
定向进化后Cne PRP8蛋白质内含子的剪接机理研究
2018-10-15 07:54李淑梦李曼孟清
生物学杂志 2018年5期
李淑梦, 李曼, 孟清
(东华大学 生物科学与技术研究所, 上海 201620)
蛋白质内含子(intein)是在前体蛋白质中的一段多肽序列,其能够借助自身发生的催化反应由前体蛋白质中成功地剪切出来,同时将两侧的蛋白质序列(蛋白质外显子,extein)以天然肽键连接,形成成熟蛋白质[1-2],这一过程被称为蛋白质内含子介导的蛋白质剪接。蛋白质内含子根据其结构特征可分为3类,即标准蛋白质内含子(canonical intein),微小蛋白质内含子(mini-intein)和断裂蛋白质内含子(split-intein)[3]。由标准蛋白质内含子和微小蛋白质内含子介导的蛋白质剪接称之为蛋白质顺式剪接(protein cis-splicing);由断裂蛋白质内含子介导的蛋白质剪接被称之为蛋白质反式剪接(protein trans-splicing)。蛋白质反式剪接的发现开辟了重组蛋白质合成的新途径,即可将一个蛋白质分两段表达后再剪接起来形成完整的蛋白质。蛋白质反式剪接还可用于基因治疗[4-7],转基因植物的防扩散[8-9],高通量的蛋白质相互作用的筛选、线粒体蛋白质的鉴定[10],以及蛋白质核转运成像等[11];体外蛋白质反式剪接可用于核磁共振研究中对大的蛋白质进行分段同位素标记[12-13],可用于蛋白质芯片的制备[14];断裂蛋白质内含子也可被用于蛋白质及多肽的环化[15-16],使环化后的蛋白质热稳定性和抗蛋白水解酶的能力增加,使环化后的多肽药物具有更好的性能。
到目前为止,从原核生物基因组中已经分离鉴定了超过600多个蛋白质内含子,且绝大多数内含子都位于DNA复制、修复以及转录相关的蛋白质中。按照现有的剪接机理假说,蛋白质剪接反应主要依靠1位(蛋白质内含子N端第一位氨基酸)和正1位(C端蛋白质外显子N端第一位)的氨基酸,认为这两个位点的氨基酸对于发生蛋白质剪接至关重要[17]。但是,当把这些蛋白质内含子克隆并且插入其他生物的蛋白质时,绝大多数蛋白质内含子失去了原有的剪接功能[18-19],即使按照现有的剪接机理假说,将蛋白质内含子插入靶蛋白质中,形成C端蛋白质外显子的第一位氨基酸为C/S/T时,也不发生剪接反应;少部分发生蛋白质剪接,但是剪接效率极低;只有部分蛋白质内含子插入模式蛋白质(如麦芽糖结合蛋白和硫氧还蛋白)中发生高效剪接反应,少数蛋白质内含子能够在外源蛋白质中发生剪接,剪接效率也很低[20]。
为了提高蛋白质内含子在生物体内和体外的剪接效率,使其真正在蛋白质工程研究和应用领域中获得应用,我们把能够在模式蛋白质的体内和体外剪接合成融合蛋白、但在其他蛋白质中不发生剪接的CnePRP8蛋白质内含子(真菌病原体CryptococcusneoformansAD血清型剪接体蛋白PRP8中的蛋白质内含子)定向进化。进化前和进化后1位和正1位的氨基酸并没有发生突变,进化前CnePRP8蛋白质内含子不剪接,进化后就发生高效率的蛋白质剪接,表象就是进化后的CnePRP8蛋白质内含子内部的氨基酸发生了7处突变,如图1。这种剪接现象与科学家们提出的剪接机制大不相同。

