
饲粮中添加叶酸和维生素B12对五龙鹅生长性能、屠宰性能、脂肪沉积及极长链脂肪酸延长酶7基因表达量的影响
2018-11-16 06:07王宝维程漫漫张名爱葛文华
动物营养学报 2018年11期
王宝维 程漫漫 孔 敏 张名爱 岳 斌 葛文华
(青岛农业大学优质水禽研究所,国家水禽产业技术体系营养与饲料功能研究室,青岛 266109)
鹅脂肪合成的主要场所在肝脏,体内脂肪的合成与分解都是通过一系列的酶促反应来完成的,这个过程受到饲粮营养成分、内分泌激素、相关酶和相关基因等的调节[1],因此,从分子的层面上研究肝脏脂类代谢的机理有着重要意义。极长链脂肪酸延长酶7(very long chain fatty acid elongase 7,ELOVL7)是调控花生酸的强候选基因。花生酸参与机体脂肪酸代谢循环,是机体内重要的饱和脂肪酸。Yang等[2]利用282头苏太猪进行全基因组关联分析,鉴别影响花生酸的候选基因是位于16号染色体上的ELOVL7基因。研究表明,过量的摄入脂肪会促进ELOVL7基因过表达,导致前列腺癌细胞的增长,敲除ELOVL7基因后,前列腺癌细胞则会衰减[3]。因此,ELOVL7基因可能是阐明脂肪摄入量与前列腺癌关联的关键因素[4],维生素B12(VB12)作为一碳代谢的辅酶,可以提高叶酸利用率,促进多种DNA合成等反应。叶酸、VB12是对神经系统发育和维持生理正常功能起重要作用的维生素,是同型半胱氨酸(Hcy)代谢的必需辅助因子,大量研究显示,补充叶酸、VB12能降低体内Hcy水平和脑血管疾病发生的危险性[5]。Stekol等[6]研究了VB12和叶酸在大鼠体内胆碱合成中的作用,结果表明VB12缺乏减少了甘氨酸的利用,叶酸缺乏减少了丝氨酸的利用,较小程度地减少了甘氨酸的利用,这2种氨基酸能够形成乙醇胺,进而合成胆碱。已有研究表明,饲粮中不同水平的营养物质可使禽肉的脂肪酸组成和含量发生变化,而脂肪酸的合成受ELOVLs基因家族的调控。在家禽上对于ELOVL1~6基因的研究较常见,关于鸡的ELOVL7结构的研究已经取得了一些进展,而ELOVL7基因在鹅脂肪代谢过程中的功能和机理研究还处于空白。叶酸与VB12组合应用的研究多见于医学报道,而在家禽营养学上的研究基本处于空白。本试验开展了饲粮中添加不同水平叶酸和VB12对肝脏中ELOVL7基因表达量的影响及ELOVL7基因在五龙鹅不同组织器官中的表达差异性研究,丰富了营养物质调控鹅脂肪酸代谢以及极长链脂肪酸延长酶基因ELOVLs家族的研究内容,旨在从分子水平上探明饲粮中添加不同水平叶酸和VB12对五龙鹅肝脏中ELOVL7基因表达量、不同组织器官中表达差异性、脂肪沉积等的影响规律。
1 材料与方法
1.1 试验材料及饲粮
叶酸(96%)、VB12(1%)均由宁夏金维制药股份有限公司生产。按照试验设计要求,将叶酸和VB12按添加比例混入载体中,并逐级搅入全价料中,再放入搅拌机混合6 min。
基础饲粮以NRC标准(1994)[7]为主要参考依据,其组成及营养水平见表1。采用高效液相色谱法测得基础饲粮中叶酸和VB12含量分别为0.40和0.00 mg/kg。

