾 皑
(1. 长安大学医院内科,陕西西安 710064;2. 西安交通大学第一附属医院消化内科,陕西西安 710061)
胃癌(gastric cancer, GC)是全球第四大最常见的癌症。尽管随着现代医疗的发展,胃癌发病率逐渐下降[1-2],但是,每年仍有100万新发病例。胃癌的高发病区包括东亚、东欧和南美洲,胃癌的5年生存率约20%~25%[3]。胃癌的发生是一个多因素多步骤的过程,涉及到许多环境、遗传、表观遗传和其他因素[4]。研究表明低社会经济地位、幽门螺杆菌感染、吸烟、饮酒和某些人口特征与胃癌高风险有关[5]。也有多项研究报道有关遗传因素在胃癌分子发病机制中的作用[6-7]。
微小RNA(micro RNA, miRNA,miR)是18~23个核苷酸长度的分子,通过与其靶mRNA的3′非翻译区结合,可以抑制原癌基因和抑癌基因的翻译,从而调控信号转导和细胞周期过程[8]。越来越多的研究证明miRNA参与人类肿瘤的发生发展过程。研究发现胃癌患者通常存在miR-21异常表达[9-10]。miR-21调控一些靶点而影响肿瘤发生。但是其确切作用机制仍知之甚少,目前尚无直接证据证明miR-21与胃癌发生之间存在相关性。本研究检测胃癌患者miR-21的靶位点rs300574、rs4272、rs4728、rs11911、rs3744935、rs8708和rs7828的多态性,分析其与胃癌患病风险的相关性。
1 资料与方法
1.1临床资料选择2015年1月至2016年12月西安交通大学第一附属医院消化内科收治的胃癌患者300例,入选条件:原发性胃癌新发病例,所有病例均符合中国《胃癌诊断标准》[11],未经过手术、放射治疗、化学治疗等方法治疗;精神状况良好;研究对象之间无直接亲属关系。选择同时期西安交通大学第一附属医院体检中心健康人群300例为对照组,入选条件:既往无肿瘤病史、慢性疾病、严重代谢性疾病及内分泌失调等;精神状况良好;甲胎蛋白和癌胚抗原阴性;研究对象之间无直接亲属关系。本研究已获得所有研究对象的知情同意,符合伦理学的规定,并通过医院伦理委员会批准。
1.2试剂与仪器全血基因组纯化试剂盒(美国Thermo Fisher公司),iPLEX Gold 试剂盒(美国Sequenom公司);MassARRAY质谱仪(美国Sequenom公司),GeneAmp 9700 PCR仪(美国ABI公司)。
1.3血样采集及DNA提取采集每位研究对象外周静脉血5 mL,置于EDTA-K2抗凝管中,现场编号并登记姓名,所有标本采集当天送实验室,置于-70 ℃冰箱保存,以备DNA提取。采用全血DNA纯化试剂盒进行DNA提取,操作步骤严格按照说明书进行。
1.4基因分型在Target Scan Human数据库(http://www.targetscan.org/)中寻找miR-21相关的靶基因及其结合位点,并选择中国人群具有较高频率(最小等位基因频率≥5%)位点,最终确定了rs300574、rs4272、rs4728、rs11911、rs3744935、rs8708和rs7828共7个位点。采用Sequenom MassARRAY SNP基因分型技术对所选择的SNP位点进行分型。单碱基延伸引物委托上海生工生物工程技术服务公司进行设计和合成。在MassARRAY质谱仪中进行反应,基因分型使用Sequenom MassARRAY RS1000软件进行,引物信息见表1。