图1 进化后的Cne PRP8蛋白质内含子氨基酸突变示意图Fig 1 Schematic of amino acid mutations in the Cne PRP8 protein intein after evolution
注:中间红色区域为定向进化之后的CnePRP8蛋白质内含子,其定向进化后突变的氨基酸位点分别被标注,其两端是KanR的N端和C端片段
本研究以我们前期获得的剪接活性高、通用性好的进化后的CnePRP8蛋白质内含子(命名为CPE)作为研究对象,通过对蛋白质内含子关键部位的氨基酸进行突变、恢复突变,研究CPE中影响其剪接反应的关键氨基酸;通过对相邻剪接位点的蛋白质外显子氨基酸的突变, 研究外显子影响CPE剪接的关键氨基酸。本研究通过定向进化突变和恢复突变确定影响CnePRP8蛋白质内含子剪接反应的关键氨基酸,探索其剪接机理,为获得具有高剪接效率和通用性的蛋白质内含子提供一定的理论支持,拓宽蛋白质内含子的应用领域。
1 材料与方法
1.1 突变体克隆的构建
引物的设计。根据CPE 7个突变位点的DNA序列分别设计恢复突变引物(52-P1、52-P2、68-P1、68-P2、114-P1、114-P2、122-P1、122-P2、146-P1、146-P2、152-P1、152-P2、172-P1、172-P2、1位-P1、1位-P2、正1位-P1、正1位-P2)。9对引物都由上海生工生物工程技术服务有限公司测序合成,见表1。

表1 引物序列
本课题前期对CnePRP8蛋白质内含子进行定向进化的结果图如图2所示,克隆体的构建过程为:将CnePRP8蛋白质内含子插入改造后的PKH载体的KanR合适位点上,并通过定向进化的过程得到CPE,质粒载体PKH的KanR抗性基因的N端具有6His-Tag,同时在克隆阶段CnePRP8蛋白质内含子内部也引入了6His-Tag,最后通过Western Blot检测证实了其定向进化后剪接效率大大提高。通过对CPE测序得到定向进化之后蛋白质内含子内部关键位点的氨基酸突变。
以本课题前期获得的CPE为模板,使用高保真的Phusion酶(NEB公司),用以上合成的一对引物或者其组合对其进行PCR扩增,循环条件为98℃预变性30 s;98℃变性5 s,55℃退火15 s,72℃延伸4 min,循环30次;终止延伸2 min。对PCR产物进行回收之后用去磷酸化酶DpnI(Thermo公司)切掉磷酸化的模板DNA,反应条件为37℃,1 h。酶切后再次回收,送到睿迪生物有限公司测序,获得序列正确后即为恢复点突变后的克隆。