表1 基础饲粮组成及营养水平(风干基础)
1)多维(不含叶酸和VB12)和微量元素为每千克饲粮提供 The multivitamin (without folic acid and VB12) and trace elements provided the following per kg of the diet: VA 1 500 mg,VD3200 IU,VE 12.5 mg,VK31.5 mg,VB12.2 mg,VB25.0 mg,烟酸 nicotinic acid 65 mg,VB62 mg,生物素 biotin 0.2 mg,泛酸 pantothenate 15 mg,胆碱 choline 1 000 mg,Fe 85 mg,Cu 5 mg,Mn 80 mg,Zn 80 mg,I 0.42 mg,Se 0.3 mg,Co 2.5 mg。2)叶酸和维生素B12为实测值,其他为计算值。Folic acid and VB12were measured values,while others were calculated values.
1.2 试验设计及饲养管理
试验用鹅由国家水禽产业技术体系育种基地高密银河润雁鹅业有限公司提供。选择初始平均体重差异不显著(P>0.05)的5周龄五龙鹅420只,随机分为7组,每组6个重复,每个重复10只鹅,公母各占1/2,Ⅰ~Ⅵ组为试验组,Ⅶ组为对照组。试验采用2×3两因素交叉等重复的析因设计,对照组饲喂基础饲粮(不添加叶酸和VB12),试验组在基础饲粮中分别添加0.25、2.00 mg/kg叶酸,0.003、0.009、0.018 mg/kg VB12,试验设计详见表2。试验期为4周。

表2 试验设计
试验前鹅舍要进行彻底全面的消毒,防止疾病的传播。采用网床平养,试验鹅自由饮水和采食,少喂勤添。搞好鹅舍内环境卫生,保持地面洁净干燥。观察鹅的健康状况,做好疾病防控工作。
1.3 样品采集与测定
1.3.1 生长性能测定
试验鹅第8周龄末,停饲12 h,逐只空腹称重,统计各组试验鹅的始重、末重和增重情况,计算平均日增重(ADG)和料重比(F/G)。
1.3.2 血清脂类代谢指标测定
试验鹅第8周龄末,各重复随机选择2只鹅,公母各占1/2,肉鹅停饲12 h后,颈静脉采血,于离心机3 000 r/min离心15 min,将分离的血清分装于1.5 mL的离心管置于-20 ℃冰箱中保存待测。
血清中葡萄糖(GLU)、总胆固醇(TC)、甘油三酯(TG)、低密度脂蛋白胆固醇(LDL-C)、高密度脂蛋白胆固醇(HDL-C)含量均由试剂盒进行测定,试剂盒购自南京建成生物工程研究所。
1.3.3 屠宰性能测定
试验鹅第8周龄末,各重复随机选择2只鹅,公母各占1/2,肉鹅停饲12 h后,颈静脉放血致死,测定屠宰性能指标包括屠体重、半净膛重、全净膛重、胸肌重、腿肌重,按照《家禽生产性能名词术语和度量统计方法》[8]测定,并计算屠宰率、半净膛率、全净膛率、胸肌率、腿肌率。
1.3.4 脂肪沉积测定
试验鹅第8周龄末,各重复随机选择2只鹅,公母各占1/2,肉鹅停饲12 h后,颈静脉放血致死,用湿法拔毛沥干水分后称重,测量腹脂重、皮脂重、皮脂厚、肌间脂带宽,按照《家禽生产性能名词术语和度量统计方法》[8]测定,计算腹脂率、皮脂率、胸肌肌内脂肪率、腿肌肌内脂肪率。
1.3.5 基因表达量测定
试验鹅第8周龄末,各重复随机选择2只鹅,公母各占1/2,颈静脉放血致死后剖开腹腔,无菌操作取出组织样品(心脏、肝脏、肾脏、腹脂、肺、肌胃、腺胃、胸肌、腿肌、脾脏、胰腺等),迅速收集到冻存管液氮保存,转移到-80 ℃冰箱保存待测。
选取肝脏样品50 mg,加入1 mL的TRNzol(Roche)试剂,粉碎匀浆后抽提RNA。使用Bio-Photometer型核酸蛋白测定仪测定其浓度和纯度,将检验合格的RNA用反转录试剂盒逆转录成cDNA用于基因表达量试验。实时荧光定量PCR反应体系为20 μL:SybrGreen qPCR Master Mix 10 μL,上、下游引物(10 μmol/L)各0.4 μL,DNA模板2.0 μL,ddH2O 7.2 μL。PCR反应程序为:94 ℃预变性5 min,94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸60 s,72 ℃修复延伸7 min,35个循环。同样对心脏、肾脏、腹脂、肺、肌胃、腺胃、胸肌、腿肌、脾脏、胰腺组织样品进行RNA逆转录为cDNA,用于基因组织表达差异性试验。
根据NCBI鹅ELOVL7基因登录号(NW_013185694.1),引物序列由Primer 5.0软件设计,以3-磷酸甘油醛脱氢酶(glyceraldehyde 3-phosphate dehydrogenase,GAPDH)为内参基因,引物由生工生物(上海)有限公司合成。基因引物序列见表3。
每个样品设置3个重复,采用2-△△CT法计算各样本中ELOVL7基因相对于内参基因GAPDH的表达量。

