
咖啡因延缓运动疲劳作用及机制研究进展
2018-10-16 02:57刘军乔德才刘晓莉
中国运动医学杂志 2018年9期
刘军 乔德才 刘晓莉
1西安体育学院健康科学系(西安710068)
2北京师范大学体育与运动学院(北京100875)
咖啡因是甲基黄嘌呤类化合物,分子式为C8H10N4O2,是世界上最为流行的中枢神经刺激剂,在自然界中,咖啡豆、茶和巧克力都是咖啡因的重要来源。咖啡因也被添加于软饮料和能量饮料中用于提神,发挥运动强力作用。2004年之前,大剂量使用咖啡因是被世界反兴奋剂组织(WADA)禁止的,后来经过重新评估咖啡因对人体的作用后,2004年版兴奋剂禁用清单中咖啡因被移除。有学者检测了此后4年(2004-2008年)的20686份运动员尿样,发现有73.8%的尿样中含有咖啡因,特别是在马拉松等耐力性项目和足球、橄榄球等团队项目中,咖啡因使用最为广泛[1]。
1 咖啡因作用有效剂量及可能机理
咖啡因生物利用度非常高,摄入后很快可被胃肠道吸收然后透过细胞膜进入组织,可通过血脑屏障,最快口服5 min后脑内浓度即可上升,60 min后即可达到血浆和脑内峰浓度。最终主要通过肝脏代谢,大约3%~10%在肾脏以原形排出,血浆中代谢半衰期大约为2.5~4.5 h[2]。
对于成年人来讲,每天400 mg左右的一般使用剂量远低于每天10 g的致毒剂量,对于12岁以下青少年,每天摄入咖啡因应不超过2.5 mg/kg。运动员使用咖啡因一般分为3种剂量等级,即高剂量(10~13 mg/kg)、中等剂量(5~6 mg/kg)和较低剂量(3 mg/kg)。但是高剂量使用咖啡因时会有一定的副作用,包括肠胃不适、紧张、精神错乱、注意力无法集中、失眠和一定程度的利尿等;中等剂量使用在发挥促力作用的同时副作用在减少,而较低剂量摄入时,促力作用依然存在,但使用者的心率、儿茶酚胺、乳酸、游离脂肪酸和甘油浓度都没有变化,推测较低剂量摄入咖啡因的促力作用可能是因为直接作用于中枢拮抗了腺苷受体[3]。动物实验也支持这一结论,Davis将咖啡因直接注射入大鼠大脑后观察对跑台运动至力竭大鼠运动能力的影响时发现,在中枢神经系统(CNS)以每只大鼠200 μg的剂量(0.6 mg/kg)使用咖啡因可以使跑台运动大鼠跑步至疲劳的时间增加50%,该剂量远小于外周有效剂量(6 mg/kg),而相同剂量用于外周是无效的,这同样表明低剂量咖啡因延缓疲劳的作用主要是依靠中枢机制,至少部分是因为拮抗了腺苷受体[4]。
体内腺苷浓度增加主要受三磷酸腺苷(ATP)代谢调控,运动时ATP分解增加导致腺苷浓度的增加。在中枢神经系统内,胞内腺苷主要由单磷酸腺苷(AMP)降解产生,纹状体胞外腺苷主要来自于胞内腺苷和环磷酸腺苷(cAMP)被磷酸二酯酶代谢的产物,所以腺苷可由激活细胞(高ATP消耗)和细胞积累cAMP(受体的刺激和腺苷酸环化酶正相关)所产生[5](见图1)。腺苷有4种受体,包括A1、A2A、A2B和A3,除A2A主要分布在纹状体,特别是局限分布于纹状体投射向苍白球外侧部(GPe)、表达有多巴胺(DA)D2受体(D2DA)、兴奋后释放脑啡肽(ENK)的中等棘状神经元(MSNs)上,其余3种在脑中有较广泛的分布。

