
喀斯特高寒干旱区不同经济树种的碳氮磷钾生态化学计量特征
2018-09-19 08:28喻阳华邢容容秦仕忆
生态学报 2018年15期
王 璐,喻阳华,邢容容,秦仕忆
1 贵州师范大学地理与环境科学学院,贵阳 550025 2 贵州师范大学喀斯特研究院/国家喀斯特石漠化防治工程技术研究中心,贵阳 550001 3 贵州大学资源与环境工程学院,贵阳 550025
生态化学计量学是一门结合了生物学、化学和物理学等基本原理,研究生物系统能量平衡和多重化学元素平衡的科学[1- 3],为研究生态系统中元素的耦合关系提供一种综合方法[4]。生态化学计量学目前已有两个基本理论,即“生长速率理论”与“内稳态理论”。“生长速率理论”指与RNA分配、基本化学元素计量比以及有机体生活史相关的机制[5]。“内稳态理论”是指有机体赖以生存的环境资源供应量与其化学元素组成保持的一种稳定状态[6]。碳(C)是构成植物体干物质的主要元素[7],氮(N)和磷(P)是生物体蛋白质和遗传物质的基本组成元素[8]。植物通过光合作用固定C,同时将部分C转移到土壤,并以凋落物的形式将C和养分逐步补偿给土壤[9]。生态系统内部的C、N、P循环在植物叶片、凋落物和土壤之间相互转换[10],使得研究“叶片-凋落物-土壤”连续体的养分含量特征复杂性增加。目前,众多学者已对不同区域、不同尺度的生态系统开展了广泛的生态化学计量特征研究。关于喀斯特地区生态系统生态化学计量方面的研究主要集中在单一组分或两组分之间。例如,胡忠良[11]对贵州喀斯特山区不同植被下土壤进行研究,提出植被类型的变化对土壤有效态养分的影响较全量养分显著,在生态系统养分循环研究中应更关注有效养分的变化。潘复静等[12]对典型喀斯特峰丛洼地植被群落凋落物进行研究,指出凋落物在P素较低的情况下具有较高的N及木质素含量,分解速率较低。罗绪强等[13]对茂兰喀斯特森林常见钙生植物叶片进行研究,发现喀斯特钙生植物具有低P、钾(K)和高Ca、Mg的特点,绝大部分植物属于P制约型植物。俞月凤等[14]对喀斯特峰丛洼地不同森林类型植物和土壤进行研究,指出喀斯特峰丛洼地土壤总体养分含量较高,其温湿条件极有利于生物繁衍和生长,生物“自肥”作用强烈,同时加速了岩石溶蚀、风化以及土壤形成和发育进程。曾昭霞等[15]对桂西北喀斯特区原生林与次生林鲜叶和凋落叶进行研究,认为原生林凋落物分解相对较慢,原生林能相对多的保留养分以供植物吸收,更能适应喀斯特石生环境。但是对于喀斯特高寒干旱区经济树种“叶片-凋落物-土壤”连续体C、N、P、K养分含量及生态化学计量特征的相关性、差异性研究鲜有报道,对于不同组分之间相互作用的养分调控因素认识不够深入。这阻碍了对化学计量“内稳态理论”、“生长速率理论”以及生态系统养分限制状况的进一步理解。因此,加强“叶片-凋落物-土壤”连续体生态化学计量的研究,具有十分重要的理论和现实意义。
毕节市七星关区撒拉溪示范区,具有典型的喀斯特地貌,生态系统退化,水土流失以及石漠化现象严重,提高养分再吸收率是该区植物适应贫乏生境的重要竞争机制。栽种经济树种是防治喀斯特石漠化的重要措施,能使经济效益与生态效益统一,实现林分的可持续经营。喀斯特地区植被恢复不应仅关注成熟林和自然林,经济林幼龄阶段也应得到重视。本文选取毕节市七星关区撒拉溪示范区的刺梨(Rosaroxburghii)、核桃(Juglansregia)2种主要经济树种作为研究对象,测定其叶片、凋落物以及根区土壤的C、N、P、K含量,探究研究区主要经济树种的养分含量及生态化学计量特征,能够揭示经济林幼龄阶段的养分需求规律,有助于深入地认识生态系统养分循环特征和系统稳定机制,以期为喀斯特地区植被恢复,养分限制状况提供一定的理论依据。
1 研究区概况