1.2 蛋白质的表达及剪接
将测序正确的质粒转入E.coliDH5α中,于含有氨苄青霉素(100 μg/mL)的 LB(1 L∶10 g蛋白胨,5 g酵母提取物,10 g NaCl)培养基中37℃培养过夜。
收集菌体后,加入1×SDS-PAGE样品缓冲液,沸水浴裂解细胞,收集样品进行聚丙烯酰胺凝胶电泳(SDS-PAGE),进行Western Blot印迹检测(anti-His antibody),质粒载体PKH的KanR抗性基因的N端具有6His-Tag,同时CPE内部也含有6His-Tag,以用于Western Blot检测,分析各个突变体的剪接反应,利用Image J软件分析Western Blot条带灰度计算其剪接效率(剪接效率为剪接产物与剪接产物和剩余前体总和的比值)。
2 结果
为了验证CPE中7个突变的氨基酸对蛋白质剪接的影响,我们对CPE中的7个突变后的位点进行了恢复突变,包含单个位点的恢复突变、两个位点组合的同时恢复突变、部分3个位点组合的同时恢复突变以及蛋白质内含子1位和正1位的突变,通过对其剪接效率来分析与CnePRP8蛋白质内含子剪接相关的氨基酸。
2.1 Cne PRP8蛋白质内含子经过6轮定向进化之后的结果
本课题前期通过对CnePRP8蛋白质内含子进行6轮定向进化的过程,得到了剪接效率高并且通用性较高的CPE。由图2的Western Blot结果图可以看出,前3个泳道分别是载体PKH、单独的CnePRP8蛋白质内含子PMCP以及定向进化之前的CnePRP8蛋白质内含子(此时的CnePRP8蛋白质内含子已经插入了PKH载体KanR的138位上)。而后面8个泳道分别是通过6轮定向进化得到的不同样品,用ImageJ软件对图片中的条带灰度进行分析,从而估算其剪接效率,得到了最高的剪接效率为78%,并将其命名为CPE。将CPE再次插入PKH载体KanR的不同位置上时,得到的剪接效率由表2所示,从而证明了CPE具有良好的通用性。在之后的实验过程中,选取了剪接效率较高但并未达到100%的插入169Ser位点的样品进行后续实验。
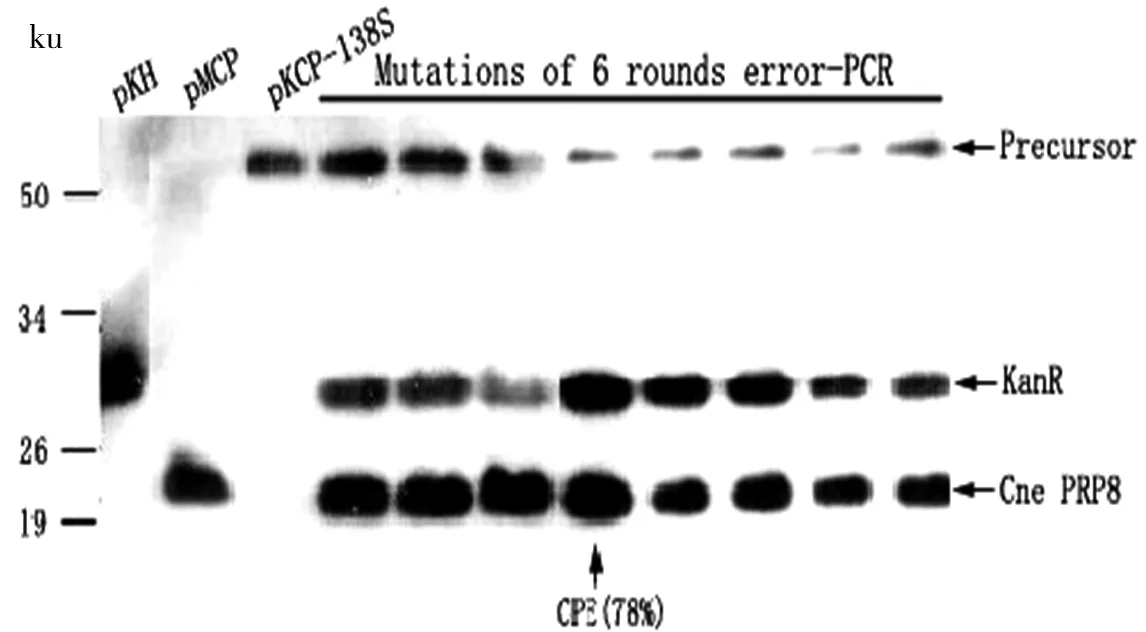
图2 Cne PRP8蛋白质内含子经过定向进化之后获得的突变体的剪接活性Fig 2 Splicing activity of mutants obtained after directed evolution of Cne PRP8 protein intein
2.2 CPE单个氨基酸位点恢复突变结果