表3 基因引物序列
1.4 统计分析
利用SPSS 17.0软件中GLM模型分析主效应和互作效应,用ANOVA和LSD法对数据进行差异显著性分析。P<0.05为差异显著。
2 结果与分析
2.1 饲粮中添加不同水平叶酸和VB12对5~8周龄五龙鹅生长性能的影响
由表4可见,饲粮中添加不同水平的叶酸和VB12的交互作用对五龙鹅末重影响显著(P<0.05),试验Ⅴ组体重最大,对平均日增重、料重比影响不显著(P>0.05);Ⅴ组的末重显著高于Ⅰ、Ⅵ和Ⅶ组(P<0.05)。与对照组(Ⅶ组)相比,饲粮中添加不同水平的叶酸和VB12有提高平均日增重的趋势,但差异不显著(P>0.05)。由此表明,饲粮中添加不同水平的叶酸和VB12对5~8周龄五龙鹅生长性能产生影响,其中,添加叶酸0.25 mg/kg和VB120.009 mg/kg(Ⅴ组)对生长性能影响最显著。

表4 饲粮中添加不同水平叶酸和VB12对5~8周龄五龙鹅生长性能的影响
同行数据肩标无字母或相同字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)。ns=不显著。表5至表8同。In the same row, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while with different small letter superscripts mean significant difference (P<0.05). ns=not significant. The same as Table 5 to Table 8.
2.2 饲粮中添加不同水平叶酸和VB12对五龙鹅血清脂类代谢指标的影响
由表5可见,饲粮中添加不同水平叶酸和VB12的交互作用对血清TG、HDL-C和LDL-C含量影响显著(P<0.05)。饲粮中添加不同水平叶酸对血清TG和LDL-C含量影响显著(P<0.05);不同水平VB12对血清GLU和LDL-C含量影响显著(P<0.05)。与对照组相比,饲粮中添加不同水平叶酸和VB12有降低血清中脂类代谢指标的趋势。由此表明,饲粮中添加叶酸和VB12能够改善五龙鹅血清脂类代谢指标。

表5 饲粮中添加不同水平叶酸和VB12对五龙鹅血清脂类代谢指标的影响
2.3 饲粮中添加不同水平叶酸和VB12对五龙鹅屠宰性能的影响
由表6可见,饲粮中添加叶酸和VB12的交互作用对胸肌率影响显著(P<0.05),对屠宰率、半净膛率、全净膛率和腿肌率影响均不显著(P>0.05)。Ⅰ、Ⅴ组胸肌率显著高于对照组(P<0.05)。由此表明,饲粮中添加叶酸和VB12能够提高五龙鹅胸肌率,减少腹脂率,改变鹅胴体组织成分构成。

表6 饲粮中添加不同水平叶酸和VB12对五龙鹅屠宰性能的影响
2.4 饲粮中添加不同水平叶酸和VB12对五龙鹅脂肪沉积的影响
由表7可见,饲粮中添加不同水平叶酸和VB12的交互作用对皮脂率、腹脂率、肌间脂带宽、胸肌肌内脂肪率和腿肌肌内脂肪率影响显著(P<0.05)。其中,Ⅴ组腹脂率显著低于其他各组(P<0.05)。饲粮中添加不同叶酸和VB12对皮脂厚无显著影响(P>0.05)。由此表明,饲粮中添加叶酸和VB12能够降低五龙鹅脂肪沉积量。