位于贵州省毕节市撒拉溪示范区,属毕节市西部六冲河流域支流区,地理坐标为105°02′01″—105°08′09″E,27°11′36″—27°16′51″N。示范区总面积8627.19 hm2,喀斯特面积占总面积的73.94%,海拔1600—2000 m,多年平均降雨量在984.40 mm,年平均气温约为12℃,≥10℃年积温3717℃,气候冬春温凉干旱,夏秋温润湿热,属喀斯特高原山地生态环境;地貌类型多样,地形破碎,耕地多分布于坡面上、台地和山间谷地,常形成环山梯土和沟谷坝地;有明暗交替的河流,漏斗、盲谷、落水洞、天窗、溶蚀洼地;地带性植被主要为针阔混交林,但是长期以来由于人类干扰过度,旱地植被占有较大优势;土壤以黄壤为主,部分地区有山地黄棕壤和石灰土[16]。研究区在种植刺梨和核桃前,曾连续多年种植玉米。裸地0—30 cm土层有机C为16.95 mg/g,全N为1.48 mg/g,碱解N为0.20 mg/g,全P为0.48 mg/g,速效P为0.008 mg/g,全K为4.62 mg/g,速效K为0.047 mg/g[17]。
2 研究方法
2.1 样品采集与处理
分别在树龄均为3年的刺梨林和核桃林中设置10 m×10 m典型样地,于两个样地中随机、均匀地选取5株健康且长势相近的树种(表1),分东、南、西、北4个方位取其冠下部(距地面2—3 m)的成熟叶片,分别混匀后装进信封。在样地中,按“梅花”五点法布设凋落物收集框,每个样地布设五个收集框,将凋落物混合收集,装入尼龙袋,带回实验室。所有植物叶片和凋落物样品放在烘箱中75℃烘至恒重,经过研磨,装入自封袋以备养分分析。

表1 采样树种基本概况
在采集植物叶片的同一样方,按照S形布点法对刺梨、核桃两类树种的根区土壤进行取样,5点取样组成混合样约1 kg。剔除凋落物、石砾及动物残体后,装入自封袋带回实验室。样品于室内自然风干、研磨、过筛保存备用。
2.2 样品分析
叶片、凋落物和根区土壤有机C采用重铬酸钾外加热法测定,全N采用高氯酸-硫酸消煮后用半微量凯氏定氮法测定,全P采用高氯酸-硫酸消煮-钼锑抗比色-紫外分光光度法测定,全K采用氢氟酸-硝酸-高氯酸消解-火焰光度计法测定。根区土壤碱解N采用碱解扩散法测定,根区土壤速效P采用氟化铵-盐酸浸提-钼锑抗比色-紫外分光光度法测定,根区土壤速效K采用中性乙酸铵溶液浸提-火焰光度计法测定。
2.3 数据处理
采用Excel 2010对本文数据进行初步分析与整理,计算平均值及标准差等。使用SPSS 20.0统计软件对数据进行分析,采用单因素方差分析(one-way ANOVA)对叶片、凋落物、根区土壤养分含量以及化学计量比进行差异性检验,并使用最小显著差数(LSD)法进行多重比较,采用Pearson相关分析法分析各组分生态化学计量特征关系。利用Origin8.6制图。N(P)再吸收率计算公式[18]为:

3 结果与分析
3.1 叶片-凋落物-土壤养分含量特征
从表2可以看出,2种经济树种的有机C、全N含量均表现为叶片>凋落物>根区土壤,核桃的全P含量为叶片>凋落物>根区土壤,刺梨不同组分中全P含量变化不显著,刺梨、核桃的全K含量均呈现出根区土壤最多,叶片次之,凋落物最少。比较2种树种各组分的养分含量可知,核桃叶片中有机C含量最多(均值为436.73 mg/g),刺梨叶片中全N、全P含量最多(均值为20.77、2.10 mg/g),核桃根区土壤中全K含量最丰富(均值为17.07 mg/g)。除刺梨不同组分的全P含量差异不显著外,刺梨不同组分的有机C、全N、全K含量以及核桃不同组分的全P、全K含量差异均达显著水平(P<0.05,下同)。
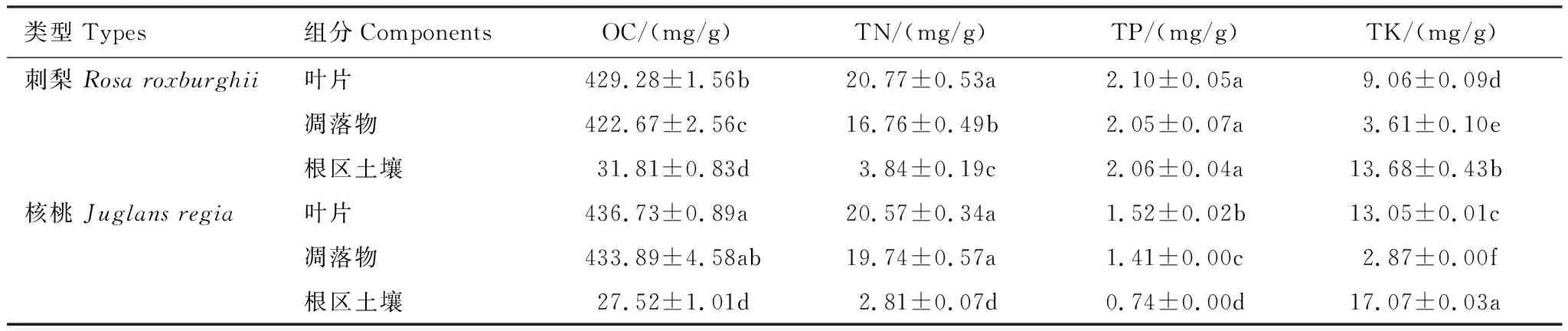
表2 不同经济树种叶片、凋落物和根区土壤的有机C、全N、全P、全K含量(平均值±标准差)
同一列数值后的不同小写字母代表同一测定指标不同组分间差异显著(P<0.05);OC:有机碳,organic carbon;TN:全氮,total nitrogen;TP:全磷,total phosphorus;TK:全钾,total potassium