由测序得知CPE的7个突变位点为:T52I、S68P、Q114R、G122C、T146I、N152D和S172R。对CPE的7个位点分别恢复突变。以CPE为模板,用表1中相对应的特异性引物和高保真的Phusion酶进行PCR实验并切除模板DNA后,转入大肠杆菌中表达,最后通过Western Blot印迹检测其剪接反应,通过条带灰度计算其剪接率。如图3所示,Precursor(~52.5 ku), KanR(~31.6 ku), CPE(~20.9 ku),其中间一条可能为断裂产物或His抗性的杂带。用ImageJ软件对图片中的条带灰度进行分析,从而估算其剪接效率,计算结果如表3所示。其中剪接率最低的是CPE-114和CPE-146,分别为58%和59%,说明114位和146位的氨基酸对蛋白质内含子的剪接反应具有较大影响。

1:CPE;2:CPE-52;3:CPE-68;4:CPE-114;5:CPE-122;6:CPE-146;7:CPE-152;8:CPE-172
图3 CPE的单个氨基酸位点恢复突变后的Western Blot
Fig 3 Western Blot result after one amino acid site recovery mutation of CPE

表3 CPE单个氨基酸位点恢复突变后的剪接效率
2.3 CPE两个组合氨基酸位点同时恢复突变结果
根据CPE 中7个突变位点,对其进行两两组合。以图3中单个氨基酸突变的克隆体为模板,用表1中各对引物和高保真的Phusion酶进行PCR实验并切除模板DNA后,转入大肠杆菌中表达,最后通过Western Blot免疫印迹检测其剪接反应,并估算其剪接率。如图4所示,Precursor(~52.5 ku), KanR(~31.6 ku), CPE(~20.9 ku),其中一条可能为断裂产物或His杂带,用ImageJ软件对图片进行分析,从而估算其剪接效率。计算结果如表4所示。结果可以看出,CPE-52-68和CPE-52-114的剪接效率明显很低,分别为24%和26%,说明这两个组合的氨基酸同时突变对蛋白质内含子的剪接反应影响较大。
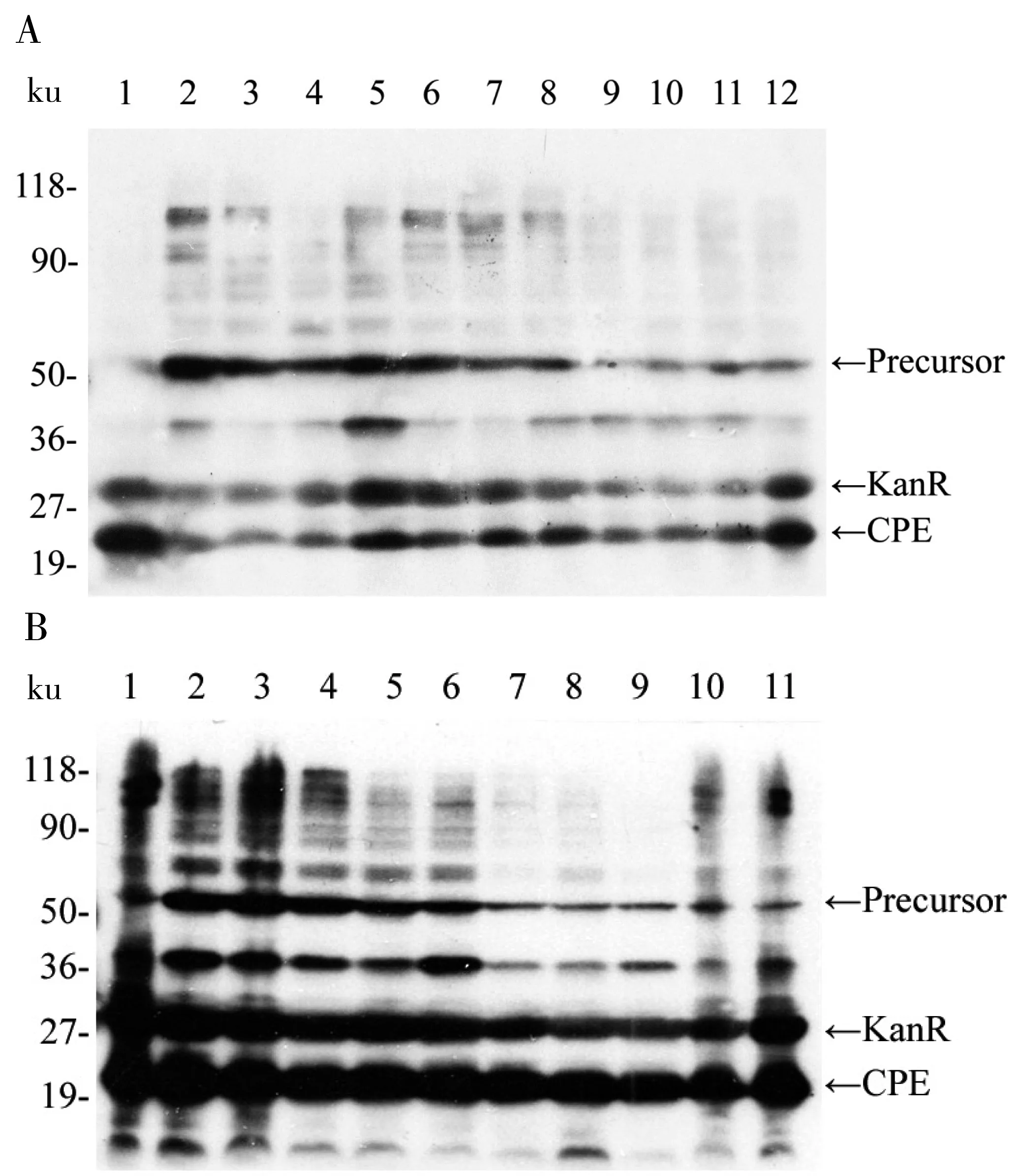
A:1为CPE,2为CPE-52-68,3为CPE-52-114,4为CPE-52-122,5为CPE-52-146,6为CPE-52-152,7为CPE-52-172,8为CPE-68-114,9为CPE-68-122,10为CPE-68-146,11为CPE-68-12,12为CPE-68-172。B:1为CPE,2为CPE-114-122,3为CPE-114-146,4为CPE-114-152,5为CPE-114-172,6为CPE-122-146,7为CPE-122-152,8为CPE-122-172,9为CPE-146-152,10为CPE-146-172,11为CPE-152-172
图4 CPE的两个氨基酸位点恢复突变后的Western Blot
Fig 4 Western Blot result after two amino acid sites recovery mutation of CPE

