
高温、干旱及其复合胁迫对多歧苏铁生理影响研究*
2018-10-23 11:17郑艳玲马焕成
西南林业大学学报 2018年5期
郑艳玲 马焕成
(西南林业大学西南地区生物多样性保育国家林业局重点实验室,云南 昆明 650224)
现代苏铁类植物包括苏铁科 (Cycadaceae)、托叶铁科 (Stangeriaceae) 和泽米铁科 (Zamiaceae) 11属约290种,主要分布在热带和亚热带地区,除少数种类外,通常不能忍受冰冻和严寒。我国仅有苏铁属 (Cycas) 1属,且是该属植物的多样性分布中心,但由于人为采挖和栖息地破坏等因素,现存种群正面临灭绝的威胁。我国苏铁属所有种类均已列为国家一级保护植物,云南分布的苏铁属植物占全国的一半左右[1]。其中具三回羽状二叉分歧复叶的多歧苏铁 (Cycasmultipinnata) 属于叉叶类苏铁,在云南省分布于红河州的屏边、河口和个旧等地,是热带石山季雨林或山地雨林下的热带森林植物区系成员[2]。研究表明,植物的分布区未必是其生长最适宜的地区而是其竞争力最强的地区,同样植物的分布区域狭窄,可能仅仅是因为地理障碍限制了其迁移[3]。多歧苏铁的分布区域较为狭窄,其生态适应性如何及其是否与该物种的局限分布及濒危状况有关尚不清楚。
干旱是全球气候变化的主要结果和主要表现之一,近半个世纪以来全球范围内干旱发生的频率和强度都在急剧增加[4]。近年来,在全球气候变化的大背景下,我国的气候与环境也发生了显著变化,气候变暖导致区域性干旱事件连年发生。近50年西南地区总体呈气温升高,降水减少,干旱化加重的趋势。对于干旱的多数研究特别是室内研究仅考虑干旱对植物的影响,然而干旱可能会发生在任何季节,因此,温度与干旱紧密联系,但是它们的复合胁迫是如何影响植物的却研究甚少。本研究以多歧苏铁离体叶片为材料,研究高温、干旱及其复合胁迫对多歧苏铁生理指标的影响,这对了解多歧苏铁的濒危机制及气候变化背景下多歧苏铁的生态适应性提供理论依据,进而为多歧苏铁的引种驯化提供参考。
1 材料与方法
1.1 实验材料及其处理
在云南省河口县 (东经103°24′,北纬22°36′) 从多年生多歧苏铁植株上剪下叶柄顶端部分,插在水中,当天带回昆明。第2天进行高温、干旱及其复合胁迫处理。共有4个处理,以在30 ℃水中处理2 h的叶片作为对照 (CK)。高温处理时,将叶柄插在盛水的广口瓶中,在50 ℃培养箱中暗处理2 h;干旱处理时,将叶柄插在0.40 g/mL的聚乙二醇 (PEG) 中于30 ℃处理24 h。干旱复合胁迫时,将在0.40 g/mL的PEG中于30 ℃处理24 h的叶片再在50 ℃培养箱中处理2 h。每处理设4次重复,每次重复1个叶片。
1.2 实验方法
1.2.1色素含量测定
采用95%乙醇浸提法[5]测色素含量,4次重复测定。