刺梨根区土壤的碱解N,速效P、速效K含量均值为0.16、0.03、0.09 mg/g,核桃根区土壤的碱解N,速效P、速效K含量均值为0.18、0.01、0.66 mg/g,核桃根区土壤中碱解N,速效K含量均高于刺梨,而核桃速效P含量则低于刺梨,2种树种根区土壤的碱解N,速效P、速效K含量均差异显著(图1)。
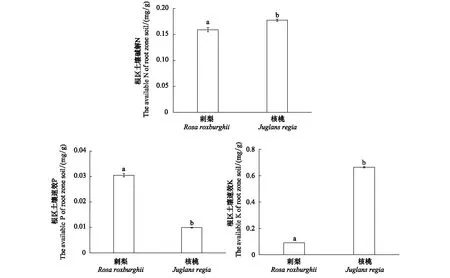
图1 不同树种根区土的碱解N、速效P、速效K含量(平均值±标准差)Fig.1 The available N, available P, available K content of root zone soil in different tree species (mean ± SD)不同小写字母表示同一养分含量不同树种间差异显著(P<0.05)
3.2 叶片N、P养分再吸收率特征
由图2可知,刺梨N、P再吸收率分别为19.23%、2.39%,核桃N、P再吸收率均值为4.05%、7.46%。刺梨N再吸收率显著高于核桃N再吸收率,P再吸收率表现为核桃>刺梨。刺梨P再吸收率低于N再吸收率,而核桃P再吸收率略高于N再吸收率。
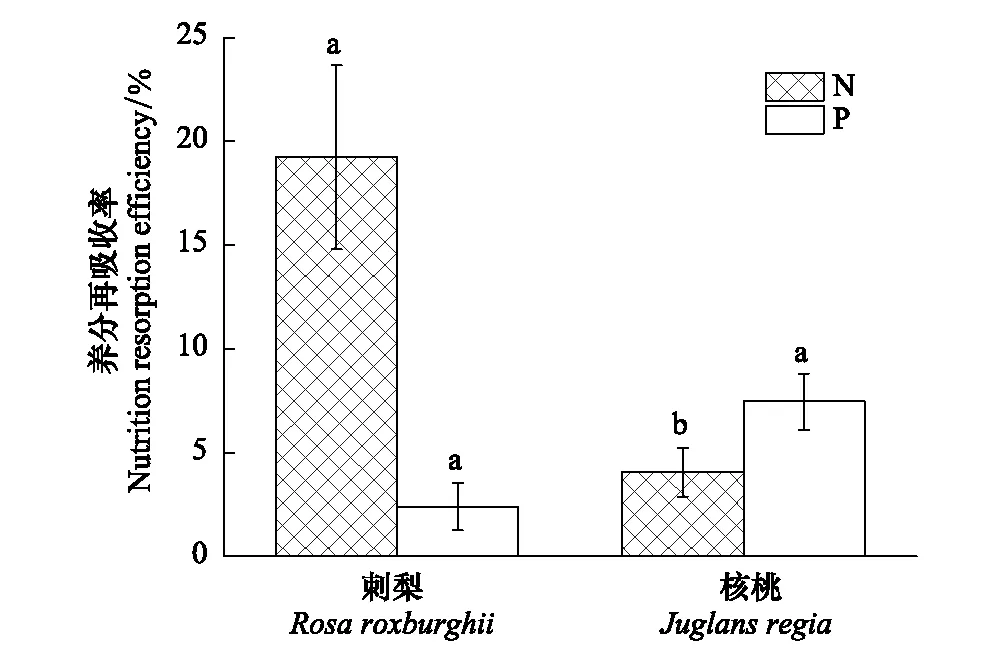
图2 不同树种植物N、P养分再吸收率(平均值±标准差) Fig.2 The N and P resorption rates of plants in different tree species (mean ± SD)不同小写字母表示同一养分再吸收率不同树种间差异显著(P<0.05)
3.3 叶片-凋落物-土壤生态化学计量特征
如图3所示,刺梨叶片、凋落物、根区土壤的C∶N均值分别为20.68、25.22、8.31;C∶P均值依次是204.30、206.14、15.45;C∶K均值分别是47.37、117.27、2.33;N∶P均值依次为9.89、8.18、1.86;N∶K均值分别为2.29、4.65、0.28;P∶K均值依次为0.23、0.57、0.15。核桃叶片、凋落物、根区土壤的C∶N均值依次为21.23、21.99、9.79;C∶P均值分别为286.66、307.77、37.10;C∶K均值分别是33.47、151.15、1.61;N∶P均值分别为13.51、14.00、3.79;N∶K均值分别为1.58、6.88、0.16;P∶K均值依次为0.12、0.49、0.04。刺梨不同组分C∶P、N∶P均低于核桃,P∶K则均高于核桃。2种经济树种的生态化学计量特征总体呈现出凋落物>叶片>根区土壤的规律。刺梨、核桃不同组分C∶K、N∶K、P∶K,刺梨不同组分C∶N、N∶P以及核桃不同组分C∶P均达显著性差异。
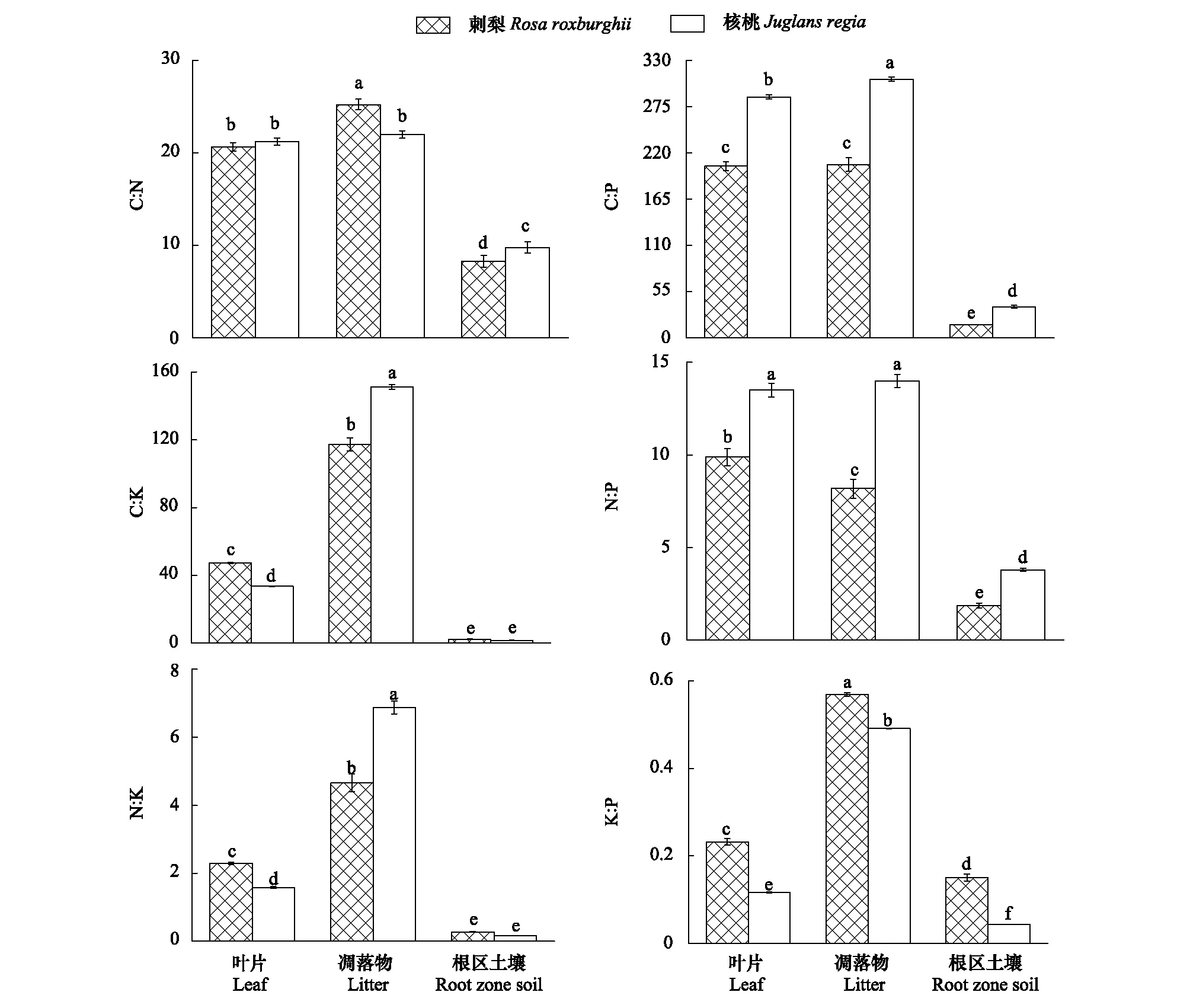
图3 不同树种叶片、凋落物、根区土壤的生态化学计量特征(平均值±标准差)Fig.3 The ecological stoichiometry characteristics of leaf, litter and root zone soil in different tree species (mean ± SD)不同小写字母代表同一树种不同组分之间的差异显著(P<0.05)
3.4 叶片-凋落物-土壤有机C、全N、全P、全K含量和化学计量比的相关性
采用Pearson相关分析法对喀斯特高寒干旱区经济树种叶片、凋落物以及根区土壤之间的有机C、全N、全P、全K含量及其化学计量比进行两两比较。由表3可以得出,根区土壤有机C、全N与叶片全P、C∶K呈显著正相关(P<0.05,下同),根区土壤有机C与叶片有机C呈极显著负相关(P<0.01,下同),根区土壤有机C与叶片全K、C∶P呈显著负相关;根区土壤全N与叶片全K、C∶P、N∶P呈显著负相关;根区土壤全P与叶片全P、C∶K呈极显著正相关,与叶片全K、C∶P、N∶P呈极显著负相关,与叶片有机C呈显著负相关;根区土壤全K与叶片全K、C∶P、N∶P呈极显著正相关,与叶片全P、C∶K呈极显著负相关,与叶片有机C呈显著正相关;根区土壤C∶P与叶片全K、C∶P、N∶P呈极显著正相关,与叶片全P、C∶K呈极显著负相关,与叶片有机C呈显著正相关;根区土壤C∶K与叶片有机C呈极显著负相关,与叶片全P、C∶K呈显著正相关,与叶片全K、C∶P、N∶P呈显著负相关;根区土壤N∶P与叶片全K、C∶P呈极显著正相关,与叶片全P、C∶K呈极显著负相关,与叶片有机C、N∶P呈显著正相关。