表1本研究所用到的引物序列
Tab.1 Primers used for this study

SNP_ID1st-PCRP2nd-PCRPUEP_SEQrs300574ACGTTGGATGGGGTCAGGGTAAACCATCATACGTTGGATGCAAAAAACAGCCACAACTTGAAAAAACAGCCACAACTTGAAAGCTArs4272ACGTTGGATGGATAAAACACCTAGATACCCACGTTGGATGCTAAGCCCCCAAATAAGCTGAAATAAGCTGCATGCATTTGTAACArs4728ACGTTGGATGCACATATGCAACCAACTAAACACGTTGGATGAGCATTTCACCGCAAACAGCCAAACAGCAAAGACTATCCCArs11911ACGTTGGATGGTTGGTTGCATATGTGTATACGTTGGATGCCCTAGGAACTTTGAAAAACCACAGTAATGAGGATTTTTTCTrs3744935ACGTTGGATGCCTCTACAGTGATACATGTCACGTTGGATGTGTGGCTGGGCCTGTCACCAGCCCTCCAGGTAGGCCCrs8708ACGTTGGATGGAGCTTACGTAGTTCTACCGACGTTGGATGTACCCCGATTACCAGAACAGAGGGTGGCAGAAGCACArs7828ACGTTGGATGGTGGTTCAGTATTATGTACGACGTTGGATGAATTGTGTGAACTTGGAAGCTTGGAAGCACACCAATCT
1.5统计学分析应用SPSS 19.0软件进行统计分析。比较两组间的基本信息,对于连续性变量,采用t检验;对于分类变量,采用χ2检验。采用直接计数法统计各位点的等位基因及基因型频数,采用χ2检验比较两组间各位点的等位基因频率的分布差异。应用SNPStats在线软件(http://bioinfo.iconcologia.
net/snpstats/start.htm)评估各SNP位点基因型与胃癌患病风险[12]。以相对优势比(odds ratio,OR)和95%可信区间(confidence intervals,CI)表示SNP与胃癌患病风险的关系。P<0.05(双侧)为差异有统计学意义。用Haploview 4.2软件进行单体型与胃癌患病风险的相关性分析。
2 结 果
2.1病例组及对照组的基本信息共收集300例胃癌患者和300例健康对照,其中胃癌组男228 例,女72例,平均年龄(56.18±13.44)岁,对照组男237例,女63例,平均年龄 (55.34±14.12)岁,胃癌病例与对照组之间在年龄、性别等人口学特征上差异无统计学意义(P>0.05),说明两组均衡可比。
2.2候选SNP基本信息miR-21的7个靶SNP位点的基本信息见表2,基因、染色体位置、最小/野生等位基因和最小等位基因频率,7个位点均符合Hardy-Weinberg平衡定律(HWEP>0.05)。采用χ2检验比较两组间各位点的等位基因频率差异,结果显示SPRY1基因上rs300574位点的等位基因“C”和Bcl-2基因上rs3744935位点的等位基因“T”与增加胃癌的患病风险具有相关性(P<0.05),而JAG1基因上rs8708位点的等位基因“C”与降低胃癌的患病风险具有相关性(P<0.05)。