表7 饲粮中添加不同水平叶酸和VB12对五龙鹅脂肪沉积的影响
2.5 饲粮中添加不同水平叶酸和VB12对五龙鹅肝脏中ELOVL7基因表达量的影响
由表8可见,饲粮中添加不同水平叶酸和VB12的交互作用对肝脏中ELOVL7基因的表达量影响显著(P<0.05);饲粮中添加不同水平叶酸和VB12均能使肝脏中ELOVL7基因的表达量高于对照组,Ⅴ组ELOVL7基因的表达量最高,显著高于对照组(P<0.05)。
2.6 肝脏中ELOVL7基因表达量与血清脂类代谢的相关性
由表9可见,肝脏中ELOVL7基因表达量与血清TG、LDL-C含量呈显著负相关(P<0.05),与TC含量呈极显著负相关(P<0.01);与血清GLU、HDL-C含量呈负相关,但不显著(P>0.05)。由此表明,肝脏中ELOVL7基因表达对血清脂类代谢具有调控作用,二者之间存在着同步反向调控机制。
2.7 肝脏中ELOVL7基因表达量与屠宰性能的相关性
由表10可见,肝脏中ELOVL7基因表达量与屠宰率呈显著正相关(P<0.05),与半净膛率、全净膛率、腿肌率呈负相关,但不显著(P>0.05),与胸肌率呈正相关,但不显著(P>0.05)。由此表明,肝脏中ELOVL7基因表达对五龙鹅的屠宰性能具有一定调控作用。
2.8 肝脏中ELOVL7基因表达量与脂肪沉积的相关性
由表11可见,肝脏中ELOVL7基因表达量与肌间脂带宽呈显著正相关(P<0.05),与皮脂率、腹脂率、腿肌肌内脂肪率呈显著负相关(P<0.05),与皮脂厚呈正相关(P>0.05)。由此可见,肝脏中ELOVL7基因表达量与鹅机体脂肪的沉积密切相关,饲粮中添加叶酸和VB12可通过调控肝脏中ELOVL7基因表达量来改善体内脂肪分配。

表8 饲粮中添加不同水平叶酸和VB12对五龙鹅肝脏中ELOVL7基因表达量的影响
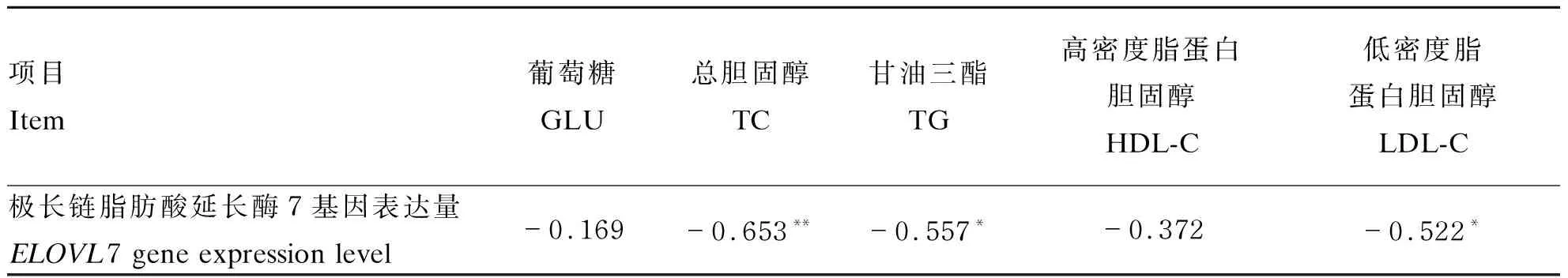
表9 肝脏中ELOVL7基因表达量与血清脂类代谢的相关性
*表示显著相关(P<0.05),**表示极显著相关(P<0.01)。下表同。* means significant correlation (P<0.05), and ** means extremely significant correlation (P<0.01). The same as below.