1.2.2叶绿素荧光参数测定
测量前,叶片暗适应30 min。采用 PAM-2500调制式叶绿素荧光仪 (WALZ, Germany) 测定 ,光系统II (PSII) 最大光化学效率 (Fv/Fm)、PSII实际光化学效率 (YII)、光化学淬灭系数 (qP)、非光化学淬灭系数 (NPQ)、PS II处调节性能量耗散的量子产量 (YNPQ)、PS II处非调节性能量耗散的量子产量 (YNO) 和电子传递速率 (ETR) 均由仪器读取,4 次重复测定。
1.2.3丙二醛含量测定
采用硫代巴比妥酸法[5]测定丙二醛 (MDA) 含量,4次重复测定。
1.2.4抗氧化酶活性测定
超氧化物歧化酶 (SOD) 活性测定采用氮蓝四唑法;过氧化物酶 (POD) 活性测定采用愈创木酚法;过氧化氢酶 (CAT) 活性测定,以1分钟内240 nm下吸光度降低0.10为1个酶活性单位[5],4次重复测定。
1.2.5可溶性糖和脯氨酸含量测定
可溶性糖含量测定采用蒽酮比色法,脯氨酸含量测定采用茚三酮法[5],4次重复测定。
1.3 数据分析
各指标采用SPSS 15.0进行统计分析,用LSD进行差异显著性检验,用Excel 2007制图表,所有数据采用均值 ± 标准差表示。
2 结果与分析
2.1 高温、干旱及其复合胁迫对多歧苏铁叶片色素含量的影响
由表1可知,与CK相比,高温及干热复合胁迫处理后多歧苏铁的叶绿素a、叶绿素b、叶绿素 (a + b) 的含量均升高,干旱处理后以上指标均下降,但均与CK无显著差异。高温、干旱及干热复合胁迫处理后,叶绿素a/b均比CK升高,但与CK无显著差异。高温处理后,类胡萝卜素含量升高;但干旱和干热复合胁迫处理后其含量降低,但均与CK无显著差异。
2.2 高温、干旱及其复合胁迫对多歧苏铁叶片叶绿素荧光参数的影响
由表2可知,高温、干旱及干热复合胁迫处理后,Fv/Fm、Y(II) 和ETR均比CK显著下降 (P< 0.05),且高温及干热复合胁迫处理间Fv/Fm、Y(II) 和ETR无显著差异,但都显著低于干旱处理 (P< 0.05)。高温处理后的YNPQ比CK显著升高 (P< 0.05),但干旱和干热复合胁迫处理后的YNPQ与CK无显著差异;干旱处理后的YNO与CK无显著变化,但高温及干热复合胁迫处理后显著升高 (P< 0.05),且干热复合胁迫处理后YNO显著高于高温处理 (P< 0.05)。高温及干热复合胁迫处理后qP显著低于CK (P< 0.05),但干旱处理后qP与CK无显著变化;高温及干热复合胁迫处理后的NPQ与CK无显著差异,但干旱处理后比CK显著提高 (P< 0.05)。