表4 CPE两个组合氨基酸位点恢复突变后的剪接效率
2.4 CPE部分3个氨基酸位点恢复突变结果
根据CPE 中7个突变位点,对其进行部分三三组合。选取了图3中剪接效率最低的CPE-114和CPE-146,以及图4中剪接效率最低的CPE-52-68和CPE-52-114为对象,将其组合成3个位点的同时恢复突变的蛋白质内含子进行实验。以图4中两个氨基酸突变的克隆体为模板,用表1中各对引物和高保真的Phusion酶进行PCR实验并切除模板DNA后,转入大肠杆菌中表达,最后利用Western Blot印迹检测,并估算其剪接率。如图5所示,Precursor(~52.5 ku), KanR(~31.6 ku), CPE(~20.9 ku),其中一条可能为断裂产物或His抗性的杂带,用ImageJ软件对图片进行分析,从而估算其剪接效率。计算结果如表5所示。从结果可以看出,3个样品的剪接效率相差不大,则可以根据之前的结果进行分析。
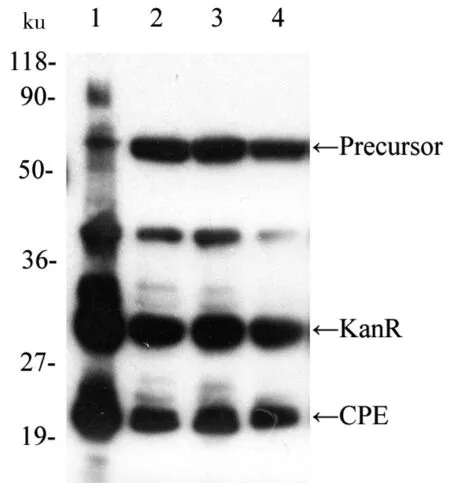
1:CPE;2:CPE-52-68-114;3:CPE-52-68-146;4:CPE-52-114-146
图5 CPE的部分3个氨基酸位点恢复突变后的Western Blot
Fig 5 Western Blot result after part three amino acid sites recovery mutation of CPE

表5 CPE部分3个组合氨基酸位点恢复突变后的剪接效率
2.5 CPE的1位和正1位突变结果

1:CPE;2:CPE-one site;3:CPE-positive one site
图6 CPE的1位和正1位的氨基酸位点突变后的Western Blot
Fig 6 Western Blot result after mutation of the one site and the positive one site amino acids of CPE
对CPE的1位和正1位分别突变看其对蛋白质剪接的影响。CPE的1位是半胱氨酸Cys,正1位是丝氨酸Ser,将其分别突变成丙氨酸Ala。以CPE为模板,用表1中各对引物和高保真的Phusion酶进行PCR实验并切除模板DNA后,转入大肠杆菌中表达,最后通过Western Blot印迹,检测其剪接反应。如图6所示,1位和正1位突变的样品图中显示3条带,Precursor、断裂产物以及杂带。图中没有CPE蛋白质内含子的条带,则可以看出蛋白质内含子并没有完全剪接,大部分发生了断裂反应,这是因为1位和正1位氨基酸残基的突变,造成蛋白质内含子无法进行剪接过程中所必需的亲核攻击反应,阻碍了蛋白质内含子完成整个剪接反应,因此,对于来自真核生物的蛋白质内含子CnePRP8,其1位和正1位氨基酸直接影响其剪接反应的进行。
3 讨论与结论