表10 肝脏中ELOVL7基因表达量与屠宰性能的相关性
2.9 ELOVL7基因组织表达差异
由图1可见,ELOVL7基因在鹅的心脏、肝脏、肾脏、腹脂、肺、肌胃、腺胃、胸肌、腿肌、脾脏、胰腺中均有表达,表达量依次为腹脂>肺>胰腺>肾脏>腺胃>心脏>肌胃>脾脏>肝脏>腿肌>胸肌。ELOVL7基因在腹脂中的表达量最高,在心脏、肝脏、肌胃、脾脏、胸肌、腿肌中的表达量差异不显著(P>0.05)。
3 讨 论
3.1 饲粮中添加不同水平叶酸和VB12对五龙鹅生长性能和屠宰性能的影响
叶酸和VB12是合成DNA、RNA的重要辅酶,对生命早期的生长发育非常重要,它们的缺乏是引起巨幼红细胞贫血的主要因素,孕期母体叶酸缺乏可导致胎儿发生神经管畸形[9]。不同叶酸水平的添加对肉仔鸡的生产性能有一定影响。随着叶酸添加水平的提高,肉仔鸡的饲料转化效率和日增重都在提高,表明叶酸作为快速生长的现代品系肉仔鸡体内合成嘌呤、嘧啶的必需物质和有效的甲基载体可促进机体的生长发育[10]。齐广海[11]报道,生长速度快的动物,对叶酸的需要量会提高。所以添加适宜水平的叶酸将能提高现代品系肉仔鸡的生长速度。本试验结果表明,饲粮中添加适宜水平叶酸和VB12对五龙鹅胸肌率和腹脂率影响显著,和对照组相比,胸肌率增加,腹脂率降低,这表明肝脏中ELOVL7基因表达对五龙鹅的屠宰性能具有一定调控作用。由此可以推断,饲粮中添加适宜水平叶酸和VB12能够改变鹅肉营养组成。