表2miR-21靶位点的基本信息及其在病例和对照组中的最小等位基因频率
Tab.2 Basic information about miR-21 target sites and minor allele frequency in the patients and the controls

SNPmiRNA基因 染色体位置等位基因最小等位基因频率病例组(n=300)对照组(n=300)HWEPOR(95% CI)Prs300574miR-21SPRY14q28.1C/T0.4360.2920.4731.875(1.462~2.404)0.001rs4272miR-21CDK67q21.2G/A0.1190.1070.1051.135(0.778~1.668)0.526rs4728miR-21SPRY213q31.1C/A0.2480.2621.0000.942(0.718~1.294)0.687rs11911miR-21SPRY213q31.1C/A0.4670.4150.9051.239(0.976~1.581)0.082rs3744935miR-21Bcl-218q21.33T/C0.2320.1480.4771.735(1.277~2.358)0.001rs8708miR-21JAG120p12.2C/T0.2010.3350.5970.503(0.382~0.761)0.001rs7828miR-21JAG120p12.2C/A0.1640.1720.2120.948(0.688~1.221)0.776
2.3候选SNP在遗传模型下与胃癌风险的相关性引入遗传模型,评估风险等位基因在4个不同遗传模型(共显性、显性、隐性和加性)下的风险,按照性别、年龄修正,结果见表3。在最佳模型-显性模型下,rs300574位点的T/C和C/C基因型与增加胃癌患病风险相关(OR=2.64,95%CI:1.73~4.02,P<0.000 1);rs3744935位点的C/T和T/T基因型也与增加胃癌患病风险相关(OR=1.58,95%CI: 1.04~2.41,P=0.033);而rs8708位点的C/T和C/C基因型与降低胃癌患病风险具有相关性(OR=0.42,95%CI:0.28~0.63,P<0.000 1)。
2.4候选SNP单体型与胃癌风险的相关性对所选择的7个SNP位点进行进一步的连锁不平衡分析,并构建单体型。结果显示位于JAG1基因上的rs8708和rs7828两个位点具有强连锁关系(图1)。经过分析单体型与胃癌风险相关性,结果显示CA单体型与降低胃癌患病风险具有相关性(OR=0.18,95%CI:0.13~0.44,P<0.000 1,表4)。
3 讨 论
本研究共选取7个位于不同基因上的miR-21靶SNP位点并检测其多态性,发现SPRY1基因上rs300574位点的等位基因“C”、Bcl-2基因上rs3744935位点的等位基因“T”和JAG1基因上rs8708位点的等位基因“C”与胃癌患病风险具有相关性。该结果首次揭示了miR-21靶SNP位点与胃癌患病风险之间的相关性,为后续深入研究miR-21与胃癌发生的具体机制提供了新线索。