表1 高温、干旱及其复合胁迫对多歧苏铁叶片色素含量的影响Table 1 Effects of heat, drought and their combined stress on pigment content of C.multipinnata leaves
注:不同小写字母表示差异显著。
注:不同小写字母表示差异显著。
2.3 高温、干旱及其复合胁迫对多歧苏铁叶片丙二醛含量的影响
由图1可知,高温及干热复合胁迫处理后,MDA含量均比CK显著降低 (P< 0.05),但干旱处理后MDA含量无显著变化。
2.4 高温、干旱及其复合胁迫对多歧苏铁叶片抗氧化酶活性的影响
由表3可知,高温处理后,SOD活性与CK无显著差异,但干旱及干热复合胁迫处理后SOD活性均比CK显著升高 (P< 0.05)。高温、干旱及其复合胁迫处理后,POD活性均比CK显著升高 (P<0.05),且干旱处理后POD活性最高。高温、干旱及其复合胁迫处理后,CAT活性均与CK无显著差异,但干热复合胁迫处理后的CAT显著高于高温及干旱处理 (P< 0.05)。
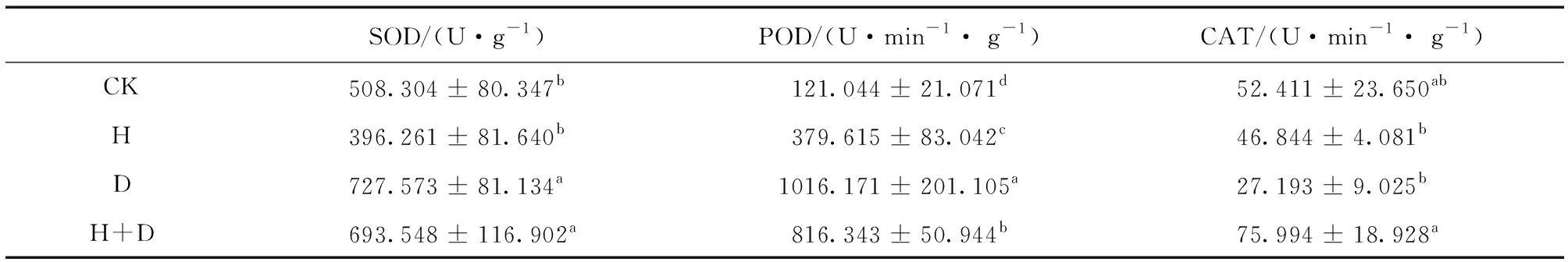
表3 高温、干旱及其复合胁迫对多歧苏铁叶片SOD、POD和CAT活性的影响Table 3 Effects of heat, drought and their combined stress on SOD, POD and CAT activities of C.multipinnata leaves
注:不同小写字母表示差异显著。

不同小写字母表示差异显著。
图1高温、干旱及其复合胁迫对多歧苏铁叶片MDA含量的影响
Fig.1 Effects of heat, drought and their combined stress on MDA content ofC.multipinnataleaves
2.5 高温、干旱及其复合胁迫对多歧苏铁叶片可溶性糖和脯氨酸含量的影响
由图2可知,高温处理后可溶性糖含量比CK升高9.51%,干旱及干热复合胁迫处理后,可溶性糖含量分别比CK降低4.75%和21.42%,但高温、干旱及干热复合胁迫处理后,可溶性糖含量与CK无显著差异。
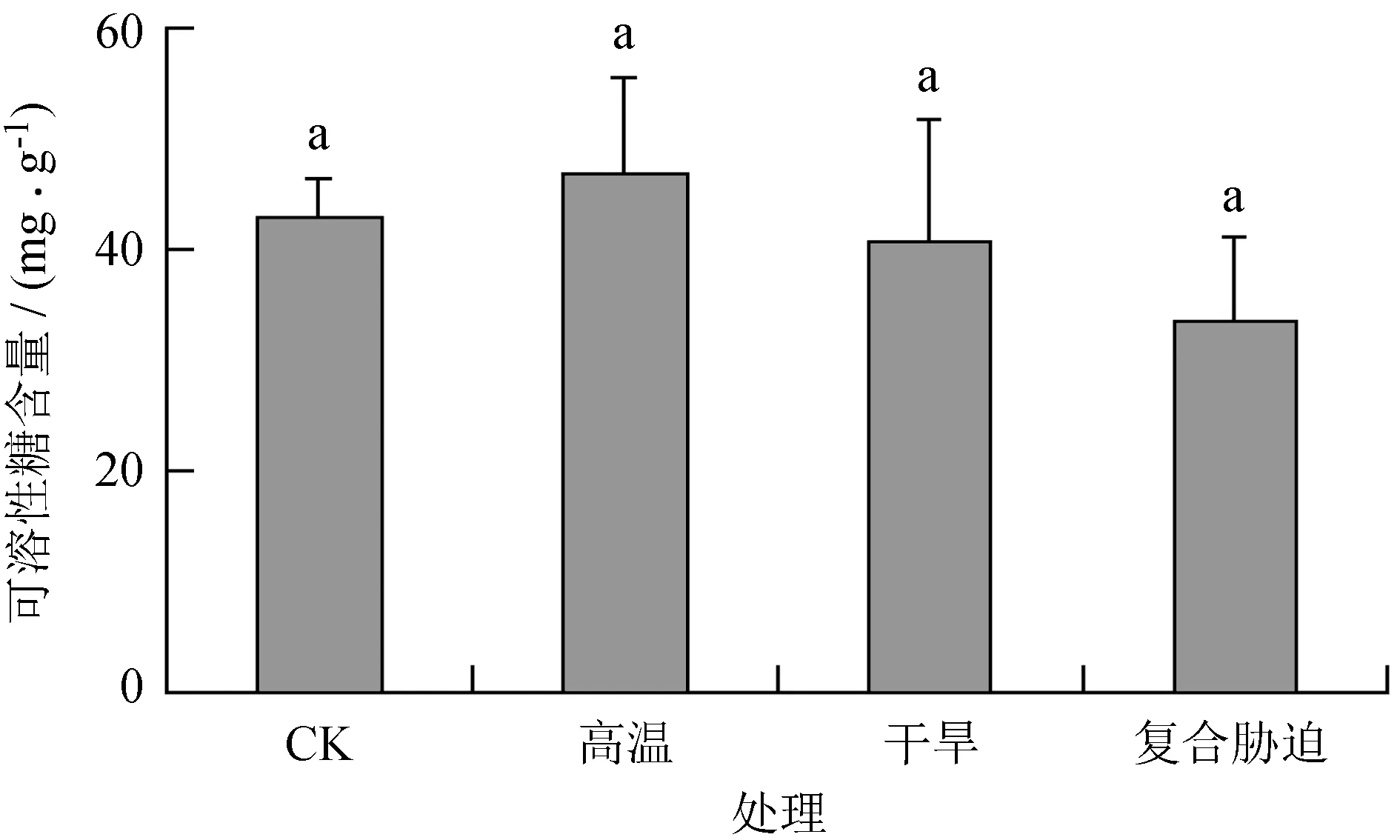
不同小写字母表示差异显著。
图2高温、干旱及其复合胁迫对多歧苏铁叶片可溶性糖含量的影响
Fig.2 Effects of heat, drought and their combined stress on soluble sugar content ofC.multipinnataleaves
由图3可知,高温、干旱及其复合胁迫处理后,可溶性糖含量分别比CK下降44.56%、61.07%和24.27%,脯氨酸含量均比CK显著下降 (P< 0.05),但干热复合胁迫处理后,脯氨酸含量与CK无显著差异。