如表4所示,根区土壤全N与凋落物全P、全K、C∶N呈极显著正相关,与凋落物全N、C∶P、C∶K、N∶P呈极显著负相关,与凋落物有机C呈显著负相关;根区土壤全P与凋落物全P、全K、C∶N呈显著正相关,与凋落物C∶P、N∶P呈极显著负相关,与凋落物全N、C∶K呈显著负相关;根区土壤全K与凋落物C∶P、C∶K、N∶P呈显著正相关,与凋落物全P、全K呈显著负相关;根区土壤C∶N与凋落物有机C、全N呈显著正相关;根区土
表3喀斯特高寒干旱区经济树种叶片与根区土壤有机C、全N、全P、全K含量和化学计量比之间的相关性
Table3RelationshipsbetweenOC,TN,TP,TKcontentsandstoichiometricratioinleafandrootzonesoilofeconomictreespeciesinkarstfrigidandaridarea
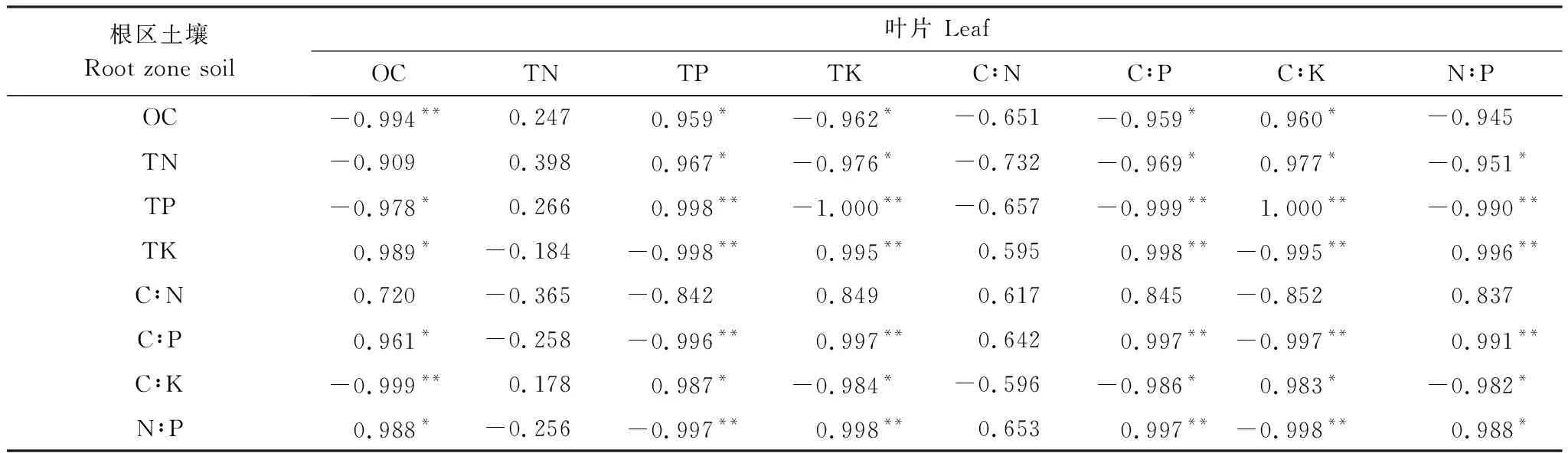
根区土壤Root zone soil叶片 LeafOCTNTPTKC∶NC∶PC∶KN∶POC-0.994**0.2470.959*-0.962*-0.651-0.959*0.960*-0.945TN-0.9090.3980.967*-0.976*-0.732-0.969*0.977*-0.951*TP-0.978*0.2660.998**-1.000**-0.657-0.999**1.000**-0.990**TK0.989*-0.184-0.998**0.995**0.5950.998**-0.995**0.996**C∶N0.720-0.365-0.8420.8490.6170.845-0.8520.837C∶P0.961*-0.258-0.996**0.997**0.6420.997**-0.997**0.991**C∶K-0.999**0.1780.987*-0.984*-0.596-0.986*0.983*-0.982*N∶P0.988*-0.256-0.997**0.998**0.6530.997**-0.998**0.988*
**极显著相关(P<0.01),*显著相关(P<0.05)
表4喀斯特高寒干旱区经济树种凋落物与根区土壤有机C、全N、全P、全K含量和化学计量比之间的相关性
Table4RelationshipsbetweenOC,TN,TP,TKcontentsandstoichiometricratioinlitterandrootzonesoilofeconomictreespeciesinkarstfrigidandaridarea

根区土壤Root zone soil凋落物 LitterOCTNTPTKC∶NC∶PC∶KN∶POC-0.745-0.8570.9310.9270.875-0.934-0.920-0.922TN-0.951*-0.993**0.995**0.996**0.999**-0.994**-0.997**-0.995**TP-0.898-0.964*0.990*0.988*0.971*-0.994**-0.989*-0.991**TK0.8770.944-0.973*-0.969*-0.9480.979*0.972*0.976*C∶N0.986*0.958*-0.895-0.899-0.9480.8940.9090.908C∶P0.9300.981*-0.992**-0.990**-0.984*0.996**0.994**0.997**C∶K-0.820-0.9060.954*0.9490.915-0.960*-0.949-0.954*N∶P0.8730.948-0.983*-0.980*-0.956*0.987*0.980*0.983*