图1 腺苷在纹状体γ-氨酸丁酸(GABA)能传出神经元中的产生[5]
2 咖啡因延缓运动疲劳作用与可能作用机理
咖啡因是腺苷A1和A2A的非特异性拮抗剂,诸多研究已经证明,咖啡因对人类和实验动物都有延缓疲劳的作用,在使用3~9 mg/kg咖啡因时,大强度跑步或自行车运动至疲劳的时间可以增加20%~50%[4]。另外,咖啡因对高强度运动,包括足球和橄榄球等需要持续很长时间的团队间歇运动项目都有益处[6,7],其机制可能涉及中枢和外周两个层面,但主要可能作用于中枢[8]。
2.1 咖啡因作用于腺苷受体延缓疲劳的中枢机制
运动冲动的发起和运动动机的产生都与DA关系密切。中枢内腺苷的产生可以抑制大脑兴奋性神经递质特别是DA的释放,降低DA、乙酰胆碱(Ach)合成,在中枢疲劳中扮演重要角色[9]。纹状体包括新纹状体和伏隔核都有密集的腺苷A2A受体和多巴胺D2受体,前者与运动功能关系密切,而后者与行为激活和努力相关的行为过程关系密切[10]。这提示咖啡因从运动冲动的发起和动机的产生两方面都将通过阻断腺苷受体而建立起一个多巴胺能的驱动[11]。最近的研究发现腺苷A2A受体和多巴胺D2受体可以形成二聚体从而影响后者的信号通路,当中枢腺苷产生增多时,会干扰DA对基底神经节到皮层间接通路的抑制作用,导致纹状体神经元兴奋性增强,基底神经节对皮层抑制过度使皮层发放的运动冲动不足诱发疲劳,而咖啡因则通过对间接通路去抑制后继续发放运动冲动[12,13],见图2。
多巴胺和环磷酸腺苷调节的磷蛋白-32(DARPP-32)是纹状体内包括腺苷、DA等多种神经递质信号转导过程中的整合因子,腺苷、DA等均可以通过与受体结合影响DARPP-32不同氨基酸残基的磷酸化而发挥不同的调节功能。有研究证实大鼠在力竭运动过程中,A2AR和D2DR是通过调节Thr34-DARPP-32磷酸化程度而发挥对运动疲劳的调控作用;运动至力竭时A2AR和D2DR调节作用异常,导致纹状体神经元兴奋性异常,使基底神经节对皮层的驱动不足引起疲劳[14,15]。当腺苷通过A2A受体刺激腺苷酸环化酶(AC)导致cAMP生成增多,进一步加强由腺苷通过cAMP/PKA级联激活的下游靶蛋白磷酸化;而咖啡因通过阻断A2A受体,减低cAMP产生,降低PKA的活性,这可依次减弱Thr34-DARPP-32的磷酸化,增加Thr75-DARPP-32的磷酸化,去放大cAMP/PKA通路的抑制作用[16](图3),最终诱导奖赏和行为敏化[17]。这从信号通路层面为咖啡因促力抗运动疲劳提供了证据。
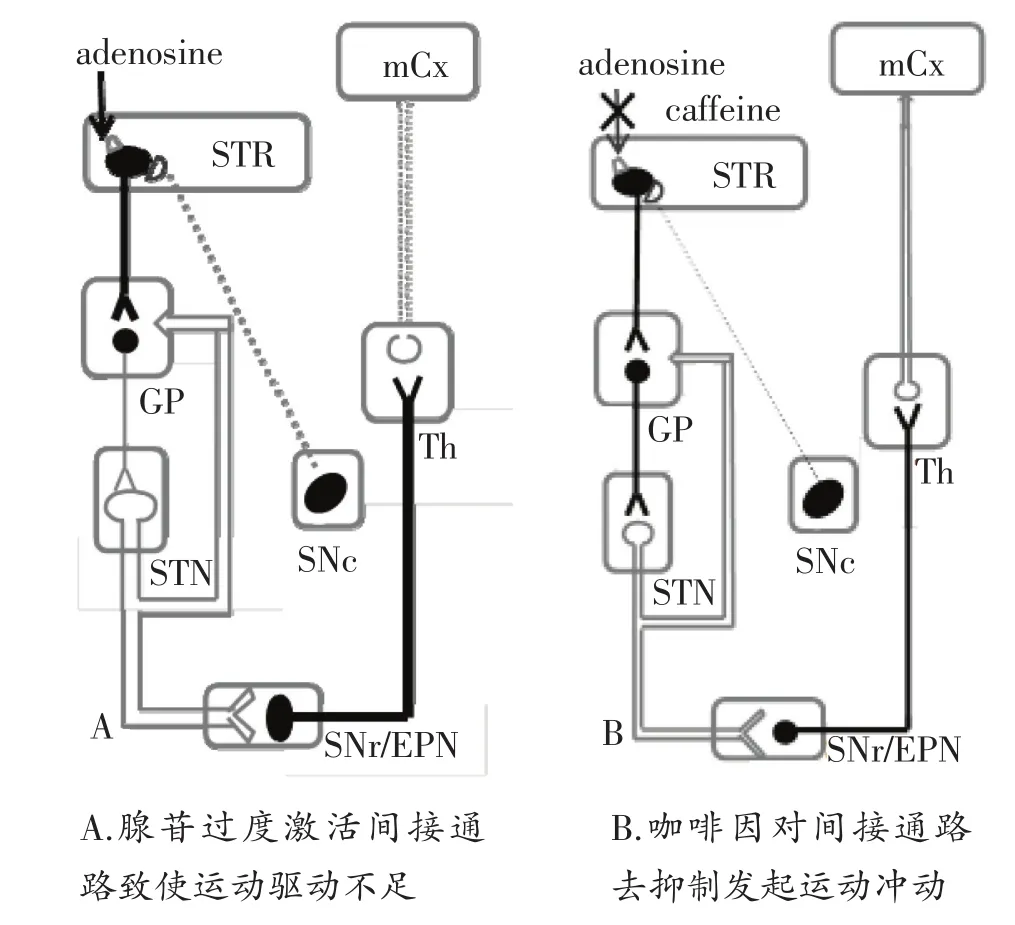
图2 咖啡因和腺苷作用于间接通路示意图
有研究设计了双盲、有安慰剂对照的实验去评价运动员分别摄入100 mg和200 mg咖啡因后对认知能力和心理能量的情绪衡量变化的敏感度,结果发现相比对照组,两组剂量的咖啡因组都削弱了人们对于能量消耗和疲劳感增加的敏感度,在认知能力和情绪状态活力指标测量等方面取得了一致的结果,认为低剂量(100 mg)咖啡因摄入即可以产生对刺激反应的提高、目标的识别,还可以改善情绪和表现[18,19]。其他研究也提示,咖啡因可以通过作用于控制运动功能相关的神经网络提高疲劳或者正常情况下的反应时[20],原因可能是通过作用于DA和肾上腺素,从而减轻疲劳对运动能力和中枢神经系统的影响[21,22](图4)。