现研究表明,蛋白质内含子的剪接活性与其1位和正1位的氨基酸密切相关,以进行剪接过程所包括的两步转酯反应和两步亲核攻击反应,但现在关于蛋白质内含子剪接机理的研究大多局限于源自原核生物的蛋白质内含子,对于源自真核生物的蛋白质内含子的剪接机理少有研究。本课题以源自真核生物的蛋白质内含子CnePRP8为研究对象,探索其剪接机理,希望可以为蛋白质内含子的剪接机理提供新的观点和理论支持。我们通过对蛋白质内含子CnePRP8进行定向进化,得到具有高剪接效率,且具有一定通用性的新蛋白质内含子CnePRP8的突变体(CPE),通过对其全长测序,确定了该新突变体的氨基酸序列。为探索CnePRP8的剪接机理,我们对定向进行中突变的氨基酸残基,进行恢复突变,并突变其关键位置氨基酸,研究参与蛋白质内含子CnePRP8剪接过程的相关氨基酸,以其更广泛地应用于蛋白质研究领域。
本文首先通过对CPE中的定向进化造成的7个突变的氨基酸位点进行恢复突变,并通过Western Blot检测其剪接反应,评估其剪接效率。新突变体的剪接效率越低,则说明其相对应的突变的氨基酸对Cne PRP8的剪接反应影响越大。Western Blot结果表明,其中两个位点组合同时恢复突变的CPE-52-68(I52T,P68S)和CPE-52-114(I52T,R114Q)的剪接效率改变最大,同时单个位点突变时,114(R114Q)和146(I146T)的剪接效率都偏低,说明T52I、S68P、Q114R、I146T位点对CnePRP8的剪接反应影响较大;当对其3个位点的组合突变时,3个突变体的剪接效率相差不大,但仍高于CPE-52-68(I52T,P68S)和CPE-52-114(I52T,R114Q)突变体的剪接效率,说明3个突变的组合抑制了CnePRP8蛋白质内含子的剪接。我们对两个氨基酸恢复突变体的剪接效率进行分析,CPE-52-114(26%)和CPE-68-114(57%)相比可以看出I52T比S68P的影响大;CPE-52-68(24%)和CPE-52-114(26%)相比可以看出S68P比Q114R的影响大;CPE-52-114(26%)和CPE-52-146(50%)相比可以看出Q114R比I146T的影响大。由于对于真核生物蛋白质内含子的二级结构的研究仍然不成熟,所以蛋白质内含子剪接活性中心所含有的具体氨基酸并不确定。但是我们可以根据实验结果推测这几个位点氨基酸的突变对蛋白质内含子活性中心构建的影响。我们对这4个位点的氨基酸结构和性质进行分析:分析52位和146位的氨基酸性质得知,从CnePRP8到CPE,52位和146位都是苏氨酸Thr突变成异亮氨酸Ile,苏氨酸Thr为极性中性不带电荷氨基酸,侧链基团为羟基-CH(CH3)-OH,空间位阻较小,而异亮氨酸Ile为非极性中性不带电荷氨基酸,侧链基团为-CH(CH3)-CH2-CH3具有较大的空间位阻,推测可能会影响蛋白质内含子剪接活性中心