**极显著相关(P<0.01),*显著相关(P<0.05)
壤C∶P与凋落物C∶P、C∶K、N∶P呈极显著正相关,与凋落物全P、全K呈极显著负相关,与凋落物全N呈显著正相关,与凋落物C∶N呈显著负相关;根区土壤C∶K与凋落物全P呈显著正相关,与凋落物C∶P、N∶P呈显著负相关;根区土壤N∶P与凋落物C∶P、C∶K、N∶P呈显著正相关,与凋落物全P、全K、C∶N呈显著负相关。
由表5可知,凋落物全N与叶片全K、C∶P、N∶P呈显著正相关,与叶片全P、C∶K呈显著负相关;凋落物全P与叶片C∶K呈极显著正相关,与叶片全K呈极显著负相关,与叶片全P呈显著正相关,与叶片C∶P、N∶P呈显著负相关;凋落物全K与叶片全P、C∶K呈显著正相关,与叶片全K、C∶P、N∶P呈显著负相关;凋落物C∶N与叶片全P、C∶K呈显著正相关,与叶片全K、C∶P、N∶P呈显著负相关;凋落物C∶P与叶片全K呈极显著正相关,与叶片C∶K呈极显著负相关,与叶片有机C、C∶P、N∶P呈显著正相关,与叶片全P呈显著负相关;凋落物C∶K与叶片C∶K呈极显著负相关,与叶片全K、C∶P、N∶P呈显著正相关,与叶片全P呈显著负相关;凋落物N∶P与叶片全K呈极显著正相关,与叶片C∶K呈极显著负相关,与叶片C∶P、N∶P呈显著正相关,与叶片全P呈显著负相关。
表5喀斯特高寒干旱区经济树种叶片与凋落物有机C、全N、全P、全K含量和化学计量比之间的相关性
Table5RelationshipsbetweenOC,TN,TP,TKcontentsandstoichiometricratioinleafandlitterofeconomictreespeciesinkarstfrigidandaridarea
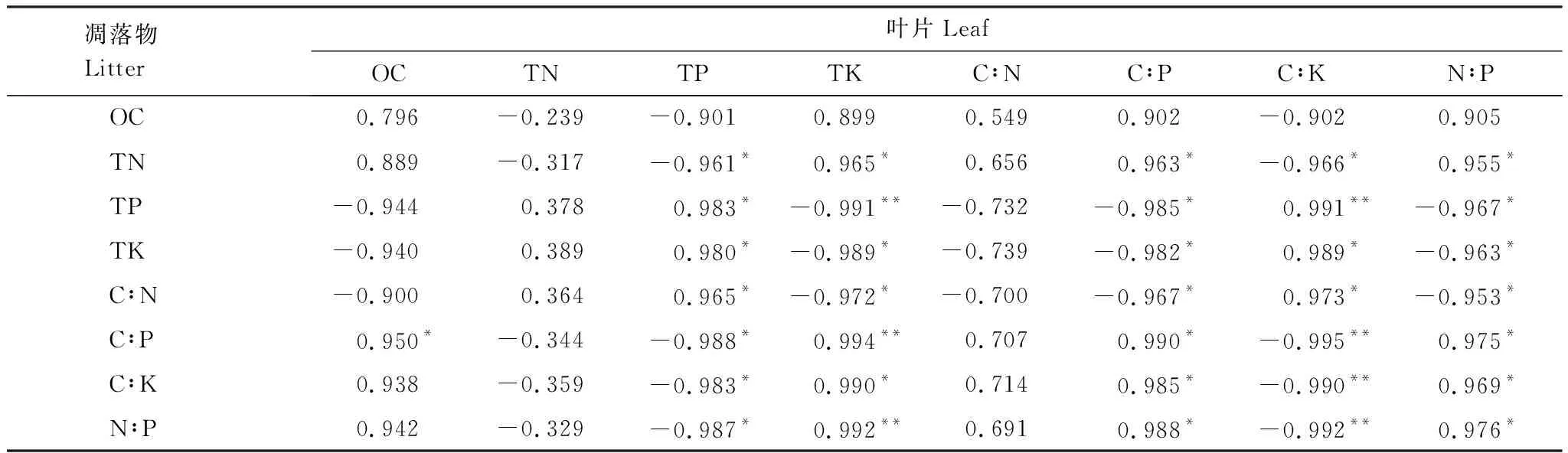
凋落物Litter叶片LeafOCTNTPTKC∶NC∶PC∶KN∶POC0.796 -0.239 -0.901 0.899 0.549 0.902 -0.902 0.905 TN0.889 -0.317 -0.961*0.965*0.656 0.963*-0.966*0.955*TP-0.944 0.378 0.983*-0.991**-0.732 -0.985*0.991**-0.967*TK-0.940 0.389 0.980*-0.989*-0.739 -0.982*0.989*-0.963*C∶N-0.900 0.364 0.965*-0.972*-0.700 -0.967*0.973*-0.953*C∶P0.950*-0.344 -0.988*0.994**0.707 0.990*-0.995**0.975*C∶K0.938 -0.359 -0.983*0.990*0.714 0.985*-0.990**0.969*N∶P0.942 -0.329 -0.987*0.992**0.691 0.988*-0.992**0.976*
**极显著相关(P<0.01),*显著相关(P<0.05)
4 结论与讨论
4.1 叶片-凋落物-土壤C、N、P、K含量特征
本研究结果表明,喀斯特高寒干旱区2种经济树种的有机C、全N含量以及核桃的全P含量均表现为叶片>凋落物>根区土壤。叶片C含量高于凋落物的C含量,这是由于凋落物在分解过程中初期分解粗有机化学成分中粗脂肪、单宁以及可溶性糖,使得凋落物中C含量显著降低[19]。还可能与叶片凋落前,植物生长速率变慢以及光合作用减弱有关。N、P都是可移动元素,植物叶片在凋落前,会有一部分养分转移到其他组分中,被重吸收利用,这导致叶片N、P含量通常高于凋落物N、P含量,这与白雪娟等[20]得出的3 种人工林不同组分中 N、P 含量大小均为叶片>凋落叶>土壤的研究结果一致。而研究区不同经济树种P含量在不同组分间的规律存在差异,这是叶片生长后期N、P由老叶向新叶迁移程度不同所引起的[20]。