表11 肝脏中ELOVL7基因表达量与脂肪沉积的相关性

柱状数据标注相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。
3.2 饲粮中添加不同水平叶酸和VB12对五龙鹅肝脏中ELOVL7基因表达量的影响
Stabler等[12]研究表明,叶酸缺乏和VB12缺乏都可引起恶性贫血,叶酸补给虽然可以缓解因VB12缺乏造成的恶性贫血,但却引起神经系统的损伤。甲基丙乙醇和总的Hcy浓度是特异性诊断VB12缺乏的敏感指标,也是和叶酸缺乏有效区分开的指标。VB12在机体内主要通过2种物质吸收和转运,包括内源因子和钴胺转运蛋白。所以,VB12实际上与核酸和蛋白质的合成密切相关[13]。VB12依赖的是蛋氨酸合成酶,能催化一个甲基基团从甲基四氢叶酸上转移至Hcy,形成蛋氨酸,最终形成活性氨基酸(SAM)。缺乏VB12将减少DNA甲基化的SAM可利用量,从而影响其基因表达[14]。VB12影响着叶酸的代谢效率,并且参与嘌呤和核苷酸的合成,同时维持DNA的合成与修复,保证染色体的稳定性。本试验结果表明,饲粮中添加不同水平叶酸和VB12对五龙鹅肝脏中ELOVL7基因表达量有显著的影响,Ⅴ组ELOVL7基因的表达量最高,体重值也最大。本试验主要是为了研究叶酸和VB12联合对家禽肝脏ELOVL7基因表达量的影响,探明该基因对机体组织营养再分配的影响规律,为家禽的肉质研究提供理论依据;关于ELOVL7基因表达对脂肪酸代谢的机理还有待于继续深入研究。
3.3 五龙鹅肝脏中ELOVL7基因表达量与血清脂类代谢指标的相关性
脂肪酸是细胞的基本组成部分,在机体的能量存储、信息传递及代谢调控等方面起着重要的作用。脂肪酸是由一条长的线性碳氢链(疏水尾)和一个末端梭基(亲水头)组成的梭酸[15]。根据其碳氢链内是否含有双键又可分饱和脂肪酸和不饱和脂肪酸2类。高含量的单不饱和脂肪酸和多不饱和脂肪酸,如人体内必需脂肪酸ω-3系列,其有利于人类的心血管健康,然而高含量的饱和脂肪酸则会增加患心脏疾病的风险[16]。因此,脂肪酸是维持机体内稳态重要成分。研究表明,ELOVL7主要参与饱和脂肪酸的合成,ELOVL1、ELOVL3及ELOVL7共同完成C18∶0、C20∶0、C22∶0、C24∶0合成途径[17]。本试验结果表明,肝脏中ELOVL7基因表达量与血清TG、LDL-C含量呈显著负相关,与血清TC含量呈极显著负相关,与血清GLU、HDL-C含量呈负相关,但差异不显著。这表明肝脏中ELOVL7基因对血清脂类代谢具有抑制调控作用,可以推断,饲粮中添加叶酸和VB12可通过提高肝脏中ELOVL7基因表达量来改善机体脂肪代谢。
3.4 五龙鹅肝脏中ELOVL7基因的表达量与脂类沉积的相关性
花生酸参与机体脂肪酸代谢循环,是人体内重要的饱和脂肪酸。就目前所知,ELVOL7是调控花生酸的强候选基因[18]。目前,关于ELOVL7基因在脂类代谢中的研究多见于猪、小鼠和水产动物,而在鹅上的研究基本处于空白,其作用机制尚不明确。本试验肝脏中ELOVL7基因表达量与脂肪沉积的相关性分析结果表明,肝脏中ELOVL7基因表达量与肌间脂带宽呈显著正相关,与皮脂率、腹脂率、腿肌肌内脂肪率呈显著负相关。这可以推断饲粮中添加叶酸和VB12能直接影响肝脏中ELOVL7基因表达,进一步影响鹅机体脂肪分配,初步探明了遗传与营养因子(叶酸和VB12联合)的关系。
3.5 ELOVL7基因表达量在五龙鹅组织表达特异性
ELOVLs是极长链脂肪酸延长酶家族基因,最早来源于酵母ELO[19]。迄今,已在生物体内识别并鉴定出7种ELOVL蛋白ELOVLl~7,其中ELOVL1、ELOVL3、ELOVL6、ELOVL7主要参与饱和脂肪酸和单不饱和脂肪酸的合成,而ELOVL2、ELOVL4、ELOVL5主要参与多不饱和脂肪酸的合成[20]。有关研究表明,ELOVLs家族是极长链脂肪酸延长关键步骤的催化剂[18]。ELOVLs基因家族在动物体内具有重要的表达调控作用[21]。杨志刚等[22]对中华绒鳌ELOVLs基因表达分析显示,各组织中均有ELOVL基因表达,表达量最高的组织为肝胰腺和肠道,在心脏中表达量最低。敲除癌细胞中ELOVL7基因能有效减少细胞中C20、C22和C24脂肪酸的含量,并直接影响癌细胞生长[3]。本试验结果表明,ELOVL7基因在腹脂中的表达量最高,在腺胃、心脏、脾脏、胸肌、腿肌中的表达量较低且差异不显著,这表明ELOVL7基因在鹅不同组织中的表达量不同,该结果丰富了极长链家禽脂肪酸延长酶基因ELOVLs家族的研究内容,为进一步深入探究ELOVLs家族的表达特点奠定基础。
4 结 论
① 饲粮中添加不同水平叶酸和VB12对五龙鹅肝脏中ELOVL7基因表达量有干预作用,饲粮中添加叶酸0.25 mg/kg和VB120.009 mg/kg使肝脏中ELOVL7基因表达量最高。饲粮适宜的叶酸和VB12组合能够提高五龙鹅胸肌率,减少腹脂率,改变了鹅酮体组织成分构成。
② 肝脏中ELOVL7基因表达对五龙鹅血清脂类代谢具有调控作用,二者之间存在着同步反向调控机制。
③ 肝脏中ELOVL7基因表达量与屠宰率呈显著正相关,与半净膛率、全净膛率、腿肌率呈负相关,与胸肌率呈正相关,这表明肝脏中ELOVL7基因的表达量对五龙鹅屠宰性能产生影响。肝脏中ELOVL7基因表达量与肌间脂带宽呈显著正相关,与皮脂率、腹脂率、腿肌肌内脂肪率呈显著负相关。ELOVL7基因在腹脂中的表达量最高,其次是肺和胰腺;ELOVL7基因在胸肌、腿肌中的表达量较低。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
青年生活(2020年10期)2020-05-26
苏州教育学院学报(2020年2期)2020-02-25
中国生殖健康(2020年6期)2020-02-01
中国生殖健康(2019年12期)2019-01-07
中国生殖健康(2018年6期)2018-11-06
青年文学家(2017年24期)2017-08-22
动物营养学报(2017年8期)2017-08-16
妈妈宝宝(2017年4期)2017-02-25
地质与资源(2016年5期)2016-02-27