图3 腺苷和咖啡因对PKA/DARPP-32调控示意图[16]

图4 咖啡因可通过很多途径影响肌肉效能[22]
2.2 咖啡因作用于腺苷受体延缓疲劳的外周机制
2.2.1 咖啡因可增加脂肪分解节约糖原以延缓疲劳
当运动开始后,肌肉工作消耗糖原使ATP消耗增加,细胞内腺苷生成增多,与脂肪细胞上的受体结合,降低脂肪酶的活性,抑制脂肪动员。此时,肌肉只能更多地依靠消耗肌糖原满足能量需求,肌糖原降低会使肌肉收缩效能下降。脂肪动员的限速酶对肾上腺素等激素十分敏感,咖啡因的摄入增加了肾上腺素和去甲肾上腺素的含量,因此使脂肪动员加快,导致运动开始后血浆中游离脂肪酸(FFA)增多,使机体对脂肪的利用增加,有效节省糖原,而使贮备有限的糖原服务于运动的后半程,从而延缓疲劳的发生。在1项研究中,受试自行车运动员需要完成2 h的等速自行车训练,但接受3种干预方式,分别为咖啡因、葡萄糖聚合物和安慰剂,共计500 mg咖啡因一半在运动开始前1 h摄入,其余在运动开始前15 min服用,结果发现,咖啡因组运动能力比对照组高了7.4%,比服糖组高了5.3%,运动2 h后咖啡因组脂肪氧化显著高于其余2组。而脂肪氧化是大强度耐力项目维持最后1 h运动的主要能源,这可能是咖啡因提高运动能力的另一机制,此外,服用咖啡因血浆FFA水平高于对照组30%[23]。另有研究证实优秀耐力运动员在急性摄入咖啡因后能耗和脂肪氧化增幅都大于相应递增的工作负荷[24]。这说明咖啡因除了主要通过中枢神经系统起作用之外,还可以影响运动中能量底物的利用率,在运动前或运动期间摄入咖啡因可以减少对糖原的依赖,增加对FFA的利用。另一研究也证实当功率自行车运动员摄入5 mg/kg咖啡因后,腿部肌肉脂肪氧化显著增加,当以80%VO2max蹬踏至力竭运动时,运动前摄入9 mg/kg咖啡因会使运动开始后肝糖原分解减少,说明咖啡因可以增加运动开始后肌肉内外对脂肪的依赖[2]。其他研究还显示咖啡因可以通过增加静息肌肉中乙酰辅酶A和柠檬酸盐的浓度而抑制糖原分解,通过抑制磷酸二脂酶提高运动时肌肉中cAMP的含量,从而促进新陈代谢,推测这也是咖啡因增力作用的机制之一[25]。
另外,机体基础代谢和交感神经活性与血浆去甲肾上腺素密切相关,增强交感神经活性可以刺激脂肪分解,增加基础代谢率[26]。研究发现,正常体重的健康男性,摄入50 mg的咖啡因,即使不运动,也可以在摄入后30 min内发现静息能耗显著增高,并至少持续4 h,使每日静息总能耗增加6%[27]。交感神经兴奋还可以促进呼吸道扩张、提高心脏输出能力,扩张血管加强组织新陈代谢,从而增强力量和增加反应速度。
2.2.2 咖啡因可通过调节Ca2+浓度增加肌肉收缩效能