本研究中,刺梨树种和核桃树种叶片有机C平均含量分别为429.28、436.73 mg/g,与曾昭霞等[21]研究的6种典型喀斯特林地植物的C含量均值(427.5 mg/g)相近,但均明显低于喀斯特峰丛洼地不同森林类型乔木叶片C含量平均值(496.15 mg/g)[14]和全球植物叶片C含量均值(464 mg/g)[22]。刺梨树种和核桃树种叶片全N含量均值(20.77、20.57 mg/g),略低于6种典型喀斯特林地植物叶片N平均含量(21.2 mg/g)[21],高于全国叶片N含量平均水平(20.2 mg/g)[23]和全球叶片N含量均值(20.1 mg/g)[24]。刺梨和核桃树种叶片全P含量(均值分别为2.10、1.52 mg/g)高于6种典型喀斯特林地植物P含量均值(1.2 mg/g)[21],也高于全国叶片P含量平均水平(1.46 mg/g)[23],其中刺梨树种叶片P含量(2.10 mg/g)还高于全球叶片P含量平均水平(1.77 mg/g)[24]。可见,与全国、全球叶片C、N、P含量相比,喀斯特高寒干旱区主要经济树种叶片呈现低C、高N、高P的规律。造成叶片C含量较低的原因可能是研究区内刺梨、核桃的栽种年限仅为3年,时间较短,还未到达生长旺盛期,养分循环以及养分利用效率较低,导致2类树种的叶片C含量偏低。同时,喀斯特高寒干旱地区特殊的生态环境、土壤特性、群落结构,以及受到不同的人为干扰程度均可导致本研究区的叶片C含量低于其他喀斯特地区。
研究区内,刺梨、核桃2种经济树种凋落物有机C含量均值分别为422.67、433.89 mg/g,全N平均含量分别为16.76、19.74 mg/g,全P含量平均水平分别为2.05、1.41 mg/g,与全球衰老叶片C、N、P均值(分别为467、10.0、0.7 mg/g)相比[25],C含量较低而N、P含量较高,与本研究中经济树种叶片呈现出的低C、高N、高P的规律一致,这表明凋落物秉承了植物叶片的性质。而本研究结果中C、N、P均值均高于桂西北喀斯特原生林凋落物C(402.47 mg/g)、N(12.93 mg/g)、P(1.03 mg/g)[21]平均含量以及次生林凋落物C(389.83 mg/g)、N(12.53 mg/g)、P(0.77 mg/g)均值[21],说明本研究区的生态环境优于桂西北喀斯特区域,植物凋落物可截留更多的养分。
本研究中,刺梨树种、核桃树种根区土壤有机C平均含量分别为31.81、27.52 mg/g,全N含量均值分别是3.84、2.81 mg/g,全P含量平均水平分别为2.06、0.74 mg/g,均高于俞月凤等[14]对喀斯特峰丛洼地人工林土壤研究得出的C、N平均含量(16.67,1.71 mg/g),核桃树种根区土壤P含量则低于喀斯特峰丛洼地人工林土壤P含量均值(0.94 mg/g)[14],同时两者均低于全球平均水平(2.8 mg/g)[26],与中国土壤P含量普遍较低的研究结果一致[26]。土壤P的来源比较固定,主要受成土母质的影响[27],这与喀斯特区域水土流失、岩石风化以及退化的石漠化生境有较大关系。
刺梨、核桃全K含量均呈现出根区土高于植物叶片、凋落物的规律,说明植物生长所需的K元素主要是从土壤中获取,因土壤中存在的K形态较多,而植物所能吸收利用的是速效K,故导致输入到植物叶片以及凋落物的K含量明显减少。在植物水分竞争中,K具有十分重要的作用,植物对K的吸收能力决定植物的耐旱性[28]。根区土壤速效K含量表现出核桃显著高于刺梨,表明核桃较刺梨具有更好的耐旱性,可能与核桃的喜光特性有关。
4.2 N、P再吸收率特征