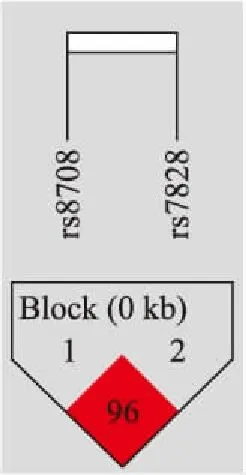
图1JAG1基因上4个候选SNP的连锁不平衡图
Fig.1 Linkage disequilibrium of four candidates SNPs inJAG1 gene
miRNA是一类新的非编码RNA,通过抑制翻译或切割/衰减mRNA而调控靶基因的表达[13]。全基因组研究显示,miRNA基因通常位于癌症相关基因组区域或脆性位点,以及杂合性缺失或扩增的最小区域,或共同的断点区域,表明miRNA在肿瘤发生中的潜在作用。在不同类型的肿瘤如乳腺癌、白血病以及肺癌和结肠癌中,也通常存在miRNA的异常表达[14],这表明miRNA与人类恶性肿瘤密切相关。CHAN等[15]报道miR-21在人类胶质母细胞瘤中的表达明显升高,并且通过阻断关键凋亡相关基因的表达而促进了恶性表型的形成。SI等[16]发现miR-21在乳腺肿瘤中高度表达,并且antimiR-21介导的细胞生长抑制与细胞凋亡增加和增殖减少具有相关性。也有报道miR-21在恶性胆管细胞中过度表达;抑制miR-21可增加个体对吉西他滨的敏感性[17]。另外,在大规模miRNA芯片检测中,VOLINIA等[18]观察到胃癌等多种实体肿瘤中miR-21的过度表达。但是,miR-21在胃癌发生中的作用和相关途径却不清楚。本研究通过对miR-21靶位点的不同基因型的分析检测,表明miR-21的靶位点rs300574、rs3744935和rs8708的多态性可能与胃癌的发生发展之间存在相关性,这也提示SPRY1、Bcl-2和JAG1的基因多态性可能与胃癌风险之间存在相关性。
表3候选SNP在遗传模型下与胃癌风险的相关性
Tab.3 Association between candidate SNPs and the risk of GC in genetic model
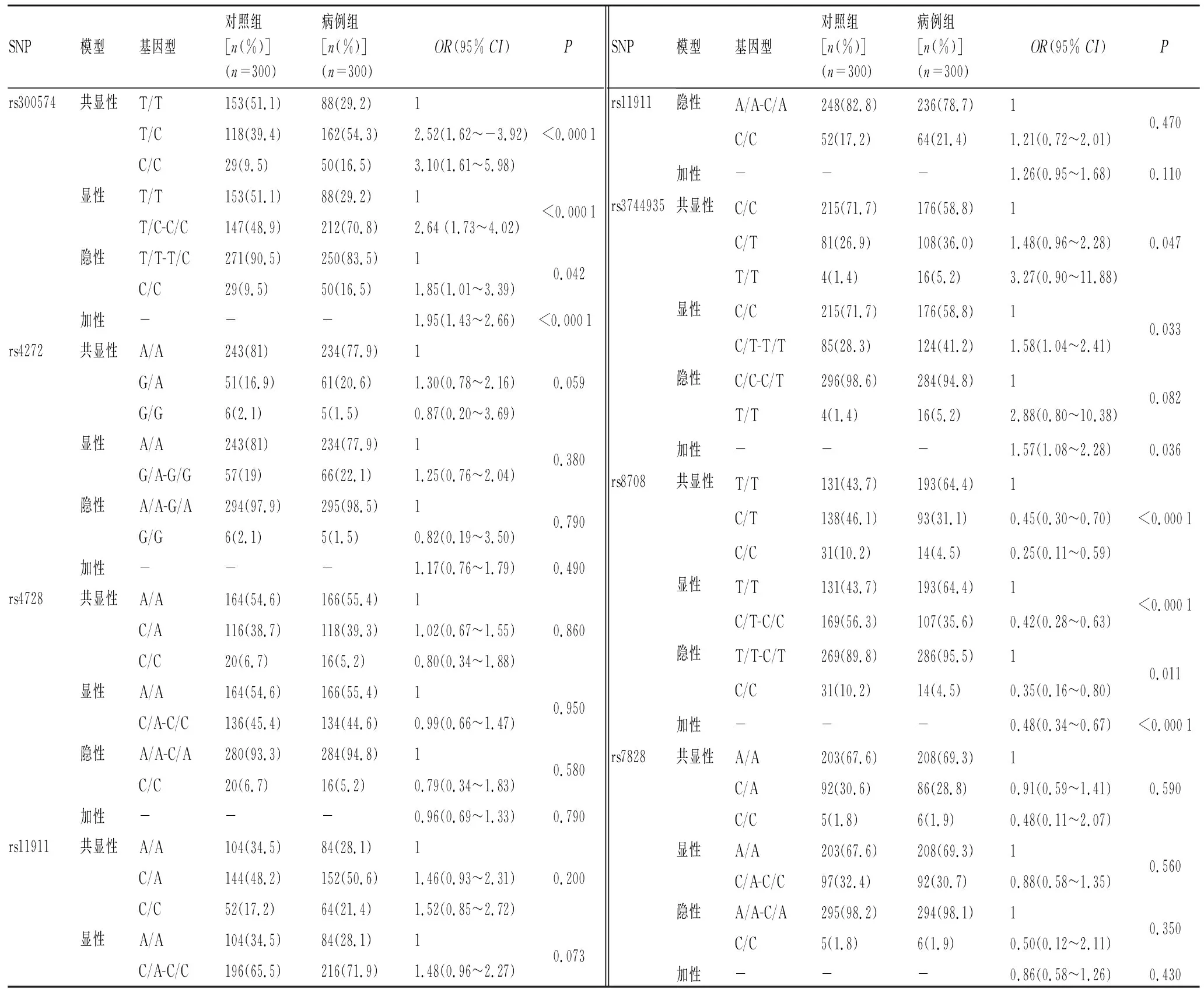
SNP模型基因型对照组[n(%)](n=300)病例组[n(%)](n=300)OR(95% CI) Prs300574共显性T/T153(51.1)88(29.2)1T/C118(39.4)162(54.3)2.52(1.62~-3.92)C/C29(9.5)50(16.5)3.10(1.61~5.98)<0.0001显性T/T153(51.1)88(29.2)1T/C-C/C147(48.9)212(70.8)2.64 (1.73~4.02)<0.0001隐性T/T-T/C271(90.5)250(83.5)1C/C29(9.5)50(16.5)1.85(1.01~3.39)0.042加性---1.95(1.43~2.66)<0.0001rs4272共显性A/A243(81)234(77.9)1G/A51(16.9)61(20.6)1.30(0.78~2.16)G/G6(2.1)5(1.5)0.87(0.20~3.69)0.059显性A/A243(81)234(77.9)1G/A-G/G57(19)66(22.1)1.25(0.76~2.04)0.380隐性A/A-G/A294(97.9)295(98.5)1G/G6(2.1)5(1.5)0.82(0.19~3.50