腺苷A1受体存在于交感神经支配的靶器官中,腺苷通过和A1受体作用结合而拮抗β-AR激动剂产生抗肾上腺素能作用,也可以通过抑制Ach的释放而阻断突触后Ca2+内流。Ca2+是骨骼肌收缩的始动因素,咖啡因在肌肉收缩初期,可作用于兰尼碱受体(RyR)而影响Ca2+通道,促进肌浆网Ca2+的释放量,抑制肌浆网对Ca2+的重摄取而延长肌肉收缩活动的持续时间,使Ca2+更利于和肌钙蛋白结合而有利于横纹肌进行兴奋收缩耦联,咖啡因还可以通过调整神经肌肉接头的功能而增强骨骼肌收缩性能,原因可能是咖啡因可通过增加脑干区5-HT的浓度,通过向脊髓运动神经元发放兴奋性投射,增加骨骼肌运动单位的自我持续放电维持其兴奋性而延缓肌肉疲劳[28]。
2.2.3 咖啡因可通过降低对努力感知而延缓疲劳
中高强度运动时,伴随着肌肉持续收缩,粗细肌丝收缩蛋白出现微小损伤并引发炎症产生痛感,咖啡因(6 mg/kg)可能通过增加进行2 h 65%VO2peak后紧接高强度冲刺跑运动员血浆β-内啡肽浓度,而减轻大强度运动引发的肌肉不适感和疼痛感,特别是对于铁人三项等对耐力和强度都有很高要求的项目更为有效[29]。有学者通过测试60%VO2peak的30 min自行车试验期间,发现分别服用5和10 mg/kg咖啡因可以降低男女被试的肌肉疼痛强度和主观用力感觉(RPE)评分,证实摄入咖啡因(4 mg/kg)可以使中等缺氧情况下高强度自行车运动员力竭时间提高12%,推测原因可能是咖啡因降低了对努力的感知,虽然并没有真正降低肌肉疲劳[30]。
3 咖啡因与神经退行性疾病
长期中低剂量咖啡因摄入不只对运动耐力有益,对中枢神经系统和长时记忆也有积极效果[31]。流行病学研究显示,长期摄入咖啡因可能对认知损害和痴呆有防护作用,与帕金森疾病(parkingson’s disease,PD)和阿尔兹海默病(alzheimer disease,AD)等神经退行性疾病发病呈负相关[32]。
3.1 咖啡因与PD
PD病理改变为中脑黑质多巴胺能神经元变性死亡造成纹状体DA含量下降,从而导致进行性的运动功能损伤。其治疗一般是采用直接围绕提高相关脑区DA含量的策略,但经过长期治疗后疗效逐渐减退并可能引起“开关”症状、木僵症或出现幻觉等。所以非多巴胺能的治疗策略如咖啡因等选择性腺苷受体拮抗剂的研究备受关注。如前所述,腺苷A2A受体和多巴胺D2受体共表达于纹状体苍白球神经元,两种受体对运动行为施加相反的影响。用多巴胺或多巴胺D2受体激动剂刺激增强运动行为,而激活腺苷A2A受体则抑制多巴胺D2受体的信号[33]。咖啡因可能通过直接阻断腺苷受体或者间接作用于相关受体[34],激活多巴胺能通路[35],减低PD模型动物多巴胺能神经元的神经毒性和神经元变性[36]。
3.2 咖啡因与AD
AD发病可能与不同脑区腺苷受体的表达失衡密切相关,正常情况下,腺苷可对神经发生双向调节作用,具体则取决于结合的受体亚型和被激活的程度。AD发病过程中,A1受体逐渐降低,A2A受体表达逐渐升高,进而引起A1受体介导的抑制作用减弱而A2A介导的兴奋作用增强,最终引起AD患者认知功能障碍[37]。咖啡因并非特异性的A2A受体拮抗剂,对于AD患者却表现出特异性A2A拮抗剂相似的药理作用,推测可能是因为A1受体表达减少而A2A增加,被动使咖啡因提高了对A2A的选择性[38]。而事实上,AD发病是因为A1受体和A2A受体作用失衡的结果,咖啡因的非选择性拮抗可能纠正了这种紊乱而产生了治疗作用。