植物生长所需N、P元素可从两方面获得,一方面植物可从土壤中直接吸收N、P元素,另一方面土壤中N、P供应缺乏时,植物会增强对该种元素的再吸收,加快养分循环速度、提高利用效率[12]。本研究中刺梨N再吸收率、P再吸收率分别为19.23%、2.39%,核桃N再吸收率、P再吸收率均值为4.05%、7.46%,刺梨N再吸收率显著高于核桃N再吸收率,说明刺梨与核桃相比,刺梨根区土壤N元素匮乏;而P再吸收率表现为核桃略高于刺梨,表明核桃根区土壤缺乏P元素可能性比刺梨大,与根区土壤全N、全P含量表现为刺梨高于核桃的研究结果不符,这是因为能被植物直接吸收利用的是有效养分[29],而非全量养分,故刺梨根区土壤碱解氮含量低于核桃根区土壤,而根区土壤速效P含量表现出刺梨显著高于核桃。与Han等[30]研究的全球植物N、P再吸收率平均水平(57.4%、60.7%)以及曾昭霞等研究的次生林N、P再吸收率平均值(36.5%、32.3%)[21]相比,刺梨与核桃N、P再吸收率均低于该研究结果,较高的再吸收率是贫瘠环境的适应机制,而本研究中较低的再吸收率说明本研究区N、P养分含量均较丰富,植物主要通过根区土壤吸收养分来适应环境,而非通过增强再吸收能力适应环境。
种群尺度与群落尺度的研究结果[17]存在差异,这可能是由于刺梨和核桃均处于幼龄林阶段,密度较小,根系分布范围有限,导致其主要依赖于对根区土壤养分的吸收,而远离根系的土壤养分未能得到充分利用。本研究结果表明,确定种植密度时,应该综合考虑对地上空间和地下空间的应用,特别是喀斯特地区,通过调控地下生境以提高地上部分生物量,对于植被恢复和可持续经营具有重要的理论和实践价值。
4.3 叶片-凋落物-土壤C、N、P计量比特征
叶片、凋落物和土壤C∶N、C∶P、N∶P代表着各组分为维持生态平衡以及为适应环境满足自身需求所面临的竞争[20]。其中,植物叶片的C∶N、C∶P能够反映其同化C的能力,在一定程度上反映植物营养的利用效率,具有重要的生态学意义[31]。本研究中刺梨、核桃叶片C∶N(20.68、21.23)、C∶P(204.30、286.66)与全球植物叶片的C∶N(22.5)、C∶P(232)[22]相比,C∶N均较低,刺梨叶片C∶P低于全球平均水平,可能是刺梨叶片的C含量过低导致这种差异,说明本研究区刺梨、核桃对C的利用效率均较低。N、P元素作为植物体内容易缺少的营养元素,是限制陆地生态系统生产力的重要因子[32]。有研究表明[33-34],植物生长的受N和P限制的N∶P临界值为14和16。N∶P小于14表示植物生长是受N限制的,N∶P大于16表示植物生长是受P限制的,N∶P在14—16时,植物生长同时受N和P的限制。而Güsewell[35]提出当N∶P小于10时,植物生长相对会受N限制,N∶P大于20表示植物生长相对受P限制。本研究中,刺梨、核桃叶片的N∶P分别为9.89、13.51,表明喀斯特高寒干旱区刺梨生长时主要受到N的限制,核桃生长时可能受到N、P的共同限制。刺梨、核桃叶片N∶P与全国平均水平(16.3)[23]、全球平均水平(13.8)[24]相比均较低。生长速率假说认为,生物体在高速生长过程中,通过富集大量P到rRNA中,以便核糖体能够快速合成大量蛋白质[36],故较低的C∶P、N∶P代表植物有较快的生长速率[22]。说明2种经济树种生长速率均较高,推测栽种年限相同时,刺梨幼树生长速率相对于核桃幼树较快,成树是否符合这个规律,需要进一步开展相关的生态化学计量研究才能确定。
N∶P是影响凋落物的分解和养分归还速率的重要因素之一,较低的N∶P使凋落物更易分解[12]。本研究中,刺梨凋落物N∶P为8.18,核桃凋落物N∶P为14.00,均低于Kang等[37]在全球尺度上研究的凋落物N∶P(18.32)以及桂西北喀斯特3种次生林凋落物N∶P(16.4)[21],说明刺梨、核桃凋落物的分解速率均较高。凋落物N∶P表现为核桃>刺梨,这使得刺梨的叶片凋落后,分解较快,不利于养分储存。相比之下,核桃凋落物能保留更多养分,归还到土壤的养分也随之增多,更多养分被输入到植物体内,最终植物才能更好地生长。喀斯特高寒干旱区因特殊的气候环境与地质环境,水土流失严重,土壤养分流失较多,故应选择保水保土功能较好的植物栽种,核桃的耐旱性较高,适应性较强,在喀斯特高寒干旱区栽种核桃有利于改善其生态环境。