)0.790加性---1.17(0.76~1.79)0.490rs4728共显性A/A164(54.6)166(55.4)1C/A116(38.7)118(39.3)1.02(0.67~1.55)C/C20(6.7)16(5.2)0.80(0.34~1.88)0.860显性A/A164(54.6)166(55.4)1C/A-C/C136(45.4)134(44.6)0.99(0.66~1.47)0.950隐性A/A-C/A280(93.3)284(94.8)1C/C20(6.7)16(5.2)0.79(0.34~1.83)0.580加性---0.96(0.69~1.33)0.790rs11911共显性A/A104(34.5)84(28.1)1C/A144(48.2)152(50.6)1.46(0.93~2.31)C/C52(17.2)64(21.4)1.52(0.85~2.72)0.200显性A/A104(34.5)84(28.1)1C/A-C/C196(65.5)216(71.9)1.48(0.96~2.27)0.073SNP模型基因型对照组[n(%)](n=300)病例组[n(%)](n=300)OR(95% CI) Prs11911隐性A/A-C/A248(82.8)236(78.7)1C/C52(17.2)64(21.4)1.21(0.72~2.01)0.470加性---1.26(0.95~1.68)0.110rs3744935共显性C/C215(71.7)176(58.8)1C/T81(26.9)108(36.0)1.48(0.96~2.28)T/T4(1.4)16(5.2)3.27(0.90~11.88)0.047显性C/C215(71.7)176(58.8)1C/T-T/T85(28.3)124(41.2)1.58(1.04~2.41)0.033隐性C/C-C/T296(98.6)284(94.8)1T/T4(1.4)16(5.2)2.88(0.80~10.38)0.082加性---1.57(1.08~2.28)0.036rs8708共显性T/T131(43.7)193(64.4)1C/T138(46.1)93(31.1)0.45(0.30~0.70)C/C31(10.2)14(4.5)0.25(0.11~0.59)<0.0001显性T/T131(43.7)193(64.4)1C/T-C/C169(56.3)107(35.6)0.42(0.28~0.63)<0.0001隐性T/T-C/T269(89.8)286(95.5)1C/C31(10.2)14(4.5)0.35(0.16~0.80)0.011加性---0.48(0.34~0.67)<0.0001rs7828共显性A/A203(67.6)208(69.3)1C/A92(30.6)86(28.8)0.91(0.59~1.41)C/C5(1.8)6(1.9)0.48(0.11~2.07)0.590显性A/A203(67.6)208(69.3)1C/A-C/C97(32.4)92(30.7)0.88(0.58~1.35)0.560隐性A/A-C/A295(98.2)294(98.1)1C/C5(1.8)6(1.9)0.50(0.12~2.11)0.350加性---0.86(0.58~1.26)0.430