4 低剂量使用咖啡因更安全
尽管咖啡因由于可促进运动能力而被广泛使用,但也有置疑的研究提出,咖啡因特别是在高温情况下使用可能会造成尿液增多而破坏体液平衡,因此,有建议军事人员和高温中作业人员应该避免饮用含有咖啡因的饮料[39],特别是摄入咖啡因大于300 mg时会使尿容量急剧增加[40]。但最近的一项关于运动中使用咖啡因与尿液生成关系的meta分析文章中,研究者在分析咖啡因注射和尿液增加的效应量的16项研究后认为,咖啡因所产生的轻微利尿作用可能会被运动所抵消,因此,担心使用咖啡因尤其是运动前注射咖啡因造成体液流失的理由不充分,认为咖啡因对于运动员、健身爱好者、还是工人或者军人都是一种安全的强力剂,可以放心使用而不必担心因此造成的体液流失[41]。基于这些研究,在医学食品和膳食营养委员会专为军事营养研究所为军人制定的2001版报告中推荐150 mg的咖啡因摄入可增强身体的耐力和体能。
但大剂量使用咖啡因仍然需要谨慎,当使用剂量超过10 mg/kg或每天使用剂量超过600 mg时,运动能力并没有呈现出相应的剂量效应,但出现副作用的机率却大为增加。大剂量咖啡因可能通过阻断腺苷受体、通过增加β1-受体而诱发心动过速和心率失常,原因可能是增加胞内环磷酸鸟苷(cGMP)进而影响到心肌收缩相关的Ca2+释放,还有可能是因为刺激了Na+-K+-ATP导致血浆K+被转运至胞内使循环浓度降低导致膜电位下降[42]。除此之外,高剂量咖啡因摄入还可能诱发低血压,尽管这些结果仍然存在争议,但对于初次大剂量摄入咖啡因的运动员来讲依然非常值得警惕。在中枢神经系统,高剂量咖啡因摄入会诱发焦虑,但正是因为咖啡因具有内在的镇痛效果,在长期摄入后停止可能会导致戒断综合症出现[43]。需要注意的是,尽管WADA已经将咖啡因使用移出禁用清单目录,但全美大学体育协会(NCAA)依然规定咖啡因使用尿中浓度不得超过15µg/mL[43]。此外,咖啡因使用者往往无视安全剂量警示,在大剂量使用咖啡因的同时,还忽略诸如咖啡、能量饮料等不同来源提供的咖啡因的累积作用,如果再同时摄入酒精和尼古丁,使用风险就更为加大。
5 小结与展望
咖啡因促力抗疲劳作用的主要机制可能是通过阻断腺苷受体而发挥的,在中枢,咖啡因通过腺苷调节多巴胺能信号通路,从激活运动神经元驱动运动和保持运动动机方面增加运动能力;在外周,咖啡因通过拮抗腺苷受体,增加交感神经活性,降低肌肉疼痛感,增强脂肪酸氧化、节约糖原等延缓运动疲劳。中低剂量使用咖啡因是有利于运动能力且较安全的。
猜你喜欢
检察风云(2022年5期)2022-04-05
中国体育科技(2022年1期)2022-03-10
中国康复(2021年6期)2021-11-30
青岛大学学报(医学版)(2021年5期)2021-11-17
中医眼耳鼻喉杂志(2021年2期)2021-07-21
科技创新与应用(2021年16期)2021-01-04
杭州化工(2020年2期)2020-08-31
首都体育学院学报(2019年5期)2019-10-18
海外星云 (2019年14期)2019-08-12
中国医药指南(2017年3期)2017-11-13