的重建,由此可以推断不同极性或侧链基团的氨基酸的改变对蛋白质内含子的剪接具有较大影响;68位点的突变是从CnePRP8到CPE丝氨酸Ser突变成脯氨酸Pro,而丝氨酸Ser为极性中性不带电荷氨基酸,侧链基团为-CH2-OH,脯氨酸为非极性中性不带电荷氨基酸,侧链基团为-C3H6,Pro通常参与形成蛋白质二级结构中的β-turn结构,因此推测其可能影响蛋白质内含子的剪接活性中心的空间构象;而114位点的突变是从CnePRP8到CPE是谷氨酰胺Gln突变成精氨酸Arg,谷氨酰胺Gln是极性不带电的氨基酸,而精氨酸Arg是带正电的碱性氨基酸,氨基酸突变之后带了正电荷,从而空间结构发生了改变,影响了CnePRP8蛋白质内含子的剪接反应。而对于其他几个氨基酸位点的影响,其对于CnePRP8蛋白质的剪接是有促进作用的,但是对于CnePRP8蛋白质内含子的剪接活性中心的构建影响较小,当然CnePRP8具体的剪接活性中心的高级结构需要后续进一步的解析和研究。现有的蛋白质剪接机理表明,决定蛋白质内含子的剪接反应的关键氨基酸是其1位和正1位氨基酸,因此我们对CnePRP8的1位和正1位氨基酸进行突变时,结果表明CnePRP8不再发生剪接反应,而仅发生了蛋白质内含子的断裂反应,说明CnePRP8与其他原核来源的蛋白质内含子类似,其1位和正1位氨基酸直接影响其剪接反应的进行。
综上所述,本研究通过定性进化与恢复突变,探索了真核来源的蛋白质内含子CnePRP8的剪接机理,实验结果表明,蛋白质内含子CnePRP8的1位和正1位氨基酸对蛋白质内含子的剪接反应具有决定性影响,这与现有的蛋白质剪接机理是相统一的;同时,通过恢复突变研究结果表明,真核生物来源的蛋白质内含子CnePRP8,其剪接反应还与其他内部位点氨基酸密切相关,尤其是T52I、S68P、Q114R和I146T 4个位点的氨基酸的突变。氨基酸侧链基团和极性的改变直接影响了CnePRP8蛋白质内含子的剪接反应。本研究为CnePRP8蛋白质内含子提供了一定的理论支持,为获得高剪接效率和通用性的蛋白质内含子奠定基础,当然对于CnePRP8的剪接活性中心结构的研究和其空间高级结构的解析等都需要进一步的研究和探索。
猜你喜欢
建材发展导向(2021年14期)2021-08-23
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
中国煤层气(2019年2期)2019-08-27
生物工程学报(2019年6期)2019-07-10
生物学通报(2019年1期)2019-02-15
生物学通报(2018年12期)2018-10-10
上海农业学报(2017年3期)2017-04-10
环境与可持续发展(2017年2期)2017-04-06
安徽医科大学学报(2016年12期)2017-01-15
天津医科大学学报(2015年2期)2015-12-22