本研究中,2种经济树种的根区土壤C∶N、C∶P、N∶P显著低于植物叶片及凋落物,这是由于凋落物在进入土壤转为土壤有机质的过程中还经历了复杂的微生物分解过程,在这个过程中大量有机态的C、N、P被矿化分解[38]。刺梨、核桃根区土C∶N为8.31、9.79均低于中国土壤C∶N范围(10—12)[39],同时低于全球土壤C∶N平均水平(13.33)[40],有机质分解速度与土壤C∶N是负相关关系[10],表明2种经济树种栽种区域有机质分解速率较快,不利于肥力的维持,但核桃根区土C∶N大于刺梨根区土C∶N,说明核桃地保肥能力较好一点。刺梨、核桃根区土C∶P(分别是15.45、37.10)显著低于全球森林0—10 cm土壤C∶P平均值(81.9)[41]、广西喀斯特地区0—10 cm土壤C∶P平均值(61)[21],较低的C∶P值体现本研究区域P有效性较高,产生差异的原因是由于本研究取土区域特殊,以及取土位置为根区附近土壤,根区土壤各种营养元素与表层土壤具有一定差异,除此之外,不同区域生长植物不相同,也会影响土壤中储存的养分含量,最终导致本研究区域的C∶P明显低于其他区域。
4.4 叶片-凋落物-土壤C、N、P及其比值的相关性
相关性分析可以揭示不同组分C、N、P化学计量比指标变量之间的协调关系,有助于对养分之间的耦合过程做出合理的解释[42]。有研究表明,土壤P含量与植物叶片P含量密切相关[43]。本研究中,根区土壤全P与叶片全P呈极显著正相关,相关系数达到0.998,这说明植物叶片中P含量的来源主要是土壤中P含量,植物根系通过吸收土壤中P元素供给叶片,满足叶片生长所需P元素。根区土壤全P与叶片全P呈极显著正相关,根区土壤C∶P与叶片C∶P、N∶P呈极显著正相关,根区土壤C∶P与叶片有机C呈显著正相关,表明植物生长过程中叶片养分含量的丰富程度与土壤养分含量密不可分。根区土壤全N与凋落物全P、C∶N呈极显著正相关,可见,根区土壤中N含量主要来自于凋落物。根区土壤与凋落物中N、P有较好的相关性,这是因为凋落物中有一部分N、P会归还到土壤里,成为土壤养分库的主要来源。“叶片-凋落物-土壤”是一个相互联系的复杂有机整体,植物叶片通过光合作用固定C,产生有机物,将其转移或以凋落物形式补给到土壤中,凋落物分解后养分返还土壤,植物体可进行重吸收,所以整个系统的生态化学计量特征具有明显的差异性和关联性[44],这一点在本研究中也有体现。相关性结果表明,喀斯特高寒干旱区经济树种叶片、凋落物以及根区土壤C、N、P及其比值之间有紧密的联系。可见,生态系统内部C、N、P元素循环是在叶片、凋落物以及土壤3个库之间转移和运输的[45]。
本研究仅对喀斯特高寒干旱区2种经济树种进行了养分含量以及生态化学计量方面的研究,所选的研究区域有限,如果想要全面评价喀斯特高寒干旱区植物生长过程的限制性养分以及更进一步理解“叶片-凋落物-土壤”之间的联系,下一步需要在喀斯特高寒干旱区开展更大尺度、更多经济树种类型的生态化学计量学研究。
猜你喜欢
中国农业大学学报(2022年11期)2022-11-07
农业现代化研究(2022年3期)2022-06-29
大众科学(2021年10期)2021-12-23
大众科学(2021年10期)2021-12-23
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
现代园艺(2017年21期)2018-01-03
中国生态农业学报(中英文)(2017年2期)2017-02-16
文化月刊·下旬刊(2014年6期)2014-08-28
植物营养与肥料学报(2014年4期)2014-04-08