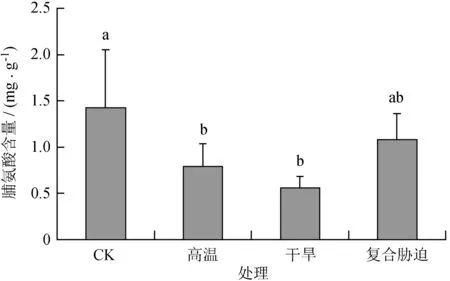
不同小写字母表示差异显著。
图3高温、干旱及其复合胁迫对多歧苏铁叶片脯氨酸含量的影响
Fig.3 Effects of heat, drought and their combined stress on proline content ofC.multipinnataleaves
3 结论与讨论
在复合胁迫下,被单一胁迫激活的信号通路可能被共同激活,同时复合胁迫特有的一些同路也可能被激活[6],因此植物可能需要独特的分子和代谢调控来适应不同胁迫的复合胁迫。在全球气候变化下,高温和干旱事件频发,且高温和干旱经常伴随发生,但高温干旱复合胁迫对多歧苏铁有何影响尚不清楚。30 ℃条件下,插在水中3天的多歧苏铁离体叶片形态上无显著变化,且其叶绿素荧光参数也无显著变化。在0.20 mg/g PEG中处理24 h的多歧苏铁离体叶片的Fv/Fm与CK并无显著差异[7],因此本研究将PEG浓度提高到0.40 mg/g。45 ℃及其以下温度热激不会影响多歧苏铁的光合活性[8],故本研究将温度设置为50 ℃。
叶绿素在光的吸收,能量传递和电子传递中起重要作用。在不同的胁迫条件下,植物的色素含量会降低或不发生变化[9-11],还有一些植物的色素含量会升高[12-13]。前期研究发现,多歧苏铁的叶绿素含量随温度升高呈现先降后升的变化趋势[8]。因此,高温、干旱及其复合胁迫处理后,多歧苏铁的色素组成和含量虽无显著变化,但并不表明这些胁迫对多歧苏铁的色素合成和降解没有影响。
叶绿素荧光参数可以指示植物的光合特性和胁迫耐性[14]。Fv/Fm,YII,ETR和qP分别反映PSII的潜在效率,线性电子传递的相对活性,通过PSII电子传递的相对速率和PSII反应中心的开放程度[15-16]。YNO是光损伤重要指标,数值升高表明植物不能有效控制所吸收的光能量[17-18]。高温和干热复合胁迫处理后,Fv/Fm、YII、ETR和qP的降低以及YNO的升高,表明多歧苏铁的光合活性在高温和干热复合胁迫下均下降。干热复合胁迫处理后YNO显著高于高温处理 (P< 0.05),表明干热复合胁迫对多歧苏铁的影响更大。干旱处理后Fv/Fm,YII和ETR比CK显著降低 (P< 0.05),但都显著高于高温和干热复合胁迫处理 (P< 0.05),表明多歧苏铁受到干旱胁迫的影响而导致光合活性下降,但干旱较高温及干热复合胁迫对多歧苏铁的影响要小。这与干旱处理后qP和YNO与CK无显著差异相一致。