表4JAG1基因候选SNP的单体型与胃癌风险的相关性
Tab.4 Association between haplotype of SNP inJAG1 and the risk of GC
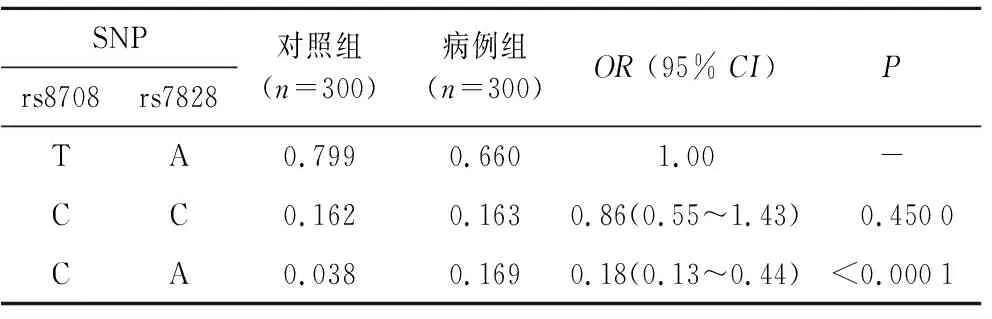
SNPrs8708rs7828对照组(n=300)病例组(n=300)OR (95% CI)PTA0.7990.6601.00-CC0.1620.1630.86(0.55~1.43) 0.4500CA0.0380.1690.18(0.13~0.44)<0.0001
SPRY1基因位于人类染色体4q28.1区域,是RAS信号通路上游的一个拮抗物。RAS基因是在各种癌症中突变频率最高的基因,其突变引起SPRY1基因过表达,而SCHAAF等[19]发现在RAS基因突变时,作为RAS信号通路的拮抗剂的SPRY1却转为激活剂作用。SPRY1基因多态性与肥胖和骨质疏松具有相关性,而有关SPRY1基因多态性与胃癌遗传易感性的文献信息较少,本研究首次发现SPRY1基因上rs300574位点的等位基因“C”与增加胃癌风险相关,后续应进一步用功能实验验证其在相关通路中的机制及作用。
Bcl-2基因位于人类染色体18q21.33区域,所编码的外线粒体膜蛋白具有抑制细胞凋亡的作用。PARKER等[20]研究报道Bcl-2相关蛋白引发的过度凋亡和促凋亡作用与低风险的骨髓增生异常综合症具有相关性。而一项国际合作项目表明MYC /Bcl-2蛋白共表达导致弥漫大B细胞淋巴瘤患者的低生存率和高患病风险[21]。KONTUREK等[22]在胃癌中观察到Bax/Bcl-2系统失调,并且Bcl-2显著上调。本研究首次发现Bcl-2基因上rs3744935位点的等位基因“T”与增加胃癌风险相关,进一步说明Bcl-2基因在胃癌发生发展中可能有重要作用。
JAG1基因位于人类染色体20p12.2区域,由JAG1编码的jagged 1蛋白是果蝇jagged蛋白的人类同源物。已有报道JAG1的突变会引起Alagille综合症。JAG1是Notch的配体,而Notch信号通路在胃癌的发生发展中起重要作用[23]。而本研究发现JAG1基因上rs8708位点的等位基因“C”与降低胃癌的患病风险具有相关性,这与之前研究结果一致,然而其涉及到的具体作用机制还有待进一步探索。
综上所述,本研究结果表明miR-21靶位点rs300574、rs3744935和rs8708的多态性可能与胃癌的发生发展之间存在相关性。这为临床上胃癌的早期诊断与预防提供了新的候选基因位点,也为后续深入研究miR-21参与胃癌发病机制提供线索。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
川北医学院学报(2022年6期)2022-06-24
昆明医科大学学报(2022年2期)2022-03-29
智慧健康(2021年17期)2021-07-30
聊城大学学报(自然科学版)(2020年1期)2020-11-27
科学导报(2020年51期)2020-09-09
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
中国神经精神疾病杂志(2013年4期)2013-03-11