生物膜会感知各种环境因子的胁迫而做出响应[19],MDA是膜脂过氧化产物,可以反映膜受损程度。对多歧苏铁而言,高温和干热复合胁迫对其光合活性影响较大,而干旱胁迫对其影响较小,但干旱处理后MDA含量无显著变化,高温和干热复合胁迫处理后MDA含量却显著降低 (P< 0.05)。对其它植物的一些研究也表明,MDA含量并不能指示一些胁迫的严重程度[20-21]。这种现象的机理尚不清楚,但它可能跟涉及膜脂过氧化过程的一些酶活性的变化有关。大量研究表明,抗氧化酶通过清除活性氧而在植物的抗胁迫过程中发挥重要作用[22],然而不同的酶在不同的胁迫条件下可能呈现出不同的响应模式,因此在植物抗胁迫过程中具体发挥作用的酶种类取决于物种和胁迫类型等因素[23-24]。在多歧苏铁受到高温、干旱及其复合胁迫时,SOD,POD和CAT活性也表现出不同的变化模式,高温下POD活性的升高以及干旱和干热复合胁迫下SOD和POD活性的升高,表明它们分别在多歧苏铁抗高温以及干旱和干热复合胁迫中起着重要作用。
渗透调节物质除了渗透调节功能外,还可作为渗透保护剂保护细胞的结构和功能[25]。 高温、干旱及其复合胁迫处理后,可溶性糖含量并无显著变化,且高温和干旱胁迫处理后脯氨酸含量显著降低 (P< 0.05)。由此表明,可溶性糖和脯氨酸并未参与到多歧苏铁对以上胁迫类型的适应过程中;其他研究也表明,特定渗透调节物质与植物抗胁迫的相关性及其在植物抗胁迫中的相对贡献因植物种类,品种和胁迫强度而异[25-28]。
综上所述,多歧苏铁受到高温、干旱及其复合胁迫处理后,光合活性下降。研究表明,多种胁迫的复合胁迫效应可能是相互拮抗或增效的[29]。根据叶绿素荧光参数判断,多歧苏铁受干旱影响最小,受干热复合胁迫影响最大,可见干旱不会减轻反而会加剧高温对多歧苏铁的生理影响,POD在抗高温胁迫中以及POD和SOD在抗干旱和高温复合胁迫中可能起了重要作用。在自然生境中,多歧苏铁的群落具多样性,反映其生态适应性较强[2]。前期研究也显示多歧苏铁对高温和干旱都具一定的耐受性[7-8],但是极端干旱和高温会降低多歧苏铁的代谢活性,尤其是当2种极端气候同时发生时这种影响效果会加剧。因此,生境破坏导致的生态因子的变化包括高温和干旱尤其是其复合胁迫是多歧苏铁濒危因素之一,在迁地保护时,也应考虑极端干旱和高温对该物种的影响。
猜你喜欢
中老年保健(2022年3期)2022-11-21
阅读(科学探秘)(2020年8期)2020-11-06
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
绿色科技(2019年2期)2019-05-21
天然产物研究与开发(2019年1期)2019-03-01
中国果业信息(2019年1期)2019-01-05
红蜻蜓·低年级(2017年2期)2017-03-29
中央民族大学学报(自然科学版)(2016年1期)2016-06-27
中央民族大学学报(自然科学版)(2016年2期)2016-06-27
